Ритм насиживания у самок белой трясогузки Molacilla alba в юго-восточном Приладожье

Автор: Кузнецова Е.С.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 16 т.6, 1997 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140148680
IDR: 140148680
Текст статьи Ритм насиживания у самок белой трясогузки Molacilla alba в юго-восточном Приладожье
Из 10 видов рода Motacilla белая трясогузка Motacilla alba имеет наиболее широкое распространение и встречается в Палеарктике повсюду, за исключением островов Тихого океана. В разных частях ареала у этого вида изучали питание и поведение при добывании корма (Прокофьева 1962; Davies 1977; Вержуцкий 1980; Резанов 1981 и др.), рост и развитие птенцов, линьку, летние перемещения, миграции (Persson 1977; Кукиш 1974а,б, 1981; Яковлева и др. 1987; Афанасьева, Рымкевич 1990 и др.), размещение, экологию и поведение на зимовках (Zahavi 1971; Fleming 1981 и др.). Биология размножения белой трясогузки в юго-восточном Приладожье подробно изучена А.И.Кукишем (1976), однако в его статье почти нет сведений о гнездовом поведении этого вида. Только в работах Леинонена (Lei-nonen 1973a,b), выполненных в центральной Финляндии, и статье японских авторов (Nakumura et al. 1984) приведены подробные сведения о поведении белой трясогузки в гнездовой период.
Целью нашей работы было описание поведения самок белой трясогузки в период насиживания на основе собранного нами материала и имеющихся в литературе данных.
Материал и методика
Работа выполнена на юго-восточном берегу Ладожского озера (Олонецкий район Карелии) на полевом стационаре "Маячино" Института биологии КНЦ РАН (60°44' с.ш., 32°49' в.д.). Исследование вели на участке береговой полосы протяженностью 5 км и шириной от 50 до 100 м. Открытая, лишенная островов водная поверхность Ладожского озера отделена от прибрежных сосняков-зеленомошников узкой полосой песчаных и песчано-каменистых пляжей. Растительный покров на пробной площади развит фрагментарно и представлен узкой полосой злаков и отдельными куртинами ив Salix spp. Прибойная полоса захламлена сплавной древесиной, разнообразным техногенным мусором, сухим тростником и водорослями. В зависимости от уровня воды в озере ширина пляжей варьирует год от года, а после продолжительных и сильных штормов изменяется даже конфигурация берегой полосы.
Белая трясогузка гнездится здесь на песчаных и песчано-каменистых пляжах. Гнезда строит на земле под укрытием прошлогодней травы, сухого тростника, бревен, нижних ветвей сосен и елей, а также в постройках на берегу.
Охотно заселяет искусственные гнездовые укрытия. Кроме того, отдельные пары поселяются внутри прибрежных лесных массивов: на свежих вырубках, при-ручейных лугах, вдоль лесных дорог, среди редин, в деревнях.
В работе мы использовали метод случайных многомоментных наблюдений (Бардин, Ильина 1986; Бардин и др. 1987). Наблюдения вели из специального укрытия, изготовленного на основе зонтика (Зимин 1983). В июне -июле 1992-1993 провели 243 ч наблюдений за насиживанием кладок разной величины. В данной статье использованы наблюдения в период от откладки последнего яйца до начала вылупления птенцов. Материалы о поведении птиц в периоды откладки яиц и вылупления птенцов предполагается опубликовать отдельно.
В ходе наблюдений мы отмечали: время начала и окончания дневной активности самки, продолжительность однократного непрерывного сеанса насиживания ( Топ) и продолжительность однократного периода отсутствия самки на гнезде (7^, вычисляли т.н. плотность насиживания — общую продолжительность насиживания в течение 1 ч дневной активности самки (в мин/ч или %).
Результаты и обсуждение
В исследуемом регионе период откладки яиц у белой трясогузки длится с апреля до июля. Первые полные кладки в Ленинградской обл. можно найти в конце апреля - начале мая (Мальчевский, Пукинский 1983), в юго-восточном Приладожье и Карелии — в первой декаде мая (Кукиш 1976; Зимин 1988). Период массового начала размножения на Северо-Западе России приходится на конец мая.
В полной кладке обычно бывает от 3 до 7 яиц (Cramp 1988). На Северо-Западе России чаще всего встречаются кладки с 5 и 6 яйцами (Мальчевский, Пукинский 1983; Зимин 1988). По нашим данным, в юго-восточном Приладожье средняя величина кладки составила 5.3±0.3 яйца (и = 16), что незначительно отличается от данных других авторов (табл. 1).
Непрерывное насиживание начинается с появления последнего яйца и, по нашим наблюдениям, продолжается от И до 13 сут, в среднем — 12.2±0.8 сут (и = 8). По материалам разных авторов, продолжительность насиживания варьирует в пределах 11-17 сут (Гладков 1954), 12-14 сут (Кукиш 1976), 11-16 сут (Cramp 1988).
Время начала дневной активности самки в период насиживания определяли по времени первого утреннего вылета из гнезда, а время окончания — по времени последнего возвращения. Первые отлучки регистрировались в 218-313 (время местное солнечное). В последний раз самка возвращалась в гнездо в 1945-2О08. Ночью перерывов в насиживании не отмечено. В период максимальной длительности светового дня (III декада июня) дневная активность самки становится немного дольше за счет более раннего ее начала. Однако на нашем материале это увеличение продолжительности дневной активности оказалось незначимым (/-критерий, Р> 0.05). По сравнению с ясными и теплыми днями, в холодные и дождливые дни время окончания днев-
Таблица 1. Величина кладки у белой трясогузки
В Приладожье самки затрачивают на инкубацию в дневное время 13.9-16.2 ч, что составляет 77-90% от времени дневной активности. По данным других авторов, самка может проводить на гнезде 85.3% (Leinonen 1973а) и 75% дневного времени (Nice 1943).
У большинства представителей рода Motacilla длительность Топ может достигать 1 ч и более (Moreau 1949; Smith 1950; Buxton 1954; Коханов 1986 и др.), хотя у более мелких воробьиных птиц эта величина обычно значительно меньше. Так, например, у Parus montanus она варьирует от 8 до 22 мин (Haftorn 1979). Средняя продолжительность Топ у белой трясогузки в Финляндии (31.9 мин) оказалась несколько больше, чем в целом у мелких воробьиных — 29.8 мин (Nice 1943). По мнению Лейнонена (Leinonen 1973а), это обусловлено большими затратами времени на переваривание более крупных и грубых кормов, потребляемых птицами этого вида.
Таблица 2. Средняя продолжительность (мин) однократных сеансов насиживания (Тол) и отсутствия самки на гнезде (TOff) в зависимости от величины кладки
|
Величина кладки |
Топ |
TOff |
||||
|
л |
X |
SE |
п |
X |
SE |
|
|
4 |
25 |
45.8 |
1.4 |
30 |
6.8 |
0.5 |
|
5 |
28 |
43.3 |
1.7 |
32 |
8.2 |
0.5 |
|
7 |
27 |
42.3 |
1.7 |
31 |
8.3 |
0.6 |
В Приладожье Топ изменялась в пределах от 20 до 80 мин, составляя в среднем 45.9±0.8 (рис. 1). Чаще всего (58% случаев) сеанс насиживания длился от 34 до 52 мин.
Время прогулок самки (То^ варьировала от 3 до 21 мин и в среднем составило 7.8±0.2 мин (рис. 1). В большинстве случаев (51%) самка покидала кладку лишь на 5-8 мин. Эти значения сходны с имеющимися в лите-
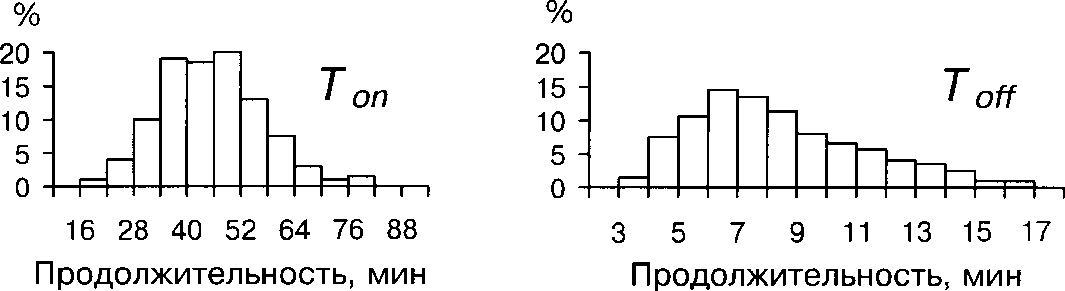
Рис. 1. Гистограммы распределения длительности периодов Топ и TOff у самок белой трясогузки в юго-восточном Приладожье ратуре данными (7.5 мин — Leinonen 1973а; 8.2 мин —Nakamura et al. 1984) и не значительно отличаются от средних значений для воробьиных в целом (8.5 мин — Nice 1943).
Продолжительность Топ несколько менее изменчива, чем продолжительность Toff (коэффициент вариации составил 27% и 39%, соответственно). Средние значения Топ и То^ отличаются у самок, имеющих кладки разной величины (табл. 2). Однако активность насиживания не зависит от величины кладки (табл. 3), и различия средних незначимы (2-критерий Тьюки, Р »0.05).
Вследствие большой продолжительности сеансов насиживания плотность насиживания также довольно велика: 34-60, в среднем 50.47±0.36 мин/ч. Эта величина является довольно устойчивой характеристикой ритма насиживания самки (СК= 10.2%). Как показал наш анализ (табл. 4), плотность насиживания слабо, но значимо увеличивается при увеличении Топ (г = 0.6, Р « 0.01) и при уменьшении 7^-самки (г = -0.5, Р«0.01).
Таблица 3. Дисперсионный анализ влияния величины кладки на продолжительность периодов Топ и То« ^ белой трясогузки
|
Вид дисперсии |
Сумма квадратов отклонений |
d.f. |
Средний квадрат |
F |
Р |
|
"Гon |
|||||
|
Факториальная |
153.79 |
2 |
76.90 |
1.11 |
> 0.05 |
|
Остаточная |
4966.36 |
72 |
68.98 |
||
|
Общая |
5120.15 |
74 |
|||
|
Факториальная |
41.70 |
2 |
20.84 |
2.39 |
> 0.05 |
|
Остаточная |
789.00 |
90 |
8.73 |
||
|
Общая |
820.70 |
92 |
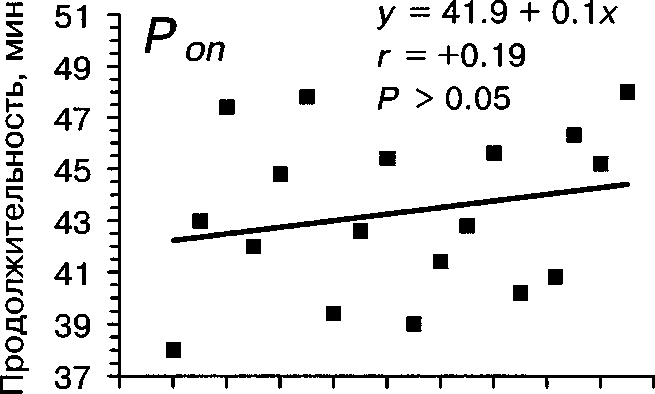
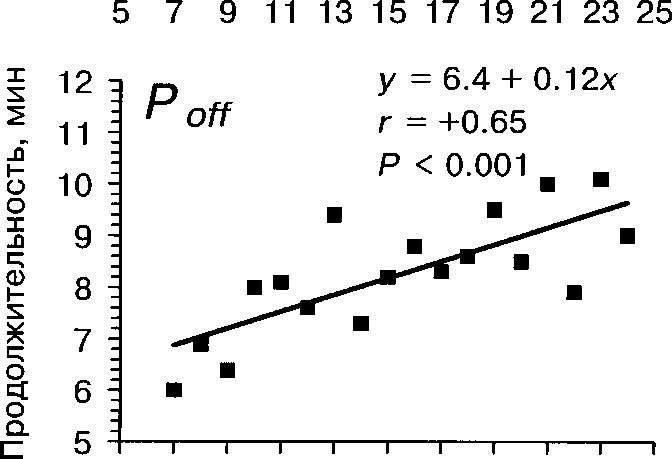
5 7 9 11 13 15 17 19 21 23 25
Температура воздуха, С
Рис. 2. Зависимость продолжительности периодов Топ и То« от температуры воздуха
Таблица 4. Значения коэффициентов корреляции между некоторыми переменными в период насиживания
|
Переменные |
Плотность насиживания, мин/ч |
Продолжительность периода Топ, МИН |
Продолжительность периода Toff, мин |
|
Плотность насиживания, мин/ч |
— |
+0.60* |
-0.50* |
|
Насиженность кладки, сут |
+0.90** |
+0.86** |
-0.98** |
|
Каледнарная дата, пятидневки |
— |
+0.76* |
-0.41* |
|
Температура воздуха, °C |
-0.83** |
+0.19 |
+0.65* |
|
Среднесуточная температура воздуха, °C |
-0.21 |
+0.37 |
-0.18 |
- Р< 0.01; ** - Р< 0.001.
ОО
•• орнитол. журн. 1997 Экспресс-выпуск № 16
Таблица 5. Изменение средней продолжительности периодов Топ и Toff и средней плотности насиживания у белой трясогузки в ходе периода насиживания
|
Сутки насиживания |
Продолжительность Топ, мин |
Продолжительность TOff, мин |
Плотность насиживания, мин/ч |
|||||||||
|
п |
lim |
X |
SE |
п |
lim |
X |
SE |
п |
lim |
X |
SE |
|
|
1 |
13 |
27-48 |
35.8 |
1.9 |
16 |
6-16 |
11.1 |
0.8 |
13 |
39-53 |
46.1 |
1.9 |
|
2 |
18 |
25-58 |
37.9 |
3.1 |
25 |
5-17 |
10.1 |
1.0 |
18 |
34-55 |
47.5 |
2.3 |
|
3 |
15 |
26-60 |
38.3 |
2.9 |
19 |
6-14 |
9.3 |
0.9 |
15 |
35-60 |
47.9 |
1.8 |
|
4 |
14 |
33-62 |
38.6 |
1.5 |
14 |
5-15 |
8.9 |
0.7 |
14 |
38-60 |
48.1 |
1.6 |
|
5 |
20 |
27-74 |
40.8 |
2.7 |
26 |
4-14 |
8.6 |
0.7 |
20 |
35-60 |
49.4 |
2.0 |
|
6 |
16 |
31-66 |
45.0 |
2.7 |
23 |
4-14 |
8.3 |
0.7 |
16 |
43-60 |
49.7 |
1.5 |
|
7 |
17 |
35-80 |
45.5 |
2.3 |
22 |
6-12 |
8.3 |
0.6 |
17 |
45-60 |
50.8 |
1.1 |
|
8 |
19 |
32-65 |
46.4 |
2.6 |
25 |
4-13 |
7.7 |
0.7 |
18 |
40-60 |
51.7 |
2.2 |
|
9 |
16 |
32-72 |
46.4 |
2.4 |
20 |
3-10 |
6.4 |
0.8 |
16 |
42-60 |
51.9 |
1.7 |
|
10 |
16 |
39-66 |
48.5 |
3.3 |
18 |
4-8 |
5.5 |
0.4 |
16 |
47-60 |
53.4 |
1.1 |
|
11 |
17 |
36-63 |
48.8 |
2.7 |
18 |
4-8 |
5.4 |
0.4 |
17 |
49-60 |
53.8 |
1.2 |
|
12 |
7 |
33-65 |
44.4 |
4.2 |
10 |
3-7 |
4.5 |
0.4 |
7 |
47-60 |
53.5 |
3.0 |
Активность насиживания более всего зависит от степени наси-женности кладки (табл. 4). При этом, если плотность насиживания и Топ возрастают от начала к концу периода насиживания, то То^ за это время почти втрое уменьшается (табл. 5). Мы объясняем это увеличением привязанности самки к кладке в ходе периода инкубации. Накануне вылупления продолжительность Топ и плотность насиживания несколько уменошаются (табл. 5). Видимо, это связано с возрастанием беспокойства самки в это время.
Длительность сеансов насиживания (Топ) и прогулок ^То^) самки — величины взаимосвязанные. При фиксированном значении сильно влияющего на эти параметры фактора насиженности кладки между ними выявлена положительная и значимая связь (коэффициент частной корреляции ту = 0.52, Р « 0.05). При этом Топ несущественно влияет на То^ как в разные дни с момента начала инкубации (табл. 6), так и за весь период в целом (г = -0.18, Р »0.05). Однако обратная зависимость более существенна: при увеличении Toff длительность последующего Топ имеет небольшую, но значимую тенденцию к увеличению (в целом за весь период г = 0.49, Р « 0.001). Наиболее сильной и значимой эта связь оказалась в течение первых 5 сут насиживания, а в последующие дни становится несущественной (табл. 6). Таким образом, длительность однократного сеанса насиживания определяется временем прогулки, в течение которого птица может собрать необходимое количество корма.
По нашим наблюдениям, активность насиживания самки зависит от температуры воздуха, времени суток и даты. При повышении температуры воздуха длительность Т^ увеличивается, а продолжительность Топ существенно не изменяется (табл. 4, рис. 2). Вследствие этого плотность насиживания также значительно уменьшается при повышении температуры воздуха (г = -0.83, Р « 0.001). Жарким летом 1992 самки насиживали кладку значимо дольше по сравнению с прохладным летом 1993, тогда как в длительности прогулок не обнаружено существенных различий (табл. 7).
Анализ показал отсутствие значимой зависимости активности насиживания от среднесуточных температур (табл. 4). Мы предполагаем, что в теплые дни самка быстрее собирает корм, что позволяет ей дольше не покидать гнездо, не увеличивая время прогулки. В холодные дни активность насекомых снижается, и самка не может найти достаточное количество корма за короткое время прогулки, чтобы затем долго насиживать кладку. Оставить кладку на длительное время в холодные дни самка не может, поскольку яйца быстро остынут и ей придется затратить значительную энергию для нагревания кладки после возвращения в гнездо.
Таблица 6. Изменение связи между ТОп и TOff на протяжении периода насиживания
|
Сутки насиживания " |
Корреляция между Топ и последующим TOff |
Корреляция между TOff и последующим Топ |
|
1 |
+0.06 |
-0.78* |
|
2 |
-0.09 |
-0.69* |
|
3 |
-0.08 |
-0.56* |
|
4 |
+0.21 |
-0.56* |
|
5 |
+0.25 |
-0.54* |
|
6 |
+0.30 |
+0.25 |
|
7 |
+0.05 |
+0.37 |
|
8 |
+0.07 |
-0.31 |
|
9 |
+0.32 |
+0.17 |
|
10 |
-0.06 |
+0.02 |
|
11 |
-0.09 |
-0.21 |
|
12 |
-0.05 |
-0.38 |
* - Р< 0.05.
Таблица 7. Средняя продолжительность (+SE)
|
периодов ТОП и TOff в 1992 и 1993 |
|||||
|
Период |
1992 |
1993 _ |
Значимость различий |
||
|
d.f. |
t |
Р |
|||
|
Топ |
52.2+0.6 |
45.9+0.4 |
196 |
2.9 |
< 0.05 |
|
TOff |
9.2+0.8 |
8.5+0.4 |
268 |
1.3 |
> 0.05 |
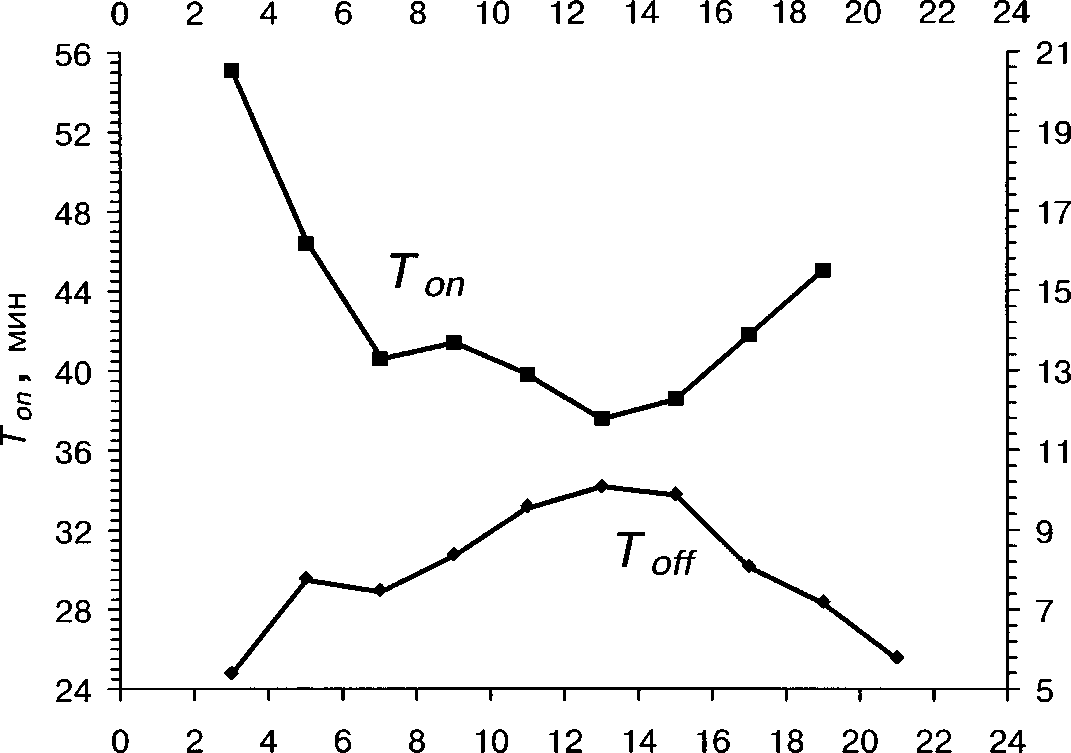
Для белой трясогузки характерна также суточная изменчивость активности насиживания (рис. 3). Как показал анализ, длительность Toff больше зависит от времени суток, чем продолжительность Топ (табл. 8). Наибольших значений длительность прогулок самки достигает в дневные часы, от 12 до 16 ч (в среднем 10.1+1.2 мин), а наименьших — в первые и последние часы дневной активности (в среднем 5.8+0.3 мин и 5.3+0.7 мин, соответственно; различия не значимы, /-критерий, Р »0.05).
Зависимость Топ от времени суток имеет обратную направленность (рис. 3). Этот параметр значительно уменьшается в середине дня (в интервале от 10 до 14 ч, в среднем 37.5+1.41 мин). Наибольших значений Топ достигает в утренние и вечерние часы (средние значения 55.1+2.9 и 43.6+3.7 мин, соответственно; различия не значимы, /-критерий Р »0.05).
1 Л Рус. орнитол. журн. 1997 Экспресс-выпуск № 16
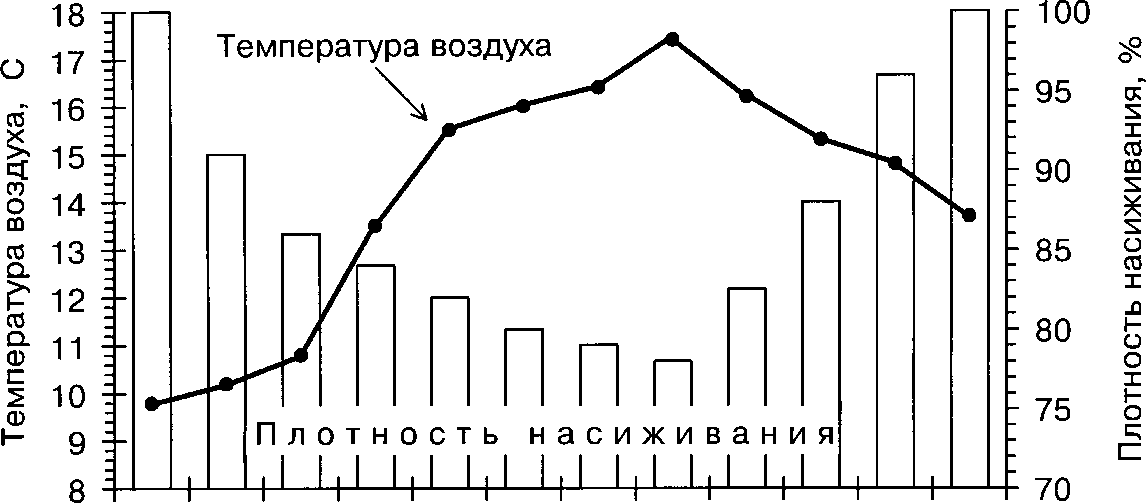
Время суток, ч
Рис. 3. Изменение средних значений плотности насиживания, длительности периодов Топ и То« и температуры воздуха в звисимости от времени суток
Таблица 8. Дисперсионный анализ влияния времени суток на активность нсиживающей самки белой трясогузки
|
Вид дисперсии |
Сумма квадратов отклонений |
d.f. |
Средний квадрат |
F |
Р |
|
Топ |
|||||
|
Факториальная |
2817.73 |
11 |
256.16 |
3.73 |
< 0.05 |
|
Остаточная |
7279.10 |
106 |
68.67 |
||
|
Общая |
10096.83 |
117 |
|||
|
Tgff |
|||||
|
Факториальная |
805.52 |
11 |
73.23 |
8.08 |
< 0.01 |
|
Остаточная |
1342.45 |
148 |
9.07 |
||
|
Общая |
2147.97 |
159 |
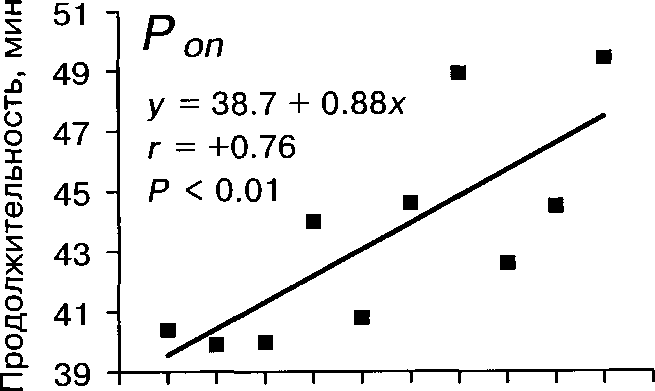
1 23456789 10 11
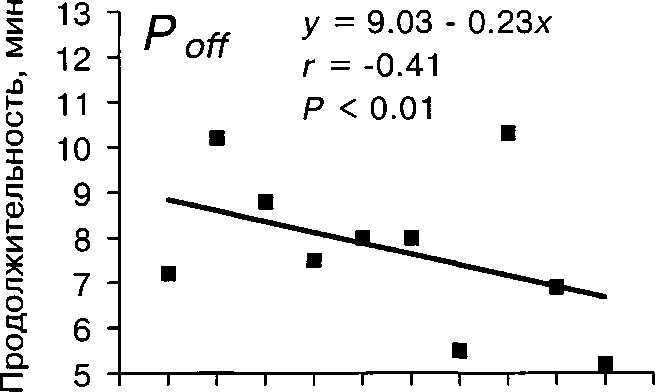
1 23456789 10 11
Пятидневки июня и июля
Рис. 4. Зависимость продолжительности периодов ТОп и То« от времени сезона
О 1 23456789 10 11 12
Сутки с начала инкубации
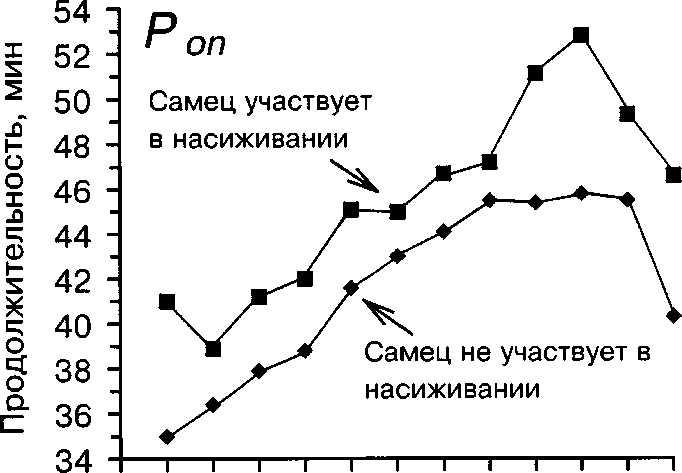
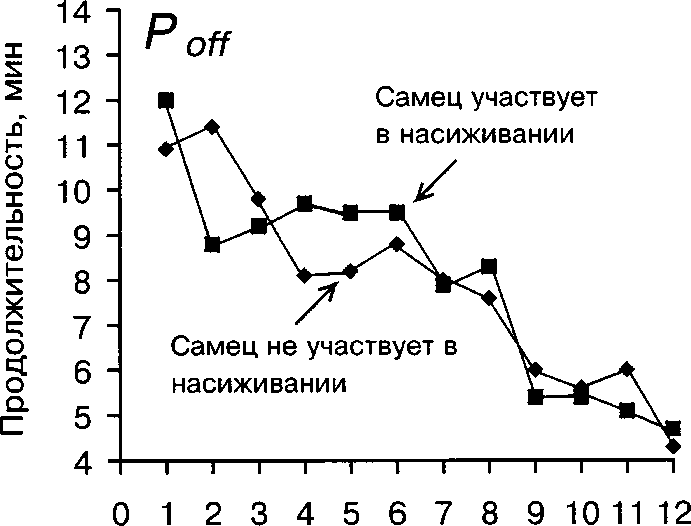
Рис. 5. Изменение продолжительности Топ и TOff у самки белой трясогузки в течение периода насиживания в зависимости от участия самца в насиживании
Как следствие, плотность насиживания также зависит от времени суток (корреляционное отношение ц = 0.53, Р « 0.001). Наименьших значений плотность насиживания достигает в дневные часы, с 12 до 16 ч ( в среднем 47.35±1.0 мин/ч), а наименьших — в утренние и вечерние часы (средние значения 54.7±0.5 мин/ч и 57.7±1.3 мин/ч, соответственно; различия не значимы, /-критерий, Р » 0.05). Мы считаем, что в изменения ритма насиживания в течение дня ведущую роль следует признать за температурным фактором — именно повышение температуры воздуха в середине дня позволяет самке покидать гнездо в этот период суток на больший срок. Кроме этого, в изменении ритма насиживания играет большую роль и внутренняя временная программа самки. .
Ритмы насиживания изменяются в течение сезона гнездования (рис. 4). Так, Топ значительно увеличивается от июня к июлю, а продолжительность То^ в этот период слабо, но значимо уменьшается (табл. 4). Возможно, такая тенденция способствует некоторому сокращению сроков инкубации яиц в конце сезона, отмеченному в Финляндии (Leinonen 1973а).
У белой трясогузки в насиживании участвуют и самка, и самец (Кукиш 1976; Мальчевский, Пукинский 1983; Nakamura et al. 1984 и др.). Однако, по нашим данным, основная роль в инкубации яиц все же принадлежит самке. Самец не всегда участвует в насиживании и не кормит насиживающую самку.
Как показали наши наблюдения, длительность сеансов насиживания самки значимо меньше, когда самец участвует в насиживании (G-критерий, Р « 0.001, рис. 5). В то же время на продолжительность прогулок самки участие самца не влияет (G-критерий, Р » 0.05, рис. 5). Вероятно, это связано с тем, что самец подменяет самку на гнезде обычно в холодную и дождливую погоду (68.5% случаев насиживания кладки самцом), когда при необходимости поддерживать более высокую плотность насиживания у самки появляется физиологическая потребность в сокращении продолжительности однократных сеансов инкубации.
Заключение
В юго-восточном Приладожье в условиях длительного светового дня для самки белой трясогузки характерна большая продолжительность дневной активности и кратковременность ночного покоя. Однако время начала и окончания дневной активности не зависит от продолжительности светового дня и погодных условий, а определяется скорее физиологическими потребностями самки.
Длительность однократного сеанса инкубации и плотность насиживания самки белой трясогузки довольно велики и превышают средние значения этих показателей для воробьиных в целом. В то же время продолжительность прогулок самки оказалась сходной со значениями, полученными для других регионов и видов воробьиных.
Ведущим фактором, определяющим ритмику насиживания, является степень насиженности кладки. Этот фактор объясняет 74% дисперсии Топ и 96% дисперсии То^. Римт насиживания также значительно изменяется в течения дня. Эту связь мы объясняем изменением температуры воздуха в течение суток и внутренней временной программой самки. Полученные результаты свидетельствуют о том, что длительность прогулки самки — наиболее изменчивая ха-14 Рус. орнитол. журн. 1997 Экспресс-выпуск №16
рактеристика. По-видимому, именно за счет изменения продолжительности То^ осуществляется регуляция ритмики насиживания самки в изменяющихся условиях. Например, изменения температуры воздуха в течение суток объясняют 42% дисперсии этой величины. Исследования показали довольно тесную связь времени прогулки и длительности последующего сеанса насиживания, особенно в первой половине периода инкубации. Вероятно, во второй половине этого периода привязанность самки к кладке возрастает настолько, что другие факторы уже не оказывают значимого влияния на Топ. Ведущими факторами, определяющими продолжительность Тот мы считаем степень привязанности самки к кладке, успешность кормежки во время прогулки и внутреннюю временную программу птицы.
Высокая плотность насиживания является следствием большой продолжительности сеансов насиживания. Это наиболее устойчивая характеристика ритма насиживания, определяемая температурой воздуха и степенью насиженности кладки. Последний фактор объясняет 81% дисперсии плотности насиживания. Другие факторы оказывают незначительное влияние.
Выражаю искреннюю признательность моим научным руководителям в аспирантуре В.Б.Зимину и А.В.Бардину.
