Результаты комплексной оценки биологических параметров в селекции сельскохозяйственных животных

Автор: Шендаков А.И.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Современные физиологические и селекционно-технологические аспекты развития животноводства
Статья в выпуске: 6 (39), 2012 года.
Бесплатный доступ
В статье приведены результаты селекции чёрно-пёстрого, симментальского и голштинского скота, а также свиней в Орловской области.
Селекция, чёрно-пёстрый, симментальский, голштинский скот, свиньи, инбридинг, мёртворождённость, трудные отёлы
Короткий адрес: https://sciup.org/147124004
IDR: 147124004 | УДК: 636.082.2:57.017.003.12
Текст научной статьи Результаты комплексной оценки биологических параметров в селекции сельскохозяйственных животных
причине мастита, С, R и X′2 – у коров с заболеваниями конечностей, С1 – у больных лейкозом. Частота антигена A1 у коров-долгожительниц составляет 0,0566, а при болезнях сердца, эндометрите и пневмонии – до 0,1818 [7]. Независимо друг от друга Г.А. Романенко [26] и Н.И. Хайруллина с соавторами [31] приводят в качестве желательного для чёрнопёстрого скота антиген G′′. В работе И.В. Кузнецовой [9] сказано, что у чёрно-пёстрых коров Рязанской области распространён аллель G 2 Y 2 E′ 3 Q′, который также часто встречается и на Урале [26]. Вместе с тем, большинство научных работ в настоящее время посвящено изучению возможности маркировки тех или иных качеств у коров (высокого удоя, долголетия, заболеваемости и пр.), остаются недостаточно исследованными эволюционные процессы в породах, маркерная способность антигенов и генетических кластеров, сходство по аллелям сложных локусов и пр.
В связи с активным внедрением в скотоводство новых методов биотехнологии [4, 6, 28, 50] исследованиями многих авторов доказано, что технологические свойства молока зависят от их генотипа по локусу гена каппа-казеина. Молоко коров с генотипом АВ и ВВ под действием сычужного фермента свёртывается раньше, чем молоко коров с генотипом АА. Используя быков, в геноме которых содержится аллель В каппа-казеина (генотип АВ и ВВ), можно не только поддерживать, но и увеличивать концентрацию аллеля В у маточного поголовья [29, 30, 32]. Однако в настоящее время недостаточно изученным остаётся вопрос изменения величины удоев при повышении концентрации гена В в стадах молочного скота.
Кроме того, в настоящее время многие зарубежные исследования посвящены изучению структуры фенотипической и генетический изменчивости, активно внедряется метод BLUP, применяется индексная селекция и подробный генетико-статистический анализ.
Таким образом, изучение биологических параметров и математическое обоснование генетических процессов в селекции животных имеет научно-практическое значение. Накопленный опыт биометрии при этом раскрывает новые возможности при изучении селекционно-генетического процесса в животноводстве.
Материалы и методы исследований
Для решения этих задач в 2004 году совместно с ОАО «Орловское» по племенной работе нами была начата комплексная работа по модернизации селекции молочного скота. Были разработаны селекционно-генетические планы для 15 ведущих хозяйств Орловской области: ОПХ «Стрелецкое» и «Красная Звезда», ЗАО «Славянское», «Берёзки» и «Куракинское», ОАО «Агрофирма Мценская», «Орловская Нива», СПК им. Мичурина и «Малиновский», ООО «Ю питер», «Маслово», «Русь», «Урицкий Агрокомплекс» и др. Была изучена молочная продуктивность коров в зависимости от генотипа и линейной принадлежности, оценены быки-производители. Воспроизводительные качества коров были изучены по 6326 отёлам в 7 хозяйствах Орловской области. Общая схема исследований представлена на рисунке 1.
Исследование биологических параметров
Генетикостатистические
r ■ r xдyм yдxм rG = i
\ r • r xдxм yдyм или
(r +r V2 xдyм yдxм rG = 1
*• r xдxм yдyм где r – фенотипические коэффициенты корреляции между признаками x и y у дочерей и матерей [25].
Дисперсионный анализ проводили с определением критерия Краскелла-Уоллиса
H _ ( E ( E xi )) , общей девиаты Dy= у ( у xi2)-H,
N факториальной девиаты
∑ ( ∑ x )2
i , силы
x
n
влияния h2x=Dx:Dy (с использованием F-критерия Фишера). Для прогнозирования поголовья свиней в России применялись уравнения регрессий, выраженные гиперболами первого и второго порядка:
Наследуемость, h2 Повторяемость, r w Изменчивость, С v , СV A Корреляции, r, r G , r x(y, z) Потенциал удоев, PS и др.
Коэффициент инбридинга, F x Частоты аллелей, р i Генетическое сходство, r i Доля тёлок и бычков, % Количество двоен , голов и пр.
bb v =« + — и y _a +--[13]. Генетический yx yx 2
xx потенциал животных был определён по формуле, приведённой Р. Ш илером с соавторами:
Способы управления биологическими параметрами
PS _ n • r w
1 - r w + n • r w
1 — r
• P x +-------- w-- A ,
1 - r w + n • r w
Рисунок 1 – Схема исследований
Коэффициенты повторяемости были определены по формуле Н. А. Плохинского:
Г 1 + Г 2 + .... + r n r w =--------------- ,
где n – число повторных циклов продуктивности, rw – коэффициент повторяемости, Px – средняя продуктивность оцениваемого животного, А – средняя продуктивность стада [49]. Генетическая вариация определялась по методике D. Houle (1992):
n
где r1, r2 и т. д. – корреляция между смежными периодами выращивания, n – количество коэффициентов корреляции. Коэффициенты наследуемости за смежные циклы продуктивности определяли по формуле Ле Руа:
CV A
V A
x
• 100 ,
h 2 n = h' • n ,1+(n-1) • rw
где h2=2b м - д . , n – количество лактаций. Были изучены множественные корреляции между селекционными признаками коров и свиней:
r x ( y , z )
( r 2 xy + r 2 « - 2 r xy r x Z r y Z )
1 -
r
2 yz
(по Г.Ф. Лакину [13]). Генетическая корреляция между промерами и удоем, удоем и содержанием жира в молоке определялась по формулам Хейзеля:
где x – фенотипическая величина среднего значения селекционного признака в стаде, V A – коэффициент аддитивной генетической изменчивости [51]. Коэффициент инбридинга (возрастания гомозиготности) вычисляли по формуле Райта-Кисловского:
Fx _ E[(1)n|+ n2-1 • (1 + f„)] • 100, где n1 и n2 – ряд, в котором находится общий предок с материнской и отцовской стороны, fa – величина коэффициента инбредного предка [25].
Величина концентрации эритроцитарных антигенов была определена как по методике Е.К. Меркурьевой [25], так и по формуле:
n й_ NA
где ni – число животных, несущих в генотипе данный антиген, N А – сумма обнаруженных антигенов у всех животных. Сходство по аллелям одного локуса
групп крови было определено по формуле Животовского: r. = у ^, где x и у - частоты аллелей данного локуса. Общее иммуногенетическое сходство было вычислено за счёт суммирования коэффициентов сходства по каждому локусу и переведено в проценты. Также была использована формула Майяла и Лингстрема [25] для вычисления общего генетического сходства:
Z x - • y j r = I = ’
V у x 2-' у У 21
где xi и yj – частоты одних и тех же аллелей (антигенов) разных учтенных локусов в двух сопоставляемых группах животных. Тестирование групп крови СП «Сабурово» проходило в лаборатории иммуногенетики ВНИИ животноводства.
Теоретическая основа исследований строилась на формуле:
y=µ+G+B+е, где y – абсолютная фенотипическая ценность, µ – среднее значение признака в популяции (стаде), G – генетические факторы, B – систематические средовые факторы, е – случайные факторы. При изучении структуры фенотипической изменчивости использовалась формула: P=G+E, где G – генотипическая изменчивость, Е – паратипическая изменчивость. Методика вычисления частоты встречаемости аллелей была построена на основе формулы Харди-Вайнберга:
(pA+qa)2=(p2AA+2pqAa+q2aa) [1, 25]
В работе были использованы каталоги быков-производителей ОАО «ЦСИО» (2001 и 2008), ОАО «Орловское (2011), а также немецкий каталог «Spermex Top Sires» (2010). Статистический анализ был проведён в компьютерных программах «Microsoft Excel» и «Статистика».
Результаты собственных исследований
Результаты скрещивания чёрно-пёстрых и палевопёстрых коров с голштинскими быками нами были опубликованы в ряде статей [8, 14-22, 33-43], из которых следует, что для производства молока оптимальной может считаться любая кровность (от 1/8 до 7/8 по голштинам) в зависимости от множества факторов – качества кормления, содержания, технологии доения и пр. Однако нами также установлено, что при увеличении в рационе коров кормовых единиц оптимальная кровность растёт: на каждые 150-200 к. ед. дополнительно в год приходится увеличение оптимальной доли голштинских генов на 12,5%, а для коров с кровностью 87,5-100% по голштинам требуется на 1500-2000 к. ед. больше, чем для низкокровного и чистопородного чёрно-пёстрого скота [43, 44, 45]. Несмотря на решённый нами вопрос об оптимальной кровности и выводе о большем влиянии отцов на генетический прогресс в популяциях крупного рогатого скота [3, 46], нами установлено, что процент генов по голштинской породе достоверно влияет на эффект селекции по удою, коррелятивные сдвиги жирности молока и живой массы [47], а также на рентабельность селекционного процесса и дополнительную прибыль [48]. Линейная принадлежность, согласно нашим исследованиям, имеет противоречивое значение, а многие линии могут по-разному показывать себя в хозяйствах в зависимости от многих факторов [27, 44]. Нами определено, что весовые коэффициенты селекционных признаков у чистопородных и помесных коров существенно отличаются [48], а процент генов голштинской породы влияет не только на аддитивную генетическую изменчивость, но и на генетические корреляции между удоем и жирностью молока [47]. В результате проведённых исследований была разработана стратегия увеличения молочной продуктивности коров Орловской области [23].
Из таблицы 1 следует, что наибольшим генетическим потенциалом удоя обладали матери канадских быков, семя которых реализовывало ОАО «Московское» в 2004 году, – 12376 кг молока при жирности 4,45%. Для немецких быков была характерна наибольшая жирность молока у матерей (4,70%). Вместе с тем, матери матерей американских быков отличались наибольшим удоем – 12175 кг при жирности 4,75%, а матери отцов у быков немецкой селекции имели самый высокий удой, что составляло 13206 кг молока жирностью 4,14%. Количество молочного жира у всех предков являлось рекордным, независимо от происхождения. Однако от 2124 дочерей немецкого происхождения в хозяйствах РФ было получено в среднем 6465 кг молока, что превысило дочерей быков американского и канадского происхождения на 150 и 1596 кг соответственно. Также они были лучшими по количеству молочного жира – 257,3 кг.
Таблица 1 – Продуктивность предков и дочерей голштинских быков ОАО « Московское »
|
Показатели |
Происхождение |
||
|
Германия |
СШ А |
Канада |
|
|
Количество быков |
24 |
14 |
8 |
|
Количество дочерей |
2124 |
754 |
749 |
|
Продуктивность матерей : Удой , кг |
11435 |
11556 |
12376 |
|
Ж ирность молока , % |
4,70 |
4,49 |
4,45* |
|
Жир , кг |
537,4 |
518,9* |
550,7 |
|
Продуктивност ь матерей матерей : Удой , кг |
10739 |
12175** |
10051* |
|
Ж ирность молока , % |
4,57 |
4,75* |
4,34* |
|
Жир , кг |
490,8 |
578,3** |
436,2* |
|
Продуктивност ь матерей отцов : Удой , кг |
13206 |
13056 |
10859*** |
|
Ж ирность молока , % |
4,14 |
3,92** |
3,99* |
|
Жир , кг |
546,7 |
511,8 |
433,3*** |
|
Продуктивность дочерей : Удой , кг |
6465 |
6315 |
4869*** |
|
Ж ирность молока , % |
3,98 |
3,83 |
3,80** |
|
Жир , кг |
257,3 |
241,9 |
185,0*** |
Примечание : *– p<0,05, **– p<0,01, ***– p<0,001 ( для всех таблиц )
Из таблицы 2 следует, что корреляция между средним удоем и жирностью молока у матерей голштинских и чёрно-пёстрых быков-производителей ОАО «ЦСИО» отечественного происхождения была выше, чем у матерей канадских быков, на 0,610 (p<0,05). Также матери матерей отечественных быков превзошли по аналогичной корреляции матерей матерей европейских быков-производителей на 0,684 (p<0,05), а, следовательно, при использовании отечественных генетических ресурсов можно более эффективно повышать жирность молока в селекции чёрно-пёстрых коров по удою.
Таблица 2 – Корреляция между удоем и жирностью молока у материнских предков голштинских и чёрно - пёстрых быков - производителей ОАО « ЦСИО »
|
Происхождение |
n |
Матери |
Матери матерей |
||
|
as 3 s О s m и g о |
S о s ч Ph о К |
as 3 s s g о C |
S Ph о |
||
|
Отечественное |
27 |
-0,247 ±0,194 |
0,212 ±0,196 |
0,025 ±0,200 |
0,279 ±0,192 |
|
Канадское |
23 |
-0,107 ±0,217 |
-0,398 ±0,200* |
0,254 ±0,211 |
-0,109 ±0,217 |
|
Европейское |
28 |
0,066 ±0,196 |
0,114 ±0,195 |
-0,373 ±0,182 |
-0,405 ±0,179* |
|
Всё поголовье |
78 |
0,097 ±0,114 |
-0,043 ±0,114 |
0,005 ±0,115 |
-0,053 ±0,114 |
Согласно расчётам по формуле Ле Руа, коэффициент наследуемости (h 2 n) удоя от 2 к 6 лактации у коров-рекордисток в Орловской области возрастал и на последнем цикле продуктивности составлял 0,154, по процентному содержанию жира в молоке и количеству молочного жира он был равен 0,829 и 0,703 соответственно (рис. 2). Это, по-видимому, объяснимо тем, что к концу продуктивного использования коровы реализовывали свой генетический потенциал максимально.
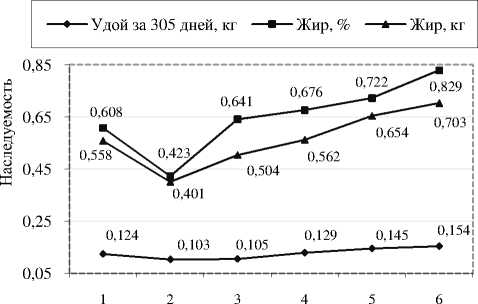
Лактации
Рисунок 2 – Коэффициенты наследуемости признаков молочной продуктивности у чёрно-пёстрых коров-рекордисток в зависимости от числа смежных лактаций, h2n
Анализ показал, что наибольшая детерминация фенотипа генотипом по удою и количеству молочного жира в ООО «Фатнево» была характерна для симментальских коров линии Флориана – 0,680 и 0,916 соответственно (рис. 3). Также была получена высокая детерминация по этим признакам у коров линий С.Т. Рокит и Мергеля. По всему поголовью она составила 0,319 и 0,316, что также можно считать положительным результатом при голштинизации палево-пёстрого скота. В хозяйстве было достаточно большое поголовье, принадлежащее разным линиям, детерминация признаков, молочной продуктивности в которых являлась достаточно высокой.
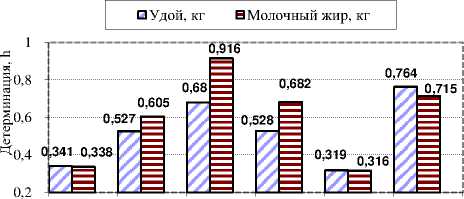
Осеано С . Т . Рокит Флориан Мергель Всё Остальные поголовье
Рисунок 3 – Детерминация признаков генотипом у коров разных линий в ООО « Фатнево » Орловской области , h
В связи с наличием высокой детерминации фенотипа генотипом в отдельных линиях хозяйства прослеживалась положительная генетическая связь между удоем и жирностью молока. Положительные коэффициенты фенотипической связи привели к высокой генетической корреляции (rG) у коров линий С. Т. Рокит, Радониса и Важного в ООО «Фатнево» – 0,507, 0,529 и 0,683 соответственно (табл. 3).
Таблица 3 – Генетическая корреляция между удоем и жирностью молока у коров разных линий в ООО « Фатнево » (n=180)
|
Линия |
Фенотипическая корреляция |
r G |
|||
|
IS CD Ph as R CD |
® Л & * * |
)S CD Ph „ P ® CD H О CL S H 57 2 >i У |
Ph н У ^ О )S g^ |
||
|
С . Т . Рокит |
0,210 |
0,031 |
0,139 |
0,183 |
0,507** |
|
Радониса |
0,876** |
0,688** |
0,410* |
0,413* |
0,529** |
|
Важного |
0,875** |
0,414* |
0,409* |
0,413* |
0,683*** |
|
Все |
0,051 |
0,020 |
0,034 |
0,011 |
0,605** |
Из таблицы 4 следует, что в ОПХ «Стрелецкое» наибольшую генетическую корреляцию имели коровы линии С. Т. Рокит – 0,804. Для животных линии У. Идеал также была характерна положительная генетическая корреляция – 0,403. Коровы линии Р. Соверинг независимо от двух отрицательных значений имели положительную генетическую связь (0,500). В то же время, для линии А. Адема была свойственна отрицательная корреляция (-0,281), а у коров родственной группы
Ривелино генетическую связь найти было невозможно из-за отсутствия аддитивного наследования.
Таблица 4 – Генетическая корреляция между удоем и жирностью молока у чёрно - пёстрых голштинизированных коров в зависимости от линейной принадлежности в ОПХ « Стрелецкое » Орловской области
|
Линия |
n |
Фенотипическая корреляция |
r G |
|||
|
I £ 8 s У c$ ° 2 4 >S 3У О |
I £ 8 s * * |
>S о Ph P p у & о |
° Ph 8" 2 ^ о )S g^ 4 |
|||
|
А . Адема |
15 |
0,303 |
0,301 |
0,023 |
-0,193 |
-0,281* |
|
Ривелино |
38 |
0,299 |
-0,102 |
0,176 |
-0,065 |
– |
|
У . Идеал |
182 |
0,195 |
0,066 |
0,029 |
0,072 |
0,403* |
|
Р . Соверинг |
212 |
0,196 |
0,068 |
-0,032 |
-0,104 |
0,500* |
|
С . Т . Рокит |
106 |
0,096 |
0,192 |
0,128 |
0,093 |
0,804** |
Примечание : Ривелино – родственная группа .
Наибольшую устойчивость удоя за завершённую лактацию в ОПХ «Стрелецкое» имели коровы с 12,5% голштинских генов – 0,594. Это превысило чистопородных животных на 0,308. Высокие показатели множественной корреляции по удою первых трёх лактаций также имели коровы с 62,5 и 75% голштинских генов – 0,412 и 0,414. Наиболее устойчивый удой за 305 дней был получен от коров с 87,5 и 37,5% генов голштинской породы – 0,648 и 0,641, что достоверно превысило контроль на 0,485 и 0,492 (табл. 5). Высокими показателями устойчивости также обладали коровы с кровностью 5/8 – 0,568, хотя при увеличении кровности до 3/4 коэффициент множественной корреляции понизился до 0,248. Полукровные коровы не отличались хорошими показателями устойчивости удоя. Наиболее устойчивая жирность молока была получена от коров с 12,5-25,0% генов голштинской породы (r x(y,z) =0,698-0,568), что значительно превысило показатель множественной корреляции других помесных генотипов, за исключением коров с 87,5% генов голштинов, которые отличались невысокими удоями. По устойчивости живой массы точных тенденций получено не было, однако наибольшими показателями обладали чистопородные коровы, а также помеси с кровностью 37,5, 62,5 и 87,5% генов голштинской породы.
Таблица 5 – Множественная корреляция между признаками первых трёх лактаций у чёрно - пёстрых коров в зависимости от генотипа в ОПХ « Стрелецкое »
|
%HF |
n |
Селекционные признаки коров , r1 ( 2,3 ) ±mr |
||
|
Удой за 305 дней , кг |
Жир , % |
Ж ивая масса , кг |
||
|
0 |
12 |
0,156±0,329 |
0,541±0,280 |
0,999±0,015 |
|
12,5 |
13 |
0,485±0,276 |
0,698±0,226 |
0,471±0,280** |
|
25,00 |
57 |
0,341±0,128 |
0,568±0,112 |
0,486±0,119** |
|
37,50 |
27 |
0,641±0,157* |
0,143±0,202 |
0,779±0,125 |
|
50,00 |
63 |
0,249±0,125 |
0,051±0,129* |
0,464±0,114** |
|
62,50 |
44 |
0,568±0,129* |
0,286±0,150 |
0,623±0,122 |
|
75,00 |
46 |
0,248±0,148 |
0,248±0,148 |
0,032±0,378*** |
|
87,50 |
10 |
0,648±0,437* |
0,437±0,340 |
0,776±0,238 |
Примечание : HF – % генов голштинской породы (0% по голштинам – контрольная группа ).
Из таблицы 6 следует, что наибольшую устойчивость удоя за 305 дней лактации имели коровы родственной группы Ривелино – 0,367. Это незначительно превысило коров линии А. Адема. Наиболее устойчивая жирность молока была получена от коров линии С. Т. Рокит (rx ( y,z ) =0,702), а по количеству молочного жира все голштинские линии имели преимущество над чистопородными коровами на 0,157-0,247.
Таблица 6 – Множественная корреляция между признаками первых трёх лактаций у чёрно - пёстрых коров разных линий в ОПХ « Стрелецкое »
|
Линия |
Селекционные признаки коров , r1(2,3)±mr |
||
|
Удой за 305 дней , кг |
Жир , % |
Молочный жир , кг |
|
|
А . Адема |
0,305±0,287 |
0,528±0,256 |
0,102±0,300 |
|
Ривелино |
0,367±0,129 |
0,374±0,129 |
0,259±0,134 |
|
У . Идеал |
0,326±0,097 |
0,168±0,101* |
0,349±0,096 |
|
Р . Соверинг |
0,288±0,114 |
0,379±0,110 |
0,260±0,115 |
|
С . Т . Рокит |
0,219±0,252 |
0,702±0,184 |
0,273±0,248 |
Примечание : Ривелино – родственная группа .
Согласно нашим опубликованным данным [48], независимо от лактации в стаде коров ОПХ «Стрелецкое» проявлялись высокие множественные корреляции удоя с жирностью молока и живой массой – до 0,732-0,811 (p<0,05). Дисперсионный анализ этих данных (рис. 4) подтверждает, что % генов чёрнопёстрой голштинской породы достоверно влиял на множественные коэффициенты корреляции между удоем, жирностью молока и живой массой у коров – 31,08±7,65% (α<5%). Лактация не имела сильного влияния на коэффициенты, поскольку был исключен фактор выбраковки (2,16±10,86%). Это позволило нам сделать предположение о том, что совместное влияния % голштинских генов и лактации (37,21±6,97%) объяснимо усилением действия генотипа с возрастом. К неучтённым факторам в данном случае можно отнести паратипические факторы.
□ Сила влияния факторов , % a Ошибка силы влияния , %
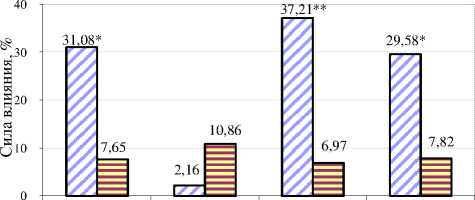
% генов лактация % генов - лактация неучтённые
Факторы
Рисунок 4 – Сила влияния факторов на множественные корреляции между удоем , жирностью молока и живой массой , % (* – α <5%, ** – α <1%)
Принято считать, что в хозяйствах рождается 50% тёлок и 50% бычков, однако, на практике даже без применения разделённого по полу семени распределение полов может быть неравномерным, а в иностранных моделях NYB и NPB [11] при определении необходимого количества быков-производителей в популяциях учитывается вероятность рождения не только телёнка, но и тёлки. Интересные данные приводят ирландские учёные S. Mc. Parland, J. F. Kearney, M. Rath и D. P. Berry в журнале «Dairy Science» (2007), где сказано, что у коров с коэффициентом возрастания гомозиготности (Fx) от 20 до 30% рождается на 3-6% больше тёлок в сравнении c аутбредными матерями [52]. Данное явление учёные объясняют теорией Trivers и Willard (1973), согласно которой ослабленным инбредной депрессией матерям проще выносить тёлку.
Из данных таблицы 7 следует, что у аутбредных коров СПК имени Мичурина, ЗАО «Куракинское» и ОПХ «Красная Звезда» рождается от 41,07 до 49,47% тёлок, от 1,13 до 6,41% телят рождаются мёртвыми. Коровы, у которых были отцами Падеграс, Миг и Дон дали 11 двоен – 6, 1 и 4 соответственно. У аутбредных коров ООО «Фатнево» было получено тёлок на 3,91% больше нормы. В ОПХ «Стрелецкое», согласно данным хозяйства, в 2008 году на 893 живых телёнка было получено всего 289 тёлок (или 32,36%). Следует также заметить, что применение инбридинга в степенях от 1,5626 до 25% в чёрно-пёстром поголовье способствовало выравниванию распределения полов в потомстве (49,66% тёлок), а в симментал-голштинском поголовье коров (Fx=0,0488-12,5%) вело к некоторому ослаблению вероятности рождения тёлок, однако у 9 коров с Fx=6,25-12,5% из 22 отёлов родилось на 2 тёлки больше, чем бычков. Также инбридинг способствовал в ООО «Фатнево» получению 5 двоен, или 4,88% от количества полученных телят. По всему чёрно-пёстрому поголовью из 4966 живых телят было получено 43,62% тёлок, в изученном симментальском поголовье и его помесях было получено приблизительно одинаковое количество бычков и тёлок, а из 6074 живых телят в исследуемых хозяйствах 44,85% составили тёлки, рождение мёртвых телят на 6326 отёлов занимало 3,92%, или 248 голов.
Таблица 7 – Воспроизводительные качества чёрно - пёстрого и симментальского голштинизированного скота в хозяйствах Орловской области
|
Всего изучено отёлов |
Ж ивых телят , голов |
Бычков , голов |
Тёлок , голов (%) |
Количество двоен |
Аборты |
Мёртворожденных |
Трудные отёлы |
||
|
голов |
% ко всем отёлам |
||||||||
|
Аутбридинг ( неродственное спаривание ) ЗАО « Славянское » ( чёрно - пёстрый и симментальский , Верховский район ) |
|||||||||
|
645 |
622 |
348 |
274 (44,05) |
– |
5 \ |
23 \ |
3,57 |
– |
|
|
СПК им . Мичурина ( чёрно - пёстрый , Верховский район ) |
|||||||||
|
1150 |
1137 |
670 \ |
467 (41,07) |
11 \ |
1 |
13 \ |
1,13 |
6 |
|
|
ЗАО |
Куракинское » ( чёрно - пёстрый , Свердловский район ) |
||||||||
|
811 |
759 |
389 |
370 (48,75) |
– |
– |
52 \ |
6,41 |
3 |
|
|
ОПХ |
« Красная Звезда |
» ( чёрно - пёстрый , Орловский район ) |
|||||||
|
1307 |
1225 |
619 \ |
606 (49,47) |
– |
3 \ |
82 \ |
6,27 |
8 |
|
|
ООО « Фатнево » ( симментальский , Болховский район ) |
|||||||||
|
619 |
588 |
271 |
317 (53,91) |
3 \ |
4 \ |
31 \ |
5,01 |
24 |
|
|
Без учёта степени родств а ОПХ « Стрелецкое » ( чёрно - пёстрый , Орловский район , по данным хозяйства ) |
|||||||||
|
925 |
893 |
604 |
289 (32,36) |
– |
18 \ |
32 \ |
3,46 |
– |
|
|
ООО « Ш аблыкинский Агрокомплекс » ( чёрно - пёстрый , Ш аблыкинский район ) |
|||||||||
|
499 |
498 |
272 |
226 (45,38) |
– |
– |
1 |
0,20 |
– |
|
|
Инбридинг ( родственное спаривание ) Чёрно - пёстрый скот ( СПК имени Мичурина , ЗАО « Куракинское » и |
ОПХ « Красная Звезда ») |
||||||||
|
153 |
147 |
74 |
73 (49,66) |
– |
– |
6 |
1,31 |
1 |
|
|
Симментальский ( ООО « Фатнево ») |
|||||||||
|
213 |
205 |
103 \ |
102 (49,76) |
5 \ |
1 |
8 |
3,76 |
14 |
|
|
По всем изученным хозяйствам области ( без учёта типа подбора ) |
|||||||||
|
Чёрно - пёстрый скот |
|||||||||
|
5168 |
4966 |
2800 |
2166 (43,62) |
11 \ |
27 |
202 |
3,91 |
18 |
|
|
Симментальский |
|||||||||
|
1154 |
1108 |
550 |
558 (50,36) |
8 \ |
6 |
46 |
3,99 |
38 |
|
|
Всё изученное поголовье |
|||||||||
|
6326 |
6074 |
3350 |
2724 (44,85) |
19 \ |
32 \ |
248 |
3,92 |
56 |
|
С увеличением доли генов голштинской породы до 50% (n=1224) в стадах Орловской области возрастало количество мёртворожденных телят – от 1,97 до 5,96% (см. рисунок 5). В дальнейшем у адаптированных к условиям России коров с кровностью 87,5% по голштинам наблюдалось уменьшение количества мёртворожденных телят 58
(1,96%), однако у завезённых из Германии, Венгрии и Ирландии чистопородных голштинов, не представленных на рисунке, проходили сложные процессы адаптации и наблюдались худшие воспроизводительные способности в сравнении с местным скотом.
I -___I Мёртворожденных, %
Полиномиальная (Мёртворожденных, %)
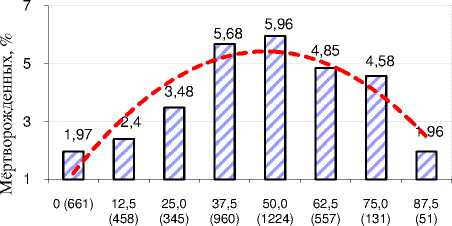
% генов голштинской породы (всего отёлов)
Рисунок 5 – Влияние голштинской породы на число мёртворожденных телят в хозяйствах Орловской области (% ко всем отёлам )
Наряду с рождением мёртвых телят, % генов голштинской породы повлиял на процентное соотношение полов в потомстве (рис. 6). Так, при возрастании кровности от 12,5 до 62,5% по голштинам у коров в хозяйствах области процент рождения тёлок возрастал с 40,06 до 50,0%, а затем прослеживалась тенденция к увеличению вероятности рождения бычков, то есть вероятность рождения тёлок следовала зависимости, которая может быть выражена уравнением параболы второго порядка. Возможно, данная тенденция связана с тем, что коровы с кровностью 12,5% генов по голштинам и чистопородные голштины имеют большую живую массу, а полукровные коровы являются средними по многим продуктивным показателям.
Тёлок, % Бычков,%
Полиномиальная (Тёлок, %) Полиномиальная (Бычков,%)
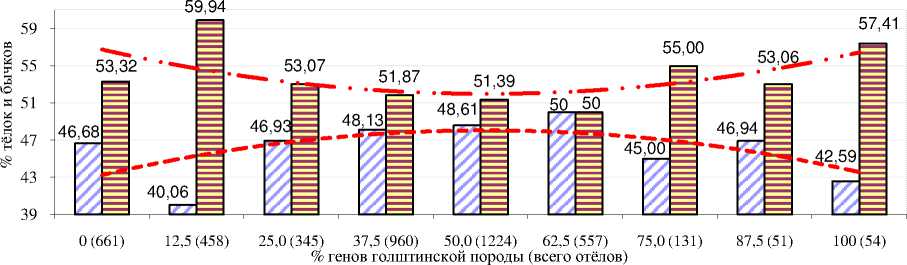
Рисунок 6 – Распределение полов в зависимости от процента генов голштинской породы в хозяйствах Орловской области
Также следует заметить, что быки-производители давали разное соотношение полов в потомстве ОПХ «Стрелецкое» (рис. 7).
№1093 (на проверке)
№2205 (на проверке)
№4427 (на проверке)
№4345 (на проверке)
Мороз 637 о
Люкс 21 ю s Хан 1876
s
3 Чек 50
у s s Мейсон 5091
Вальтер 739
Лидер 129
Стаж 735
^^^^^<
47,5
zzzzzzzzzzzzzzzzzz#
18,18
7,41
56,34
22222222222222222222^
fZZZZZZZZZ,
\T//Z////^
32,14
25,86
\f////////z
V/ZZ//Z
31,34
24,19
vzzzzzzzzz\ fZZZZZZZZZZ
33,1
34,43
61,54
TZZZ^
20,59
5 15 25 35 45 55 65
% тёлок
Рисунок 7 – Влияние быков - производителей на вероятность рождения тёлок в ОПХ « Стрелецкое »
Так, у большинства быков ОАО «ЦСИО», семя которых использовалось в хозяйстве, вероятность рождения тёлки составляла от 20,59 до 34,43%, у быков ОАО «Невское» (без кличек), которые находились на проверке в хозяйстве, процент рождения тёлок составил от 7,41 до 56,34%.
Только у двух быков рождались преимущественно тёлки (Мороз 637 и №2205). Это позволяет сделать вывод о возможности увеличения поголовья тёлок в хозяйствах за счёт использования семени отдельных быков, являющихся улучшателями по молочной продуктивности дочерей. Также от быка Лидера 129 в 2008 году было рождено 10 мертвых телят, от Стажа 735 и Вальтера 739 – по 4, от Мейсона 5091 – 2, от ленинградских быков №2205 и №1093 – 4 и 5 соответственно. Из 18 случившихся в хозяйстве абортов 5 приходилось на семя Мейсона 5091, по 3 на быков №4345 и Вальтера 739. При использовании семени Хана 1876, Люкса 21 и Мороза 637 не было ни абортов, ни мёртворожденных телят.
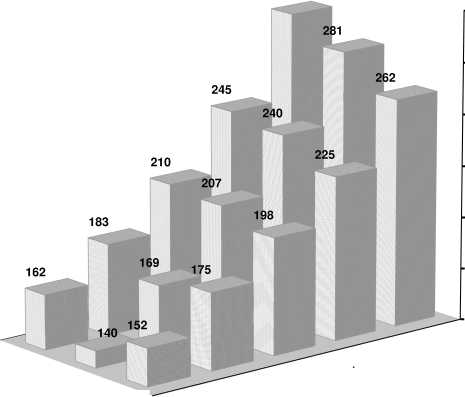
Из таблицы 8 следует, что с 2001 по 2008 год у красно-пёстрых голштинских быков-производителей ОАО «ЦСИО» добавилось 16 эритроцитарных антигенов, 3 антигена выбыло, у чёрно-пёстрых голштинских быков добавилось 14 антигенов, выбыло 4. За тот же период времени при сравнении голштинов разной масти между собой было обнаружено 3 антигена, характерных для краснопёстрых животных, и 22 антигена, характерных для чёрно-пёстрых животных. При этом чёрно-пёстрые голштины были значительно разнообразнее по иммуногенетическим особенностям и имели наибольшее количество эритроцитарных антигенов в сравнении с изученными породами.
Таблица 8 – Динамика иммуногенетического полиморфизма у голштинских быков ОАО « ЦСИО »
|
Порода |
Антигены |
|||
|
Отсутствующие |
Изменения с 2001 года |
Характерные |
||
|
Добавившиеся |
Выбывшие |
|||
|
Красно - пёстрые голштины |
H, D, Z’, G 1 , G 3 , O 2 , O 5 , Q 1 , O x , Q 2 , T 2 , Y 1 , D' 1 , D' 2 , E' 2 , I' 1 , I' 2 , J 2 , K', O' 2 , P', Y', G'' 1 , G'' 2, A'' 2 , B'', D'', O'', Q'', W 1 , W 2 , X', C'' 1 , C'' 2 , F 1 , F 2 , V 1 , V 2 , N', M 2 , S 2 , U 1 , U 2 , U' 1 , U' 2, R', T', Y, E 3 , P' 2 , M, G, E', S, U'', O, Q' 1 , J' 1 , J' 2 , I, F' |
A 2 , B 1 , G 2 , I 1 , I 2 , O 3 , O 4 , P 2 , Q, T 1 , C 2 , C'', R 1 , V, L', G'' |
O 2 , J' 1 , K' |
O 3 , P 2 , T 1 |
|
Чёрно - пёстрые голштины |
H, D, G 1 , O 3 , P 2 , T 1, O 5 , Q 1 , O x , Q 2 , T 2 , D' 1 , D' 2 , I' 1 , I' 2 , J 2 , O' 2 , P', Y', G'' 2, A'' 2 , B'', D'', O'', Q'', W 1 , W 2 , X', C'' 1 , C'' 2 , F 1 , F 2 , V 1 , V 2 , N', M 2 , S 2 , U 1 , U 2 , U' 1 , U' 2, R', T' |
Z', G 3 , C', Y, E 3 , P' 2 , M, G, E', O, J' 1 , J' 2 , I, F' |
G 1 , T 1 , Y', J 2 |
Z’, G 3 , O 2 , Y 1 , E' 2 , K', G'' 1 , U 1 , Y, E 3 , P' 2 , M, G, E', S, U'', O, Q' 1 , J' 1 , J' 2 , I, F' |
Общее иммуногенетическое сходство было максимальным между холмогорскими и чёрнопёстрыми голштинскими быками «ЦСИО» (0,913) в связи со скрещиванием этих пород, между чёрнопёстрыми голштинскими и чёрно-пёстрыми быками (0,902), а также между голштинами разной масти в связи с родственным происхождением (0,901) [42].
Нами также получены данные, в которых маркерная способность эритроцитарных антигенов возрастала с увеличением частоты встречаемости от 0,02 до 0,04. При этом наибольшим удоем обладали дочери чёрно-пёстрых голштинских быков при частоте встречаемости антигенов больше 0,03 – 5194 кг молока, наблюдалось увеличение удоев у коров с частотой встречаемости антигенов ниже 0,01 – 4601 кг молока по первой лактации. Это, по-видимому, объяснимо тем, что в популяции чёрно-пёстрого голштинизированного скота России в последние годы шло активное внедрение эритроцитарных антигенов, маркирующих высокий удой.
У чёрно-пёстрых голштинских быков, семя которых реализовывало ОАО «Орловское» в последние годы, 71 раз встречался антиген F, 30 – H', 25 – Х 2 , 24 – W, 23 – Q', 20 – А 1 , 18 – E, 17 – Z, 14 – Y 2 , Е' 1 и С 1 , 13 – G 2 , 11 – L и I 2 , 10 – O 1 , 9 – S 1 и C 2 . При этом 34 антигена встречались от 2 до 5 раз, 24 – всего по одному разу. Это подтверждает активное внедрение в поголовье коров Орловской области разнообразных аллелей. Наиболее удачными при этом были быки с иммуногенетическим сочетанием B 1 G 2 K – до 6578 кг молока и 246 кг молочного жира от 26 дочерей при оценке по качеству потомства.
Комплекс исследований, проведённых нами в отрасли свиноводства, позволяет обобщить несколько важных выявленных закономерностей:
-
1) в отечественном поголовье генетический потенциал многоплодия у свиноматок можно выразить уравнением нелинейной регрессии: yx=9,421+0,62058 x -0,0557x2 (где х – номер опороса), при этом наибольшее многоплодие характерно к 5-6 опоросу – до 11,1 поросёнка в среднем, после чего следует понижение до 10,05 голов к 10 опоросу;
-
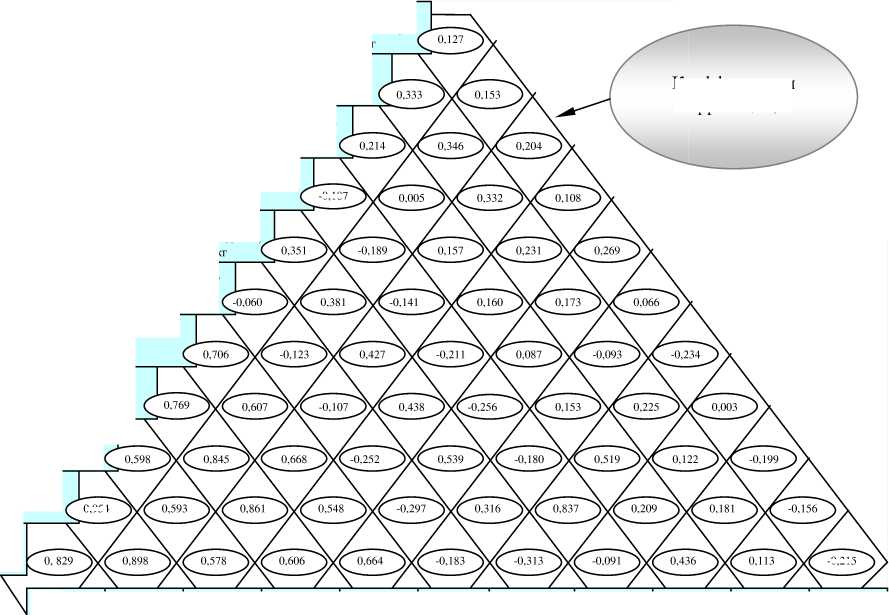
2) у свиней с развитыми мясосальными качествами (рис. 8) прослеживалась наибольшая зависимость живой массы от обхвата груди (r=0,954) и длины туловища (r=0,829), в то время как между живой массой, обхватом груди и длиной туловища множественная корреляция (r x(y,z) ) составляла 0,955, а живую массу ( х ) можно было прогнозировать по
обхвату груди ( y ) и длине туловища ( z ) по формуле: xy,z = -30,0938-1,8143y + 3,4303z (рис. 9);
-
3) на сегодняшний день, выразив 1991 год первым годом кризиса ( х ), для прогноза поголовья свиней в России можно вывести две формулы, одна из которых является гиперболой первого порядка: у = 15833 + 23,3 x x (при продолжении спада поголовья), вторая: V = 19 24 + 19,007 , где х — год от начала кризиса (при yx , x 2
ослаблении спада поголовья). Это означает, что при сохранении тенденций отечественного свиноводства к 2040 году в России будет от 16,3 до 19,4 млн. голов свиней. Однако в случае увеличения поголовья в ближайшие 5-6 лет можно будет применять для построения уравнения прогноза параболу третьего порядка ( yx=a+bx+cx2+dx3) , а, следовательно, при развитии отрасли к 2025 году можно будет получить до 23-25 млн. голов свиней.
Ретроспективный анализ , проведённый нами, показал, что наименьшая общая фенотипическая изменчивость (C v ) у свиноматок каликинской породы была характерна для количества сосков, высоты в холке, длины туловища и многоплодия – 3,1, 4,8, 4,1 и 4,9% соответственно, а наибольшая – для живого веса свиноматок и количества поросят при отъёме (14,0 и 17,2% соответственно). При этом σ по промерам колебалась от 3,4 см (глубина груди) до 9,9 см (обхват груди), по живому весу свиноматок составила 27,5 кг. В стаде ГПЗ им. Георгиевского за последние 10 лет отбора и селекционной работы живая масса ливенских хряков осталась приблизительно на прежнем уровне и в 2007 году составила 260,4 кг (n=42). При этом некоторое уменьшение их живой массы (на 2,1 кг) в сравнении с 2000 годом компенсировалось развитием мясных качеств и длины туловища (на 3,1 см). Проявлялась тенденция к возрастанию баллов за телосложение – с 90,7 до 91,2, несколько снизилась эффективность случек – с 84,0 до 81,4% (-2,6%). Также наметилась тенденция увеличения многоплодия у дочерей – с 9,73 до 10,01 поросёнка, однако при этом уменьшилось среднее количество поросят в 2 месяца (с 9,33 до 8,64 голов), что компенсировалось увеличением среднего веса 1 поросёнка в 2 месяца на 1,34 кг. Однако следует заметить, что отечественное поголовье свиней, разводимых в ЦФО, существенно уступает по мясным качествам импортным породам.
Взаимосвязанные признаки
Плодовитость
Число сосков
Вес поросёнка при рождении,
Число поросят при отъёме
Вес поросёнка при отъёме,
Глубина груди, см
Число поросят в 1 месяц
Вес гнезда в 1 месяц, кг
Обхват груди, см
Высота в холке, см
Длина туловища, см
-0.187
0,954
-0,215
Плодовитость
Число сосков
Живой вес, кг
Высота в холке, см
Число поросят при отъёме
Обхват груди, см
Глубина груди, см сло сят в 1
сяц
Вес гнезда в 1 месяц , кг
Длина туловища, см
Вес поросёнка при рождении, кг
Рисунок 8 – Коэффициенты корреляции между селекционными признаками каликинской породы свиней , r

оэффициенты орреляции , r
Ж ивая масса,
Длина туловища, см
140 см
150 см
150 см
170 см
160 см
140 см
160 см 130 см
кг
уди, см
Рисунок 9 – Зависимость живой массы свиноматок от обхвата груди и длины туловища , кг
Также, применяя селекцию по генам в молочном скотоводстве, мы выяснили, что 18 швицких быков немецкого происхождения (47432 дочери) с генотипом BB по гену каппа-казеина имели племенную ценность +699 кг молока и +24,5 кг молочного белка. По результатам оценки дочерей это превышало быков с генотипом АА на 103 кг молока и 2,5 кг молочного жира. Концентрация генотипа BB у швицких быков немецкого происхождения составила 0,474, а аллеля В по данному гену – 0,685.
Средний генетический потенциал чёрно-пёстрых коров в Орловской области (PS) достигал 6500-8000 кг молока, симменталов – 5000-5500 кг, голштинов – до 10000-11000 кг молока за 305 дней лактации. При этом в 2010 году в области насчитывалось более 1000 коров с удоями выше 7000 кг молока и 90 коров с удоями выше 10000 кг молока. В общей генетической изменчивости селекционных признаков на долю аддитивной изменчивости приходилось до 75-90%, на случайные генетические эффекты – 10-25%, при скрещивании неаддитивная изменчивость возрастала.
Таким образом, использование в селекции сельскохозяйственных животных подробных методов генетико-статистического анализа позволяет выявить многие скрытые процессы, протекающие в стадах, а также важные для теории науки и практики животноводства биологические закономерности. В России и Орловской области существуют предпосылки увеличения молочной продуктивности коров в связи с наличием достаточно высокой генетической изменчивости селекционных признаков. Наличие положительной генетической корреляции между селекционными признаками может позволить быстрое увеличение целого ряда продуктивных показателей в стадах молочных коров. В области намечены резервы увеличения поголовья тёлок и мясной продуктивности свиней. Проведённые исследования в перспективе позволят ускорить работу по выведению нового типа скота и начать модернизацию селекции в свиноводстве Орловской области, включая применение BLUP.
Теоретический и научно - практический журнал . Основан в 2005 году
Учредитель и издатель : Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования « Орловский государственный аграрный Университет »
Адрес редакции: 302019, г. Орел, ул. Генерала Родина, 69. Тел.: +7 (4862) 45-40-37 Факс: +7 (4862) 45-40-64
Свидетельство о регистрации ПИ №ФС77–21514 от 11.07.2005 г.
Сдано в набор 12.11.2012 г. Подписано в печать 29.11.2012 г. Формат 60х84/8. Бумага офсетная.
Гарнитура Таймс.
Объём 12,5 усл. печ. л.
Тираж 300 экз. Издательство Орел ГАУ, 302028, г. Орел, бульвар Победы, 19. Лицензия ЛР №021325
от 23.02.1999 г.
Ж урнал рекомендован ВАК Минобрнауки России для публикаций научных работ, отражающих основное научное содержание кандидатских и докторских диссертаций
Список литературы Результаты комплексной оценки биологических параметров в селекции сельскохозяйственных животных
- Бакай, А.В. Генетика [Текст]/А.В. Бакай, И.И. Кочиш, Г.Г. Скрипниченко. -М.:КолосС, 2007. -448 с
- Букаров, Н. Новый уровень познания маркерных генов групп крови у скота [Текст]/Н. Букаров, Е. Лебедев, И. Морозов//Молочное и мясное скотоводство. -2005. -№ 7. -С. 39-41
- Буяров, В.С. Эффективность селекции молочного скота [Текст]/В.С. Буяров, А.И. Шендаков, Т.А. Шендакова//Животноводство России. -2011. -№ 1. -С. 41-44
- Генджиева, О.Б. Изучение генетического разнообразия калмыцкого скота с использованием ISSR-фингерпринга [Текст]/О.Б. Генджиева, Г.Е. Сулимова//Зоотехния. -2009. -№ 3. -С. 4-5
- Исламова, С. Порода и антигенный состав крови быков-производителей [Текст]/С. Исламова, Ф. Исламов//Молочное и мясное скотоводство. -2006. -№ 5. -С. 34-35
- Исламова, С. Применение ДНК-технологии в селекции [Текст]/С. Исламова, Ф. Исламов, И. Долматова //Молочное и мясное скотоводство. -2007. -№ 5. -С. 2-4.
- Коровушкин, А.А. Иммуногенетические маркеры устойчивости коров к заболеваниям [Текст]/А.А. Коровушкин//Зоотехния. -2004. -№ 6. -С. 9-11
- Крюков, В.И. Совершенствование симменталов в СПК «Фатневский» [Текст]/В.И. Крюков, А.И. Шендаков, В.М. Николина//Зоотехния. -2004. -№ 6. -С. 11
- Кузнецова, И.В. Мониторинг генетической структуры популяции крупного рогатого скота чёрно-пёстрой породы [Текст]/И.В. Кузнецова//Зоотехния. -2009. -№ 2. -С. 2-3
- Кузнецов, В.М. Основы научных исследований в животноводстве [Текст]/В.М. Кузнецов. -Киров, Изд. Зонального НИИСХСВ. -2006. -568 стр
- Кузнецов, В.М. Разработка оптимальных программ селекции в молочном скотоводстве/В.М. Кузнецов//Зоотехния. -1996. -№ 1. -С. 5-13
- Кузнецов, В.М. Современные методы анализа и планирования селекции в молочном стаде [Текст]./В.М. Кузнецов -Киров, Изд. Зонального НИИСХСВ. -2001. -116 с
- Лакин, Г. Ф. Биометрия [Текст]/Г. Ф. Лакин. -М.: Высшая школа. -1990. -352 с.: ил
- Ляшук, Р.Н. Совершенствование чёрно-пёстрого скота в Орловской области [Текст]/Р.Н. Ляшук, А.И. Шендаков, М.В. Востров//Молочное и мясное скотоводство. -2005. -№ 7. -С. 20-22
- Ляшук, Р.Н. Совершенствование молочного скота в Орловской области [Текст]/Р.Н. Ляшук, А.И. Шендаков, М.В. Востров//Молочное и мясное скотоводство. -2007. -№ 1. -С. 22-26
- Ляшук, Р.Н. К вопросу о голштинизации чёрно-пёстрого скота в Орловской области [Текст]/Р.Н. Ляшук, А.И. Шендаков, М.В. Востров, В.В. Сорокин//Вестник Орёл ГАУ. -2007. -№ 1. -С. 38-41
- Ляшук, Р.Н. Результаты селекции молочного скота в Орловской области [Текст]/Р.Н. Ляшук, А.И. Шендаков//Аграрная наука. -№ 9. -2007. -С. 25-27
- Ляшук, Р.Н. Повышение генетического потенциала молочного скота [Текст]/Р.Н. Ляшук, А.И. Шендаков, М.В. Востров, В.В. Сорокин//Зоотехния. -2007. -№ 11. -С. 3-6
- Ляшук, Р.Н. Зоотехнические аспекты реализации продуктивного потенциала голштинизированного чёрно-пёстрого скота [Текст]/Р.Н. Ляшук, А.И. Шендаков, В.В. Сорокин, Е.Г. Амелин.//Аграрная наука.- 2008. - №2. - с. 21-22.
- Ляшук, Р.Н. Селекционно-генетическая оценка быков-производителей по потенциалу молочной продуктивности [Текст]/Р.Н. Ляшук, А.И. Шендаков, Т.А. Шендакова//Сельскохозяйственная биология. -2008. -№ 4. -С. 23-29
- Ляшук, Р.Н. Совершенствование системы отбора молочного скота [Текст]/Р.Н. Ляшук, А.И. Шендаков, Т.А. Шендакова, В.В. Машкей//Аграрная наука. -2008. -№ 8. -С. 31-34
- Ляшук, Р.Н. Повышение генетического потенциала молочного скота [Текст]/Р.Н. Ляшук, А.И. Шендаков, В.В. Сорокин//Зоотехния. -2009. -№ 3. -С. 2-3
- Ляшук, Р.Н. Основные направления развития молочного скотоводства в Орловской области [Текст]/Р.Н. Ляшук, В.Н. Масалов, А.И. Шендаков//Вестник Орёл ГАУ. -2011. -№ 1. -С. 9-13
- Ляшук, Р.Н. Селекционно-генетический потенциал импортированного молочного скота [Текст]/Р.Н. Ляшук, А.И. Шендаков, А.В. Сурженков//Вестник Орёл ГАУ. -2010. -№ 2. -С. 30-32
- Меркурьева, Е.К. Генетика [Текст]/Е.К. Меркурьева, З.В. Абрамова, А.В. Бакай и [др.] -М.: Агропромиздат. -1991. -446 с.: ил
- Романенко, Г.А. Генетические маркеры в селекции уральского чёрно-пёстрого скота [Текст]/Г.А. Романенко//Аграрный вестник Урала. -2009. -№ 4. -С. 82-83
- Самусенко, Л.Д. Разведение симментальского скота по линиям в Орловской области [Текст]/Л.Д. Самусенко, А.И. Шендаков//Зоотехния. -2009. -№ 6. -С. 2-4
- Терлецкий, В.П. Оценка племенных животных по полиморфизму генов и ДНК/В.П. Терлецкий, Н.В. Дементьева, Е.С. Усенбеков//Зоотехния. -2001. -№ 1. -С. 14-16
- Тинаев, А. Племенные ресурсы быков-производителей чёрно-пёстрой породы/А. Тинаев, Л. Калашникова, Т. Ганченкова [Текст]//Молочное и мясное скотоводство. -2009. -№ 1. -С. 5-7
- Тинаев, А. Хозяйственно-полезные признаки чёрно-пёстрого скота с разными генотипами каппа-казеина [Текст]/А. Тинаев, Л. Калашникова, К. Аджибеков//Молочное и мясное скотоводство. -2005. -№ 5. -С. 30-32
- Хайруллина, Н. Влияние сочетания локусов эритроцитарных антигенов быков-производителей на структуру стада крупного рогатого скота/Н. Хайруллина, Н. Фенченко, З. Ярмухаметова, Р. Ахмадуллин//Зоотехния. -2007. -№ 6. -С. 5-6
- Черных, А. Генотип каппа-казеина и качество молока чёрно-пёстрых коров/А. Черных, Л. Калашникова [Текст]//Молочное и мясное скотоводство.-2008. -№ 5. -С. 9-10
- Шендаков, А.И. Молочная продуктивность симментал-голштинских коров [Текст]/А.И. Шендаков//Молочное и мясное скотоводство. -2002. -№ 2. -С 16-17
- Шендаков, А.И. Влияние генотипа коров на сыропригодность молока [Текст]/А.И. Шендаков, В.Н. Данилов, В.И. Крюков//Молочное и мясное скотоводство. -2003.-№ 8. -С. 16-18
- Шендаков, А.И. Сыропригодность молока симментальских коров и помесей [Текст]/А.И. Шендаков, В.И. Крюков, В.Н. Данилов//Зоотехния. -2004. -№ 2. -С. 29-30
- Шендаков, А.И. Совершенствование симментальского скота в Орловской области [Текст]/А.И. Шендаков, В.И. Крюков//Молочное и мясное скотоводство. -2004. -№ 7. -С. 10-11
- Шендаков, А.И. Использование потенциала голштинского скота [Текст]/А.И. Шендаков//Зоотехния. -2005. -№ 8. -С. 5-7
- Шендаков, А.И. Устойчивость признаков у симментал-голштинских коров [Текст]/А.И. Шендаков//Зоотехния. -№ 7. -2006. -С. 4-5
- Шендаков, А.И. Совершенствование симментальского скота в Орловской области [Текст]/А.И. Шендаков, В.И. Крюков//Зоотехния. -№ 7. -2007. -С. 4-6
- Шендаков, А.И. Состав, качество и сыропригодность молока коров в зависимости от генотипа [Текст]/А.И. Шендаков, В.И. Крюков//Известия Орловского ГТУ, серия «Легкая и пищевая промышленность». -2003. -№ 3-4
- Шендаков, А.И. Модернизация селекции в молочном скотоводстве Орловской области [Текст]/А.И. Шендаков//Молочное и мясное скотоводство.-2008. -№ 6. -С. 15-19
- Шендаков, А. И. Генетические аспекты модернизации молочного скотоводства [Текст]/А. И. Шендаков, Т. А. Шендакова//Вестник Орёл ГАУ. -2009. -№2. -С. 30-35
- Шендаков, А.И. Результаты использования потенциала голштинского скота в Орловской области [Текст]/А.И. Шендаков//Зоотехния. -2010. -№ 2. -С. 6-9, 23
- Шендаков, А.И. Комплексный анализ результатов селекции молочного скота в Орловской области/А.И. Шендаков//Вестник Орёл ГАУ. -2010. -№ 2. -С. 16-22
- Шендаков, А.И. Результаты голштинизации молочного скота в Орловской области [Текст]/А.И. Шендаков//Аграрный вестник Урала. -2010. -№ 11 (77). -С. 70-72
- Шендаков, А.И. Влияние генетических и средовых факторов на интенсивность роста и молочную продуктивность чёрно-пёстрого голштинизированного скота [Текст]/А.И. Шендаков, Т.А. Шендакова//Вестник Орёл ГАУ. -2010. -№ 5. -С. 83-90
- Шендаков, А.И. Оценка эффективности отбора скота чёрно-пёстрой породы по молочной продуктивности [Текст]/А.И. Шендаков//Вестник ОрёлГАУ. -2010. -№ 6. -С. 93-100
- Шендаков, А.И. Результаты использования генетического потенциала молочного и комбинированного скота в Орловской области [Текст]/А.И. Шендаков, Т.А. Шендакова//Вестник Орёл ГАУ. -2011. -№ 1. -С. 14-21
- Шилер, Р. Селекция в животноводческой практике [Текст]./Р. Шилер, Я. Вахал, Я. Винш, перевод с чешского Г.Н. Мирошниченко; под ред. Д.В. Карликова. -М. Колос. -1981
- Эрнст, Л.К. Изучение влияния прилития крови голштинского скота на изменение генофонда крупного рогатого скота отечественных пород с использованием ДНК-микросателлитов [Текст]/Л.К. Эрнст, Н.А. Зиновьева, Е.Н. Коновалова и [др.]//Зоотехния. -2007. -№ 12. -С. 2-5
- Houle, D. Comparing Evolvability and Variability of Quantitative Traits [Text]/D. Houle//Genetics. -1992. -130:195-204
- Parland, S. Mc. Inbreeding effect on milk production, calving performance, fertility, and conformation in Irish Holstein-Friesians [Text]/S. Mc. Parland, J. F. Kearney, M. Rath, D. P. Berry//Dairy Science. -2007. -90:4411-4419
