Результаты предварительного фенетического анализа черепа синантропных домовых мышей (Mus musculus) предгорной популяции Центрального Кавказа
 предгорной популяции Центрального Кавказа")
Автор: Кучинова Екатерина Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 4-2 т.17, 2015 года.
Бесплатный доступ
Впервые для домовой мыши, обитающей в условиях предгорий Центрального Кавказа выделено 37 фенов неметрических признаков черепа, представляющих собой различного рода отверстия для прохождения кровеносных сосудов и нервов. Также проведено фенетическое сравнение синантропной домовой мыши с малой лесной мышью. Уровень фенетического разнообразия выше у домовой мыши по сравнению с малой лесной.
Домовая мышь, лесная мышь, неметрические признаки черепа, центральный кавказ
Короткий адрес: https://sciup.org/148203887
IDR: 148203887 | УДК: 575.2:599.323.4(470.64)
Phenetic preanalysis results of synanthropic house mice (Mus musculus) cranium from the piedmont population in the Central Caucasus
37 phenes of the cranium nonmetric parameters representing different openings for blood vessels and nerves are first distinguished for the house mouse occupying the foothills of the Central Caucasus. Phenetic comparison between the synanthropic house mouse and lesser common field mouse is also made. Phenetic diversity level is higher in the house mouse than in the lesser common field mouse.
Текст научной статьи Результаты предварительного фенетического анализа черепа синантропных домовых мышей (Mus musculus) предгорной популяции Центрального Кавказа
разных популяциях [12, с. 143]. Таким образом, возникновение мелких скелетных аберраций на черепе происходит в пределах генетически детерминированных границ и зависит, в пределах нормы реакции, от множества внешних и внутренних факторов, проявление которых обусловлено «эпигенетическим полиморфизмом» [11, 15]. Выше изложенное убедительно доказывает пригодность фенетического метода для широкого применения в популяционной биологии, в частности, для изучения популяционной структуры и дифференциации популяций различных видов млекопитающих. Перспективной территорией для изучения изменчивости широкоареальных видов является Кавказский регион. Наличие на Кавказе факторов поясности в горах и широтной зональности на смежных равнинах создают условия для того, чтобы оценить проявление внутривидовой изменчивости в градиенте влияния комплекса средовых факторов. Здесь домовая мышь населяет различные биотопы образуя высокую плотность населения на равнине, а с подъемом в горы, проявляет, высокую привязанность к жилищу человека [16].
Учитывая, что домовая мышь на Кавказе не изучалась методами фенетического анализа перед нами в основном стояли следующие задачи: 1) выделение фенов на черепе домовой мыши; 2) проведение корреляции с целью выбраковки признаков, связанных с полом, друг другом и т.д.; 3) составления каталога фенов для домовой мыши предгорий Центрального Кавказа.
МАТЕРИАЛ И МЕТОДЫ
Для проведения фенетического анализа была изучена серия черепов домовой мыши коллекционного фонда ИЭГТ из предгорий Централь- ного Кавказа. На данном этапе объем выборки составлил 40 экз. черепов взрослых животных. Относительный возраст зверьков определяли по стертости коренных зубов [17]. При проведении фенетического анализа (выделении, описании фенов, обработке результатов) использовали методы фенетики разработанные и предложенные А.Г. Васильевым [12, 18]. Поиск и учет фенов проводили с помощью микроскопа Carl Zeiss (Stemi 2000). Большая часть использованных нами в работе состояний неметрических признаков, и их кодовых сокращений взяты из литературных источников, а те, которые обнаружены впервые, уже были описаны ранее другими авторами для других видов [9-10, 12, 14, 18-20]. Изначально было выявлено порядка 80 билатеральных и медиальных признаков, включая единичные и редкие. Очень редкие фены исключались из дальнейшего анализа, как малоинформативные. Связь проявления фенов с полом и друг с другом оценивали на основе расчета непараметрических коэффициентов корреляции Спирмена. После этого часть признаков, проявивших значимую связь с указанными выше факторами исключили из дальнейшего анализа.
В процессе классификации черепов, согласно методике, предложенной А.Г. Васильевым с соавторами [19], отмечали наличие того или иного признака для каждой особи, а затем в каждой выборке для билатерально проявляющихся признаков подсчитывали число сторон на которых фен (устойчивое состояние порогового признака) проявился, и учитывали общее число изученных сторон.
В конечном итоге анализировали частоты 37 неметрических признаков, которым присвоены порядковые номера и коды на основе латинских наименований (см. рис. 1 и каталог фенов).
Каталог неметрических дискретных признаков (фенов) черепа домовой мыши предгорной выборки Центрального Кавказа
-
1 – дополнительное лобное отверстие впереди основного FFracan
-
2 – дополнительное лобное отверстие сзади основного FFracpo
-
3 – отсутствие лобного отверстия FFracan(-)
-
4 – одинарное переднее отверстие на чешуйчатой кости FTmacan
-
5 – центральное отверстие на чешуйчатой кости FTM
-
6 – заднее дополнительное височное отверстие FTmacpo
-
7 – наличие нижнего дополнительного височного отверстия FTmasif
-
8 – наличие одинарного окна на сосцевидной кости FeMs (+)
-
9 – отсутствие окна на сосцевидной кости FeMs (-)
-
10 – наличие одинарного подъязычного отверстия FHgsi
-
11 – наличие удвоенного подъязычного отверстия FHgdu
-
12 – наличие одинарного бокового предчелюстного отверстия FPmla
-
13 – верхнечелюстное отверстие на скуловом отростке FMxzm
-
14 – переднее отверстие на скуловом отростке верхней челюсти FMxzmor
-
15 – наличие одинарного отверстия в крыловидной ямке FRTI
-
16 – наличие одинарного дополнительного круглого отверстия FPD
-
17 – наличие отверстий на перегородке между овальным и круглым отверстиями FLTI
-
18 – наличие одинарного дополнительного
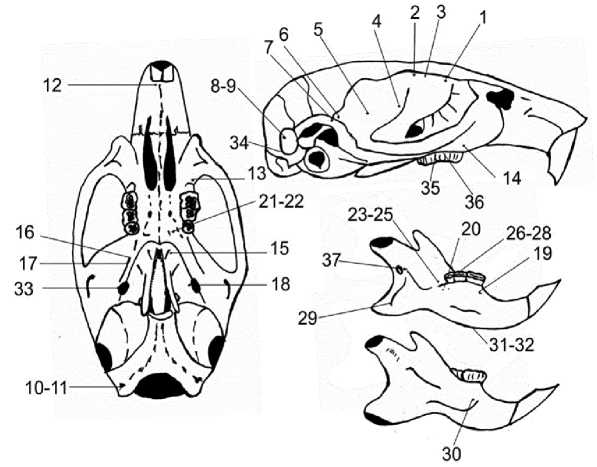
Рис. 1. Расположение фенов неметрических признаков на черепе домовой мыши
отверстия в области овального отверстия с вентральной стороны в крыловидной ямке Fasac
-
19 – наличие одинарного отверстия с лингвальной стороны альвеолы M1 FMtlg
-
20 – наличие двойного отверстия с лингвальной стороны альвеолы M3 FMblgpodu
-
21 – наличие одинарного дополнительного небное отверстие FPl
-
22 – наличие двойного дополнительного небное отверстие FPldu
-
23 – наличие одинарного отв. В области М3 на внутренней поверхности FPC (M3)
-
24 – наличие тройного отв. В области М3 на внутренней поверхности FPCtr (M3)
-
25 – наличие более 3 отверстий в области М3 на внутренней поверхности FPCmx (M3)
-
26 – наличие одинарного отверстия в области M2 на внутренней поверхности FPC (M2)
-
27 – наличие двойного отверстия в области M2 на внутренней поверхности FPCdu (M2)
-
28 – наличие более 3 отверстий в области M2 на внутренней поверхности FPCmx (M2)
-
29 – отсутствие отверстия в области сочленовно-угловой вырезки FMbacpo (-)
-
30 – наличие отверстия на массетерной площадке нижней челюсти выше linea oblique FMas
-
31 – наличие тройного отверстия на теле нижнечелюстной кости FOMtr
-
32 – наличие более 3 отверстий на теле нижнечелюстной кости FOMmx
-
33 – дополнительное отверстие в области овального с латеральной стороны FOvala
-
34 – множественное отверстие на затылочной кости в сочленовной ямке FFsOсmx
-
35 – наличие отверстия на боковой стороне альвеолы M2 верхней челюсти FPL (M2 )
-
36 – наличие отверстия на уровне M1 верхней челюсти FPL (M1 )
-
37 – наличие дополнительных отверстий в области венечного отверстия с внутренней стороны FMan
Фенетические дистанции (MMD) между выборками и их средние стандартные отклонения (MSD) рассчитывали по формулам, предложенным С. Хартманом [21]. Различия статистически значимы на уровне р < 0.05 при MMD > 2 MSD.
Для оценки фенетического разнообразия использовались общепринятые показатели Л.А. Животовского [22]. Внутрипопуляционное разнообразие оценивали с помощью показателя – μ, а концентрацию редких фенов с помощью – h.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Направленная ассиметрия (встречаемость на левой и правой сторонах черепа) выявлена по двум признакам (наличие переднелобного и заднего бокового предчелюстного отверстий). C полом оказались связаны 8 признаков (наличие одинарного отверстия в основании крыловидного отростка, двойное дополнительное круглое отверстие, наличие тройного отверстия с лингвальной стороны альвеолы М3, наличие одинарного медиального отверстия и окна на клиновидной кости, наличие одинарного отверстия и более трех отверстий на теле нижнечелюстной кости, отверстия по бокам клиновидной кости). Связь друг с другом проявили около 50% признаков.
При сравнении самцов и самок домовой мыши по частоте встречаемости признаков достоверные различия выявлены только по 1 признаку (наличие тройного отверстия на теле нижнечелюстной кости FOMtr) из 37. Причем, данный признак достоверно чаще встречается у самок, чем самцов. Уровень фенетических дистанций между полами, оцененный по всему комплексу признаков оказался статистически недостоверным MMD = - 0.00686±0.0115 (Хи-квадрат= 31,85; d.f.=37).
Учитывая, что домовая мышь в теплые периоды года выселяется из жилищ человека в природу, где она в предгорьях Центрального Кавказа симбиотопично встречается с широко распространенным и многочисленным видом – малой лесной мышью ( A. uralensis ), представляло интерес сравнить оба вида, происходящие с одного высотного уровня (предгорья) Центрального Кавказа.
С целью сопоставимости данных при межвидовом сравнении использовали только равное число неметрических признаков, встречающихся у обоих видов. Из 37 признаков выделенных у домовой мыши сходство между отмеченными видами установлено только по 13 признакам, по ним и проводили дальнейшее фенетическое сравнение (таб. 1).
Как видно из табл. 1 при внутривидовом сравнении у малой лесной мыши между зверьками разного пола не выявлено достоверных различий по частоте встречаемости какого-либо признака. У домовой мыши достоверные различия выявлены только по встречаемости одного признака - FOmx. В целом различия между полами недостоверны как у малой лесной MMD = - 0.0091±0.0213 (Хи-квадрат= 8,308; d.f.=13), так и домовой мыши MMD = 0.0061±0.0194 (Хи-квадрат= 14,61; d.f.=13).
При межвидовом сравнении достоверные различия между самцами выявлены по частоте встречаемости 5 признаков из 13 – Ftmacan, FeMs (-), FPmla, Fasac и FOmx. Данные признаки за исключением последнего достоверно чаще встречаются у самцов домовой мыши по сравнению с таковыми малой лесной мыши (табл. 1). При аналогичном сравнении самок достоверные различия также выявлены по встречаемости 5 неметрических признаков. Причем различия отмечаются практически по тем же признакам, что и у самцов за исключением признака FFracan. Как и у самцов, данные признаки достоверно чаще встречаются у самок домовой по сравнению с самками лесной мыши. Также обращает на себя
Таб. 1 . Частоты встречаемости фенов неметрических признаков черепа в популяциях домовой и малой лесной мышей (%) в предгорных экосистемах Центрального Кавказа
|
Код признака |
A. uralensis |
M. musculus |
Внутривидовое сравнение |
Межвидовое сравнение |
||||
|
33 |
33 |
33 |
33 |
|||||
|
n=40 |
n=34 |
n=40 |
n=40 |
Хи-квадрат |
Хи-квадрат |
|||
|
1 |
2 |
3 |
4 |
1-2 |
3-4 |
1-3 |
2-4 |
|
|
1. FFracan |
0 |
0 |
2.5 |
10 |
* |
|||
|
2. FFracan (-) |
0 |
0 |
2.5 |
5 |
||||
|
3. FTmacan |
7.5 |
12.1 |
27.5 |
15 |
* |
|||
|
4. FeMs (-) |
0 |
0 |
20 |
20 |
*** |
** |
||
|
5. FHgsi |
39.5 |
54.8 |
52.5 |
42.5 |
||||
|
6. FPmla |
0 |
0 |
40 |
37.5 |
*** |
*** |
||
|
7. FPD |
27.5 |
31.3 |
27.5 |
22.5 |
||||
|
8. Fasac |
18.4 |
32.3 |
67,5 |
70 |
*** |
** |
||
|
9. FMtlg |
37.5 |
26.5 |
60 |
55 |
||||
|
10. FFsOcmx |
62.5 |
58.8 |
30 |
30 |
||||
|
11. Mblgpodu |
25 |
23.5 |
25 |
10 |
||||
|
12. FPCdu |
15 |
20.6 |
27,5 |
34 |
||||
|
13. FOMmx |
85 |
67.6 |
57.5 |
82.5 |
* |
*** |
||
Примечание: n-число изученных сторон черепа. Уровень значимости межгрупповых различий по отдельным признакам: * – p < 0.5; ** – p < 0.01; *** – p < 0.001
Таб. 2. Характеристика фенетического разнообразия черепа домовых и лесных мышей Центрального Кавказа
При попарном сравнении двух видов по показателям внутрипопуляционного разнообразия – μ Л.А. Животовского следует, что уровни фене-тического разнообразия выше у домовой мыши по сравнению с лесной (табл. 2).
При сравнении зверьков по полу в пределах каждого вида по показателю μ различия выявлены у обоих видов, но половые различия сильнее выражены у малой лесной мыши, причем у самок уровень фенетического разнообразия выше, чем у самцов. У домовой мыши напротив фенети-чески более разнообразными являются самцы. При межвидовом сравнении по показателю μ различия выявлены во всех вариантах сравнения. По показателю h (концентрации редких фенов) различия оказались недостоверными во всех вариантах сравнения, хотя наибольшая доля редких фенов отмечается у зверьков обоих полов домовой мыши.
Таким образом, результаты фенетического анализа показали, что между двумя видами (А. uralensis и M. musculus) существуют значительные различия, как по частоте встречаемости отдельных признаков, так и по показателям разнообразия. При этом наибольшим фенетическим разнообразием характеризуются домовые мыши.
Список литературы Результаты предварительного фенетического анализа черепа синантропных домовых мышей (Mus musculus) предгорной популяции Центрального Кавказа
- Лапшов, В.А. Человек и популяционная экология синантропных грызунов/В.А. Лапшов, В.В. Кучерук//Синантропия грызунов. Характер поведения. Особенности обитания в постройках человека. Методы учетов. Пути ограничения численности/Ин-т эволюц. морфолог. и эколог. жив-х им. А.Н. Северцова РАН; . -Москва, 1994. -С. 4-14.
- Grunberg, H. Genetical studies on the skeleton of the house mouse/H. Grunberg//IV. Quasicontinuos variations. J. Genet. 1952. Vol. 51. -P. 95-114.
- Berry, R.J. Epigenetic polymorphism in wild population of Mus musculus/R.J. Berry//Genetic Reserch, Cambr. 1963. Vol. 4. -P. 193-220.
- Berry, R.J. The Evolution of island population of the house mouse/R.J. Berry//Evolution. 1964. -Vol. 18. -№3. -P. 468-483.
- Тимофеев-Ресовский, Н.В. Очерк учения о популяции/Н.В.Тимофеев-Ресовский, А.В. Яблоков, Н.В. Глотов. -М.: Наука, 1973. -278 с.
- Яблоков, А.В. Фенетика. Эволюция, популяция, признак/А.В. Яблоков. -М.: Наука, 1980. -135 с.
- Яблоков, А.В. Введение в фенетику популяций/А.В. Яблоков, Н.И. Ларина. -М.: Высш. Школа, 1985. -160 с.
- Васильев, А.Г. Популяционная структура обыкновенной слепушонки: многомерный морфометрический и фенетический аспекты сравнения поселений вида в Южном Зауралье. Морфологическая и хромосомная изменчивость мелких млекопитающих/А.Г. Васильев, Н.Г. Евдокимов, В.П. Позмогова//Екатеринбург: Наука. Уральское отделение. 1992. -С. 37-51.
- Песков, В.Н. Фенетика и феногеография водяной полевки (Arvicola terrestris L.)/В.Н. Песков, И.Г. Емельянов//Вест. зоологии. -2000. -№3. -С. 39-44.
- Песков, В.Н. Фенетический анализпопуляционной структуры водяной полевки (Arvicola terrestris L.) Центральной и Восточной Палеарктики/В.Н. Песков, И.Г. Емельянов//Вест. Зоологии. -2000. -№6. -С. 65-70.
- Васильев, А.Г. Эволюционно-экологический анализ устойчивости популяционной структуры вида (хроно-географический подход)/А.Г. Васильев, И.А. Васильева, В.Н. Большаков. -Екатеринбург, 2000. -132 с.
- Васильев, А.Г. Эпигенетические основы фенетики: на пути к популяционной мерономии/Алексей Геннадьевич Васильев. -Екатеринбург: Академкнига, 2005. -640 с.
- Васильев, А.Г. Гомологическая изменчивость морфологических структур и эпигенетическая дивергенция таксонов: Основы популяционной мерономии/А.Г. Васильев, И.А. Васильева. -М.: Товарищество научных изданий КМК, 2009. -511 с.
- Амшокова, А.Х. Изменчивость неметрических признаков черепа малой лесной мыши (Rodentia, Muridae) на Центральном Кавказе. Сообщение 1. Внутривидовая изменчивость в связи с градиентом высоты/А.Х. Амшокова, Ф.А. Темботова//Животный мир горных территорий. Позвоночные. Нальчик. -2009. -С.168 -180.
- Уоддингтон, К.Х. Основные биологические концепции. На пути к теоретической биологии/Уоддингтон Конрад Хэл. -М.: Мир, 1970. -С.108 -115.
- Темботов, А.К. Млекопитающие Кабардино-Балкарской АССР/Темботов Асланби Казиевич. -Нальчик, 1960. -С.92-93.
- Клевезаль, Г.А. Принципы и методы определения возраста млекопитающих/Клевезаль Галина Александровна. -М: Т-во научных изданий КМК, 2007. -283 с.
- Васильев, А.Г. Пакет прикладных программ PHEN 3.0. /А.Г. Васильев//1995. -URL:: http://ecoinf.uran.ru (дата обращения 12.08.2015).
- Васильев, А.Г. Сравнение мигрирующих и оседлых особей рыжей полевки по комплексу неметрических признаков/А.Г. Васильев, И.Г. Васильева, В.Н. Большаков//Экология. 1996. Т.27, -№5. -С. 371-377.
- Васильева, И.А. Феногенетический анализ популяций малой лесной мыши (Apodemus uralensis Pall.) в эоне влияния Восточно-Уральского радиоактивного следа/И.Г. Васильева, А.Г. Васильев, Н.М. Любашевский//Экология. -2003. -№6. -С.325 -332.
- Hartman, S. E. Geographic variation analysis of Dipodomys ordii using non -metric cranial trains/S. E. Hartman//J. Mammal. -1980. -V.61, №3. -Р.436 -448.
- Животовский, Л.А. Показатели популяционной изменчивости по полиморфным признакам. Фенетика популяций/Л.А. Животовский//М.: Наука. -1982. -С. 38-44.
