Результаты исследований линейного роста лиманды (Limanda limanda L., 1758) Баренцева моря
 Баренцева моря")
Автор: Стесько Алексей Владимирович
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Статья в выпуске: 1 т.17, 2014 года.
Бесплатный доступ
Представлены результаты исследований роста лиманды Баренцева моря с помощью метода "biological intercept". Выполнена оценка варьирования изменчивости динамики индивидуальных показателей роста в течение первых пяти лет жизни рыбы. Проведено моделирование кривых роста.
Лиманда, обратные расчисления, рост - возраст, отолиты
Короткий адрес: https://sciup.org/14294666
IDR: 14294666
Текст научной статьи Результаты исследований линейного роста лиманды (Limanda limanda L., 1758) Баренцева моря
Лиманда (ершоватка, Limanda limanda L., 1758) – вид семейства камбаловых, широко распространенный в Северном, Белом и Баренцевом морях, а также возле берегов Исландии. Она обитает преимущественно в прибрежных районах на глубинах от 10 до 100 м, плотных промысловых скоплений не образует, встречается в приловах при промысле прочих видов донных рыб (Вилер, 1983).
Для изучения одной из основных характеристик любого вида рыб – темпа линейного роста – используется метод обратных расчислений. Он позволяет смоделировать темп роста рыб даже по сравнительно небольшой выборке. Ценность метода состоит и в том, что с его помощью можно узнать характеристики роста рыб младших групп, исследования которых затруднены, ввиду того что такие особи могут находиться в труднодоступных для проведения исследований местах (например, на мелководьях). Метод обратных расчислений известен с начала ХХ в. (Lea, 1910). Современными исследователями используются различные варианты расчета возраста рыб по чешуе или отолитам (Брюзгин, 1969; Campana, 1990).
На настоящее время наиболее полно исследован рост лиманды, обитающей у берегов Европы ( Lee , 1972; Osae, De Veen , 1972; Ortega Salas , 1988; Lozan , 1989; 1992; Rijnsdorp et al ., 1992; Bolle et al ., 2001; Lee et al ., 2006; 2007) и Исландии ( Josson , 1966; 2006; Dab Common , 2013). В работах А.С. Шерсткова (2005; 2007) отражены особенности роста лиманды Белого моря.
В меньшей степени изучены характеристики роста баренцевоморской лиманды. В литературе встречаются сведения о возрастном составе и сроках нереста лиманды, относящиеся преимущественно к первой половине ХХ в. ( Есипов, Сластников , 1932; Бараненкова , 1952; Андрияшев , 1954). Для решения задачи обновления информации ФГУП "ПИНРО" проведены исследования линейного роста лиманды Баренцева моря.
2. Материал и методика
Возраст рыб определяли по отолитам методом слома и обжига отолита ( Hassenger , 1991). Отолит разламывали в поперечном сечении так, чтобы слом проходил через ядро, обжигали в пламени спиртовки. Готовый препарат рассматривали в капле глицерина под стереомикроскопом под увеличением 32х в проходящем свете. Ввиду того что рост самцов и самок у большинства видов камбал отличается ( Lozan , 1989), данные обработаны и представлены отдельно по полам.
Для выяснения зависимости роста отолита от увеличения размеров тела лиманды измерялась длина отолита от переднего до заднего края (от парарострума до антирострума). Зоны роста отолита измерялись по большему радиусу при помощи шкалы окуляр-микрометра, а также видеокамеры DCM-310 и программы ScopePhoto (China). Данные были использованы в ходе проведения обратных расчислений роста. Для определения степени отличия результатов измерений правых и левых отолитов было проведено сравнение ширины случайно отобранных отолитов из проб лиманды ( N = 51 экз.). Коэффициент корреляции составил 0,91, что позволяет с одинаковой точностью провести расчисления по большему радиусу правых и левых отолитов. В целях обеспечения методической однообразности использовались преимущественно правые отолиты.
Рост рыб оценивали по наблюденным данным, с помощью модели Берталанфи ( Шибаев , 2007), а также методом обратных расчислений "biological intercept" ( Campana , 1990; Campana, Jones , 1992). Данный метод включает модифицированное уравнение Роза Ли ( Брюзгин , 1969) и подразумевает введение поправочных коэффициентов длины рыб и отолитов на ранних стадиях развития в основную формулу:
La = (Oa – Oc)(Lc – Li)(Oc – Oi)–1, где La – длина рыбы в возрасте а; Lc – длина выловленной рыбы; Oa – радиус годовых колец отолита рыбы в возрасте а; Oc – радиус годовых колец отолита выловленной рыбы; Li и Oi – размеры рыбы и отолита "biological intercept".
Оптимальными для определения параметров L i и O i являются значения, измеренные сразу после выклева личинки. Параметры для "biological intercept" могут быть взяты не обязательно при измерении особей из целевой популяции. Так, С.Е. Кампана и К.М. Джонс допускают использование личинок, искусственно выращенных при условиях, соответствующих среде обитания исследуемой популяции ( Campana, Jones , 1992). Подобных опубликованных данных по лиманде Баренцева моря не найдено. В.К. Есипов (1949) приводит лишь примерную длину личинок лиманды – 2-3 мм. Для расчетов были использованы параметры роста лиманды побережья Исландии, жизненный цикл которой проходит в условиях относительно сходных с условиями жизни баренцевоморской лиманды. Данные по размерам личинок лиманды после выклева получены Г. Джонсоном - 2,7 мм ( Jonsson , 1966). Аналогичное значение (2,7 мм) приводит О. Ли для лиманды Ирландского моря ( Lee et al ., 2006); он применяет метод "biological intercept" для обратных расчислений роста личинок лиманды, однако использует для этого лапиллус (lapillus), а не более крупный отолит – сагитту (sagitta). Для личинок такие результаты более надежны ( Morales-nin et al ., 1999), однако в настоящем исследовании результаты измерений О. Ли использовать невозможно, поскольку размеры и рост сагитты и лапиллуса у рыб отличаются ( Practical handbook… , 2009; Шелехов , 2000).
Размеры отолита высчитаны на основе аналогий, проведенных Г. Джонсоном по отношению к морской камбале как к близкому виду, а также аквариальных исследований морской камбалы Ф. Ховенкампа, Дж. Витте и Н.Г. Журавлевой ( Howenkamp, Witte , 1991; Журавлева , 1996). Расчеты проведены на основе прямой пропорции. Длина личинок морской камбалы при выклеве составила 7 мм, радиус отолита (sagitta) – 10 мкм. При известной ( Jonsson , 1966) длине личинок лиманды (2,7 мм) расчетный радиус отолита равен 3,9 мкм. Тогда поправочные коэффициенты для лиманды составили L i = 2,7 мм, O i = 3,9 мкм. Данные в единицах шкалы окуляр-микрометра переведены в единицы СИ. Измерения отолитов при работе с камерой DCM выполнены непосредственно в единицах СИ.
Статистическая обработка материала произведена по общепринятым методикам при помощи пакета программ MS Office ( Ивантер, Коросов , 2011).
В работе были использованы следующие условные обозначения: S – стандартное отклонение; CV – коэффициент вариации; M ± m – арифметическая средняя и ошибка средней; R – коэффициент корреляции; R 2 – коэффициент детерминации.
3. Результаты и обсуждение
Во всех обработанных возрастных пробах лиманды Баренцева моря доминировали самки в возрасте 8-11 лет и самцы – 3-5 лет (рис. 1).
На юго-востоке Баренцева моря (Западный и Восточный прибрежные районы, Канинская банка) наиболее часто встречались самки в возрасте 8-10 лет и самцы – 5-6 лет. На востоке Баренцева моря в Вайгачском районе преобладали рыбы обоих полов в возрасте трех лет. На западе Мурмана доминировали рыбы в возрасте 5-7 и 14-17 лет. Виду того что в рамках географического ( Limits… , 1953) и экосистемного ( Introduction… , 2012) районирования восточной границей Баренцева моря является пролив Карские ворота, в настоящей работе рассматривается рост лиманды Баренцева моря как единой популяции, независимо от района обитания.
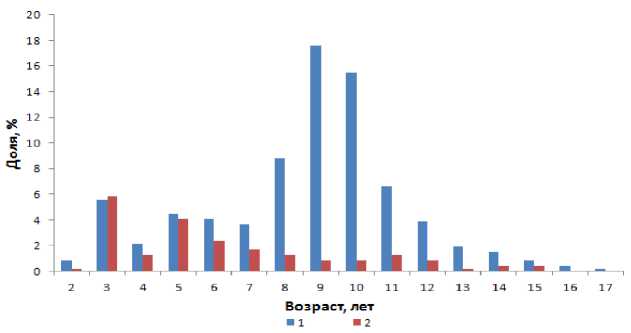
Рис. 1. Возрастной состав самок (1) и самцов (2) лиманды (N = 529 экз. и N = 144 экз. соответственно) в пробах за 1990-2013 гг.
Положительная корреляция между показателями ширины колец отолита (в мм) и длины рыбы (в мм) составила 0,90 для самок и 0,95 для самцов. Тренд линейной зависимости длины рыбы от ширины годовых колец отолита выявил отношение:
– для самок:
y = 192,32 x + 26,084;
– самцов:
y = 160 x + 16,085;
где х – ширина годовых колец; у – длина рыбы; при R 2 = 0,82 (самки) и R 2 = 0,90 (самцы).
Высокое значение коэффициентов корреляции и детерминации в отношении размеров отолитов и длины рыб позволяет применить методы обратного расчисления линейного роста.
Результаты проведенных обратных расчислений длины самок и самцов лиманды положительно коррелировали с наблюденным темпом роста ( R = 0,98) (рис. 2).
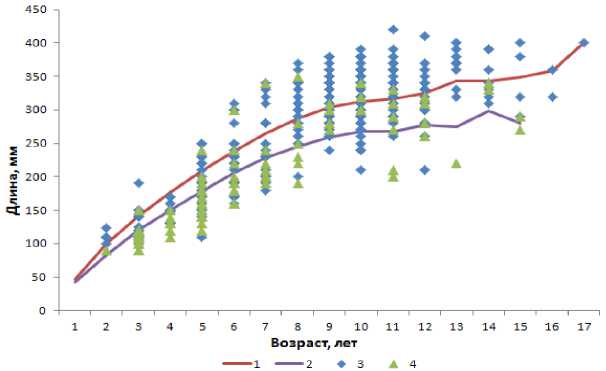
Рис. 2. Рост лиманды:
1 – самок ( N = 411 экз.), 2 – самцов ( N = 112 экз.) (по данным обратных расчислений);
3 – самок ( N = 529 экз.), 4 – самцов ( N = 144 экз.) (по данным наблюдений)
Различия в росте самок и самцов лиманды соответствовали данным, представленным в работе ( Lozan , 1989): максимальные значения возраста и длины у самок были выше, чем у самцов. Отличия в темпах роста самок и самцов отчетливее заметны по мере увеличения возраста рыб.
Согласно литературным данным ( Есипов, Сластников , 1932) максимальный возраст лиманды редко превышает 13 лет. По нашим данным, максимальный возраст лиманды Баренцева моря оценивается в 17 лет ( Стесько , 2013). Стоит отметить, что предыдущие исследования лиманды
Баренцева моря основывались на методе определения возраста по целым отолитам, при котором возможно занижение возраста рыб старших групп (Hassenger, 1991; Стесько, 2013). Кривые темпа роста самцов и самок, основанные на расчисленных данных, согласуются друг с другом несколько лучше, чем построенные по фактическим данным (R = 0,99 и R = 0,97 соответственно).
Среднее значение CV длины самок, по данным расчислений, колебалось от 10 до 24 % и в среднем составило 14 %. Наибольшие колебания длины были отмечены у рыб в возрасте 1+ и 2+ лет. С возрастом CV длины тела лиманды постепенно снижался.
Для оценки величины колебаний относительного прироста на протяжении первых пяти лет жизни были выделены четыре группы рыб в зависимости от значения относительного прироста на определенном году жизни. В первую группу вошли особи, чей относительный прирост в возрасте от двух до восьми лет находился стабильно ниже одного стандартного отклонения –1S от средней M, во вторую – в пределах от –1S до М, третья группа была представлена особями от M до +1S и четвертая – выше +1S. В результате проверки выяснилось, что лишь во второй и третьей группах нашлись единичные экземпляры, чей относительный рост был стабилен на протяжении пяти лет. Доля самок, чей относительный прирост находился в пределах ±1S, снизилась с 70 % (в возрасте двух лет) до 9 % (на восьмом году жизни). Относительный прирост в пределах ±2S наблюдался у 65 % самок в возрасте до восьми лет. Таким образом, в исследованных пробах не было обнаружено рыб со стабильно быстрым или стабильно медленным относительным приростом в течение первых пяти лет жизни, что может свидетельствовать о неустойчивости динамики роста лиманды. Подобные результаты могут быть обусловлены различиями в росте отдельных поколений, индивидуальными особенностями каждой особи, а также различными условиями роста рыбы на начальных этапах развития. Так, А.С. Шерстковым (2005) установлено, что рост лиманды Белого моря на участках с неодинаковыми условиями имеет существенные отличия. При этом не исключено, что рыбы могут совершать единовременные (раз в жизни) миграции в районы с более или менее благоприятными условиями, что впоследствии отразится на их росте.
Колебания относительного роста могут быть обусловлены особенностями нереста лиманды. В Баренцевом море эта рыба нерестится вдоль всего побережья Мурмана с мая по август, предпочитая губы и заливы ( Казанова , 1949; Бараненкова , 1952; Мухина , 2009). Длительность эмбрионального развития, сроки выклева личинок и их дальнейший рост могут оказывать влияние на интерпретацию темпа роста согласно обратным расчислениям и на определение возраста рыбы. Ранний нерест и дальнейшие благоприятные условия будут способствовать быстрому росту рыбы и, соответственно, в сравнении с другими особями ее линейные размеры будут больше. Также имеется вероятность методической ошибки: она может произойти ввиду слишком малого радиуса годового кольца или крупного – малькового, в результате чего исследователь может переоценить возраст рыбы на год или недосчитать его ( Report… , 2007; 2010).
Относительный прирост длины тела самок лиманды достаточно резко снижался в первые три года жизни, после чего падение темпа роста большинства особей замедлялось. Тем не менее относительный прирост длины тела некоторых старших рыб был выше, чем младших (рис. 3). Доля таких рыб в общей пробе была незначительна. Вероятно, ведущую роль здесь играют индивидуальные особенности их организма и трофические условия.
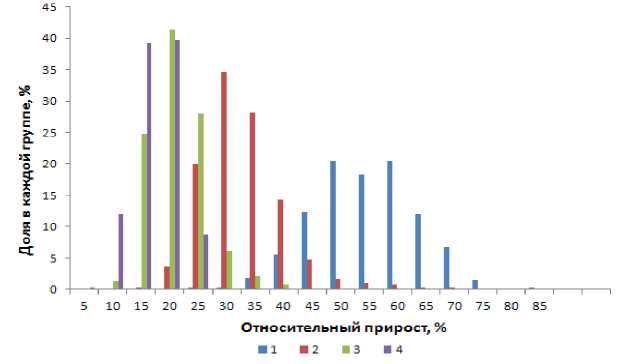
Рис. 3. Колебания относительного прироста длины тела самок лиманды в возрасте 2-5 лет по долям внутри каждой возрастной группы (по данными обратных расчислений); N = 411 экз.
На основе общепринятых моделей проводился анализ темпа роста рыбы и определялось отношение длины и массы лиманды:
– самок:
y = 0,00002 х 2,89;
– самцов:
y = 0,00001 x 2,99;
где х – длина; у – масса; при R 2 = 0,94; N = 529 и 144 экз. соответственно.
Показатель степени, близкий к трем, давал возможность использования модели Берталанфи для описания роста рыб ( Мина, Клевезаль , 1976; Шибаев , 2007).
При помощи уравнения Берталанфи на основании фактических данных были построены кривые роста самцов и самок лиманды. Значения переменных составили для самок L max = 400; K = 0,15; t 0 = 0,12; самцов – L max = 339; K = 0,17; t 0 = 0,25 (рис. 4).
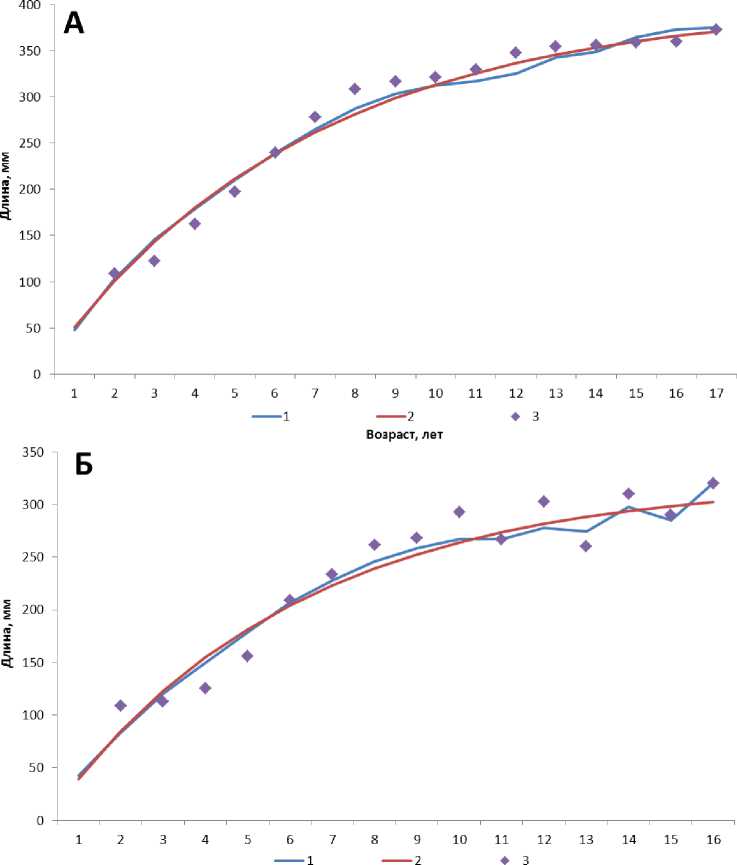
Возраст, лет
Рис. 4. Кривые роста самок (А) и самцов (Б) лиманды согласно усредненным наблюденным размерам (1), информации метода обратных расчислений (2) и результатам моделирования (3)
Коэффициент корреляции между полученными рядами данных на основе моделирования и обратных расчислений составил 0,98. Корреляция между наблюденным ростом и рассчитанными параметрами в среднем составила 0,96. Рост лиманды согласно данным проб описывался уравнением вида:
– самок:
y = 123,22 ln( x ) + 24,075;
– самцов:
y = 102,76 ln( x ) + 21,967, где х – возраст; у – длина; при R 2 = 0,99.
4. Заключение
Данные проведенных расчислений длины самок и самцов лиманды по методу "biological intercept" соответствовали наблюденному темпу роста ( R = 0,99).
Среднее значение CV длины самок согласно данным расчислений колебалось от 10 до 24 % и в среднем составило 14 %. Наибольшие колебания показателей длины были отмечены у рыб 1+ и 2+ лет.
В исследованных пробах не было обнаружено рыб со стабильно быстрым или стабильно медленным относительным приростом в течение первых пяти лет жизни, что может свидетельствовать о неустойчивости динамики роста лиманды.
Рост лиманды может быть описан при помощи модели Берталанфи.
