Роль артериоло-венулярных анастомозов в циркуляции нейтрофильных гранулоцитов

Автор: Марков Игорь Иванович, Маркова Валерия Игоревна, Малыхина Татьяна Викторовна, Антонян Янис Эдуардович, Софонов Матвей Витальевич, Рассолов Владимир Николаевич
Журнал: Морфологические ведомости @morpholetter
Рубрика: Оригинальные исследования
Статья в выпуске: 1 т.25, 2017 года.
Бесплатный доступ
Эмиграция нейтрофильных гранулоцитов (далее - НГ), основной функцией которых является фагоцитоз, происходит исключительно из просвета венул. Между тем, НГ «обвиняются» в том, что они могут блокировать микроциркуляцию в большом круге кровообращения и, тем самым способствовать развитию феномена no - reflow в головном мозге и миокарде. Вероятно, а priori считается, что НГ поступают в венозное русло транскапиллярно, хотя их морфологические параметры должны этому препятствовать. Возможна ли циркуляция НГ в венулы через артериоло-венулярные анастомозы (далее - АВА)? Целью работы явилось выяснение роли АВА в циркуляции нейтрофильных гранулоцитов. Были изучены препараты кишечника, скелетных мышц, миокарда, головного мозга, брюшины и селезенки кошек (n=7) и белых крыс (n=17). Методы - внутрисосудистая импрегнация по Ранвье, универсальный метод импрегнации различными солями серебра, рутинные гистологические методы окраски парафиновых срезов. У интактных животных нейтрофильные гранулоциты поступают в венулы только через АВА. В миокарде и головном мозге, где нет АВА, НГ за счет феномена гемосепарации поступают исключительно в эпикард, в первом случае, и в мягкие ткани лицевой области и твердую мозговую оболочку - во втором.
Нейтрофильные гранулоциты, артериоло-венулярные анастомозы, транскапиллярная миграция клеток
Короткий адрес: https://sciup.org/143177156
IDR: 143177156 | DOI: 10.20340/mv-mn.17(25).01.02
The role of arterio-venous anastomoses in the neutrophilic granulocytes circulation
Migration of neutrophilic granulocytes (NG), whose main function is phagocytosis, is primarily caused by the venule lumen. Nevertheless, NGs are often "blamed" for their ability to block microcirculation in the systemic circulation and thus to develop the no - reflow phenomenon in the cerebrum and myocardium. Apparently, NGs are a priori believed to enter the venous bed trans capillary, although their morphological features should prevent it. Are NGs able to circulate into venules via arterio-venous anastomoses (AVA)? The aim of the research is to elucidate the role of AVA in neutrophilic granulocyte circulation. The paper presents the study of the intestine, skeletal muscles, myocardium, cerebrum, peritoneum, and spleen in cats (n=7) and white rates (n=17). The research methods are intravascular impregnation by the Ranvier method, a universal method of impregnation with various silver salts, routine histological methods of staining paraffin sections. In intact animals, neutrophilic granulocytes enter the venules via AVAs only. In the myocardium and cerebrum, which have no AVAs, due to the blood separation phenomenon, NGs enter the epicardium exclusively in the former case and the facial soft tissues and pachymeninx in the latter.
Текст научной статьи Роль артериоло-венулярных анастомозов в циркуляции нейтрофильных гранулоцитов
Введение. Вопрос о механизмах блокады микрососудистого русла различных органов и, прежде всего, головного мозга и сердца, при различных экстремальных состояниях – вопрос первостепенной важности. От его решения зависит ответ: возможна ли ликвидация в клинических условиях феномена не восстановления кровотока (no-reflov)? В настоящее время многие авторы признают участие нейтрофильных гранулоцитов (НГ) не только в регуляции кровотока, но и в блокаде микроциркуляции [1-6]. По образному выражению [7], «лейкоциты подобно пастухам разгоняют эритроциты по своим капиллярам» [с. 173]. Феномен «no-reflov» был обнаружен в большинстве органов и тканей, испытавших острую ишемию, поскольку он имеет универсальный механизм возникновения и однотипный характер развития. Так, в работе [2] указано, что феномен «no-reflov» «состоит в массовой блокаде лейкоцитами венозных микрососудов» [с. 615] сосудистой оболочки головного мозга крыс, после перевязки у них обеих общих сонных артерий. Но каким образом НГ, имеющие жесткую конструкцию цитоскелета, вязкую цитоплазму и диаметр 12-15 мкм [8], проходят через капилляры в венулы? Ведь только из венул НГ способны эмигрировать в интерстициальное пространство [8, 9]. Этот вопрос в литературе не только не обсуждался, но даже и не ставился. Очевидно, что a priori признается транскапиллярная циркуляция НГ. Однако, вероятнее всего, НГ поступают в венозное русло органов через артериоло-венулярные анастомозы (АВА).
Цель работы – выяснить роль артериоло-венулярных анастомозов в циркуляции нейтрофильных гранулоцитов.
Материал и методы. Изучены препараты кишечника, сердца, головного мозга, брюшины, десны, языка, селезенки кошек (n=7) и белых крыс (n=17). В суправитальных условиях, под общей анестезией [Золетил (Virbac, Франция) в дозировке 125 мкг/кг] проводилась интрасосудистая импрегнация по Ранвье. Фрагменты органов импрегнировались по универсальному методу [10], а парафиновые срезы окрашивались гематоксилином и эозином, по Ван-Ризону, Романовскому - Гимзе. Все манипуляции с животными осуществились в соответствии с российскими этическими нормативами.
Результаты исследования и обсуждение. Типичные артериоло-венулярные анастомозы (АВА) с постоянным кровотоком обнаружены в эпикарде (рис. 1а), в твердой мозговой оболочке (рис. 1б), в подслизистой основе кишечника (рис. 1в). В миокарде, в мягкой мозговой оболочке, в коре головного мозга, в селезенке АВА обнаружить не удалось. Более того, в 10
просвете артериом и венул этих тканевых структур определялась высокая концентрация эритроцитов при полном отсутствии НГ (рис. 2). И тем не менее, в просвете «синусоидов» миокарда, отличающихся от типичных венул миокарда отсутствием в сосудистой стечке перицитов, небольшое количество НГ выявлялось у интактных кошек постоянно (рис. 3). У интактных же крыс, у которых отсутствуют межтрабекулярные пространства, НГ в просвете синусоидов отсутствовали. Эти данные свидетельствуют о том, что «синусоиды» могут сообщаться с камерами сердца [11] и НГ могут поступать в них во время диастолы. Циркуляции НГ в типичном микрососудистом русле миокарда препятствует процесс преорганной гемосепарации [12]. Он происходит последовательно, поэтапно по мере продвижения крови от сердца к месту назначения. Эффективность гемосепарации, разделение потоков крови, зависит от соотношения между углами отхождения магистральных артерий от аорты, перепадом диаметров материнских и дочерних ветвей и скорости кровотока в них [13]. Так, углы отхождения обеих венечных артерий от аорты равны ≈70°, а перепады диаметров – 1:15-17. Углы отхождения обеих общих сонных артерий близки к 90°, а соотношения диаметров – 1:2,5. В чем же биологическая целесообразность феномена преорганной гемосепарации? Очевидно, что в процессе гемосепарации в микрососудистое русло каждого органа и даже в микрососудистое русло каждой ткани органа поступает кровь различная по гематологическим показателям. Закономерно, что наиболее эффективная гемосепарация происходит в экстраорганном русле органов с высоким уровнем дифференцировки рабочих структур [14]. Так, после гемосепарации на уровне бифуркации левой общей сонной артерии и на уровне бифуркации плечеголовного ствола и бифуркации правой общей сонной артерии, в головной мозг поступает кровь с высоким гематокритом (0,584-0,712) и с выраженной лейкопенией [(2.5-3.0) х 109/л]. В наружные же сонные артерии поступает кровь с низким гематокритом (0,258-0,324) и с высоким лейкоцитозом [15]. Высокая эффективность гемосепарации в системе экстракраниальных артерий обусловлена так же значительной объемной скоростью кровотока во внутренних сонных артериях, в 2 раза превышающей этот показатель в наружных сонных артериях [16]. Такая разница в объемной скорости кровотока в артериях с практически одинаковым внутренним диаметром связана с несколькими факторами [17]. Во-первых, не смотря на ригидность черепа, несжимаемость мозга и ликвора в микрососудах мягкой мозговой оболочки возможно 20% увеличение объема крови. Во-вторых, в интракраниальных отделах внутренних сонных артерий происходит резкое снижение пульсового давления и значительное уменьшение сосудистого сопротивления [18]. В-третьих, функционированием «церебрального сердца», основным условием эффективности которого является синхронизация ритмических сокращений пиальных вен с артериальной пульсовой волной. В-четвертых, обе волны и артериальная волна расширения, и волна активного сокращения пиальных вен, возникающая синхронно и рефлекторно, управляются из единого центра – сердечного пейсмекера [18]. В-пятых, в кровоснабжении твердой мозговой оболочки участвует средняя оболочечная артерия. В связи с этим, именно в её микрососудистом русле существует большое число типичных АВА, через которые в венулы поступают НГ из наружной сонной артерии, низкий гематокрит и высокое осмотическое давление.
Напротив, в микрососудистом русле мягкой мозговой оболочки – высокая концентрация эритроцитов, но нет НГ и АВА. Таким образом, при отсутствии в твердой мозговой оболочке лимфатических микрососудов, резорбция ликвора из субарахноидального пространства осуществляется, очевидно, венозными микрососудами. Необычайно высокое содержание НГ наблюдается в микрососудах слизистой оболочки полости рта и, особенное, в микрососудах зубодесневых соединений. Высока и скорость миграции НГ в ротовую полость: около 30000 НГ в минуту и 4,3 х 10/у – за сутки [19].
Этому способствуют своеобразные сосудистые конструкции, подобные «шпилькам для волос», расположенные на уровне десневых сосочков и функционирующие как противоточные обменники, а также многочисленные АВА, обнаруженные на стыке прикрепленной десны и переходной складки и в области эпителиального прикрепления десны к зубу (рис. 4). При общем количестве циркулирующих в крови НГ идеального мужчины (масса – 70,0 кг, рост – 170 см) = 3,5 х10/10 за сутки в его полости рта погибает 1/800 их часть. Такая значительная потеря НГ происходит на площади слизистой оболочки полости рта, составляющей всего =137см² или менее 1/10000 площади слизистой оболочки желудочно-кишечного тракта [20]. 2/3 многочисленных физиологических функций НГ необходимо отметить их значительную роль в связывании и элиминации эндотоксина [21]. Ранее считалось [22], что весь эндотоксин, поступающий из кишечника, элиминируется клетками Купфера печени. В настоящее время установлено, что в норме до 6% объем крови, оттекающей от кишечника через портокавальные анастомозы, поступает, минуя печень, в общую циркуляцию. В связи с этим в периферической крови здоровых людей определяется до 13% активированных эндотоксином НГ [23]. При различных экстремальных ситуациях пропускная способность портокавальных анастомозов может увеличиваться, соответственно увеличивается и количество поступающего в общую циркуляцию эндотоксина. В ходе эволюции выработан ряд механизмов, обеспечивающих защиту организма от вредного действия эндотоксина [24].
К таким механизмам авторы относят различные виды взаимодействия эндотоксина с НГ:
-
1. Неспецифическое взаимодействие гидрофобных структур НГ с эндотоксином;
-
2. Связывание эндотоксина рецепторным белком СД18;
-
3. Формирование комплекса эндотоксина LВР- белком плазмы, а затем взаимодействие комплекса с поверхностным клеточным рецептором СД14;
-
4. Fc-зависимое связывание комплекса эндотоксин – антитело класса IgE с Fc–рецептором НГ [25].
Последний вид взаимодействия эндотоксина и НГ приводит к фагоцитозу и не активации эндотоксина, т.е. носит выраженный протективный характер [23]. Однако приходиться сомневаться в том, что только НГ защищают организм от эндотоксиновой агрессии. Полученные нами данные свидетельствуют о том, что поступление плазмы крови, содержащей не только эндотоксин, но и другие антигены, в микрососудистое русло органов, имеющих гистогематические барьеры, лимитируется преорганной гемосепарацией. Это, в первую очередь, касается головного мозга и миокарда. Более того, в микрососудистом русле и головного мозга, и миокарда отсутствуют АВА, через которые циркулируют НГ. С этих позиций морфофункциональные механизмы, осуществляющие преорганную гемосепарацию, должны рассматриваться как неотъемлемые части гистогематических барьеров, в том числе, и гематоэнцефалического. Отсутствие АВА в миокарде и в головном мозге исключает возможность блокады их микрососудистых русел НГ. Кроме того, на микрофотографиях [2], де-
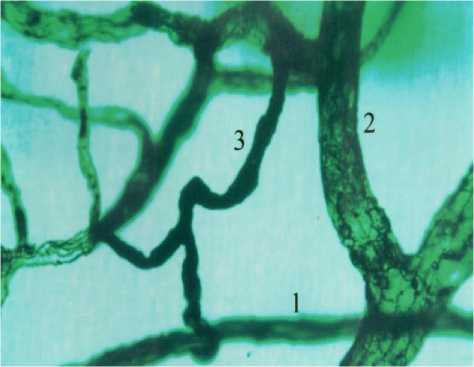
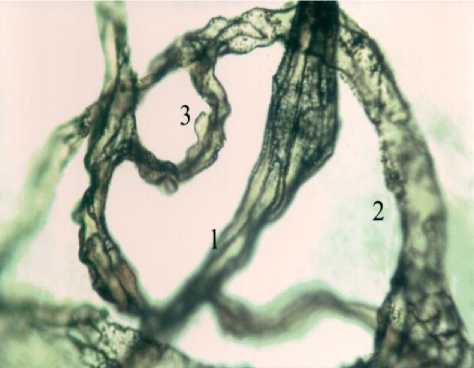
Рис. 1-Б
Рис. 1-A
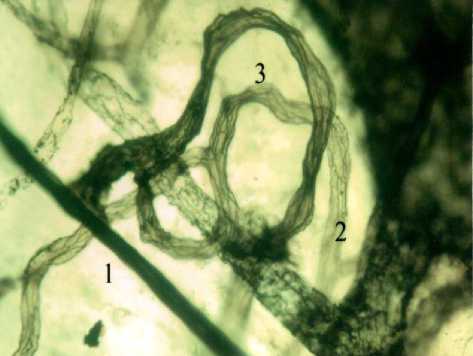
Рис. 1. Артериоло-венулярные анастомозы с постоянным кровотоком: в эпикарде (1-А), в подслизистой основе кишечника (1-Б), в большом сальнике (1-В) кошки. Обозначения: 1 - артериола, 2- венула, 3 - артериоловенулярный анастомоз. Внутрисосудистая импрегнация по Ранвье. Ув. 900.
Рис. 1-В
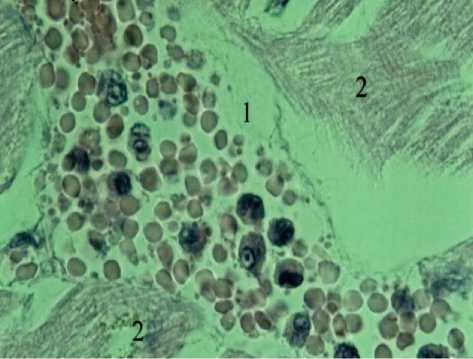
Рис. 2. Нейтрофильные гранулоциты в просвете синусоида (1) миокарда (2) кошки. Окраска гематоксилином и эозином. Ув. 900.
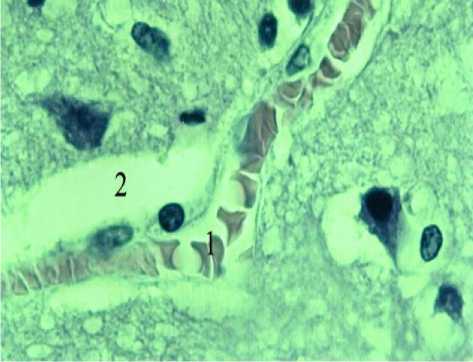
Рис. 3. Венула (1) коры головного мозга (3) кошки. 2 -перивазальное пространство. Окраска по Ван Гизону. Ув. 900.
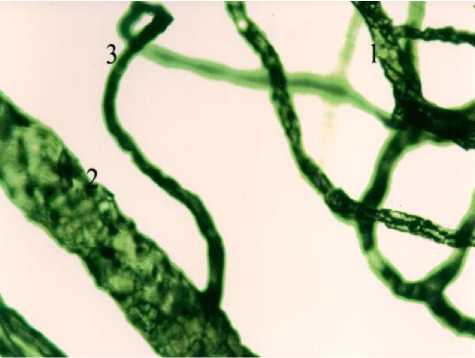
Рис. 4. Артериоло-венулярный анастомоз (3) в области эпителиального прикрепления десны к зубу кошки. 1) артериола, 2) венула. Внутрисосудистая импрегнация по Ранвье. Ув. 900.
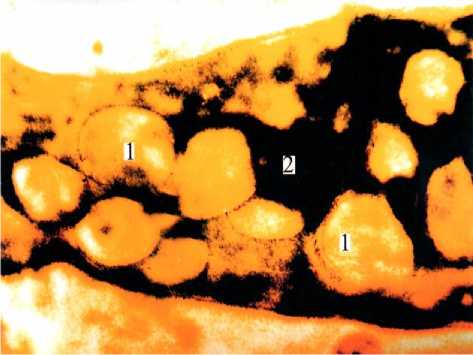
Рис. 5. Блокада лимфатического капилляра (2) набухшими и десквамированными эндотелиоцитами (1). Универсальный метод импрегнации. Ув. 900.
монстрирующих блокаду пиальных венул НГ, видны не НГ, а набухшие эндотелиоциты. Позже блокада кровеносных и лимфатических микрососудов набухшими и десквамированными эндотелиоцитами была проиллюстрирована (рис. 5) в работах [26, 27].
Список литературы Роль артериоло-венулярных анастомозов в циркуляции нейтрофильных гранулоцитов
- Иванов К.П. Изменение микроциркуляции при лейкоцитозе/К.П. Иванов, Ю.И. Левкович//физиол. журн. Им. И.М. Сеченова, 1992, № 6, с. 86 -90
- Иванов К.П. Лейкоциты как причина дисфункции микроциркуляции/К.П. Иванов, Н.Н. Мельникова//Бюлл. экспер. биол., 2006, № 6, с. 631 -634
- Маянский Д.Н. Роль нейтрофилов в ишемическом и реперфузионном повреждении миокарда/Д.Н. Маянский, С.Д. Маянская//Тер. арх., 2001, № 12, с.84 -88
- Редчиц Е.Г. Участие полиморфноядерных лейкоцитов в ишемической болезни сердца/Е.Г. Редчиц//Кардиология, 1989, № 12, с. 119 -124
- МиIIапе K. Neutrophyles and endothelial changes in reperfusion injury/Mullane//Trends Cardiovascul. Med. 1991, № 1, р. 282 -289
- Inoue T. Expresion of PMN leukocyte adnesion molecules and its clinical significance in patients treated with percutaneous transluminal coronary angioplasty/T. Inone, J. Sakal et al.//1. Amer. Coll. Cardiol., 1996, v. 28, p. 1127 -1133
- Гончаренко А.И. Компьютерное исследование минисердец/А.И. Гончаренко, Е.Е. Пастухова//Лесн. вестн., 2004, № 3, с. 159 -186
- Хэм А., Кормак Д. Гистология: Пер. с анг. М. Мир. 1983, т.2, с.254
- Воспаление, иммунитет и гиперчувствительность. Пер. с анг. Под ред. Я.Л. Рапопорта. -М. Медицина, 1975, с. 560
- Марков И.И. универсальный метод элективного выявления аргирофильных структур/И.И. Марков и др.//Морфологические ведомости, 2016, № 1, с. 116 -119.
- Grant R.T. Observation on the anatomy of the Thebesian vessels of the Heart/R.T. Grant, L. Vico//Heart, 1929, v.25, № 1, p. 103 -123.
- Адыширин-Заде Э.А. Структурные основы циркуляторного гомеостаза/Э.А. Адыширин-Заде//В кн: Морфологические аспекты органной гемоциркуляции. Куйбышев, 1988, с. 5 -8
- Марков И.И. Роль экстраорганных артерий и микрососудистого русла органов в формировании гистогематических барьеров/И.И. Марков, А.И. Марков и др.//Морфологические ведомости, 2002, № 1 -2, с. 75 -78.
- Мчедлишвили Р.И. Приоткрывая покровы неизвестного в физиологии и патологии микроциркуляции крови/Р.И. Мчедлишвили//Пантол. физиол., эксперт. терапия, 1991, № 3, с. 3 -7
- Цымбалов Э.Е. Структурная организация микрососудистого русла слизистой оболочки полости рта/Э.Е. Цымбалов//Автореф. дис.. канд. мед. наук. Саранск, 2008, с. 26
- Морошкин В.С. Состояние экстра-интра-краниального кровотока у больных ишемической болезнью сердца/В.С. Морошкин, О.М. Моисеева//Артериальная гипертензия, 2012, № 2, с. 153 -157
- Абасов Б.Х. Различия во внешнем строении шейного отдела общей сонной артерии и ее ветвей и возможности замещения дефектов перемещением наружной сонной артерии/Б.Х. Абасов//Автореф. дис.. канд. мед.наук, Баку, 1960, с. 21
- Кошев В.И., Петров Е.С. Гидродинамический флаттер и антифлатерная стабилизация в сердечно-сосудистой системе//Самара, 2007, 407 с.
- Быков В.Л. Тканевые и клеточные защитные механизмы слизистой оболочки полости рта/В.Л. Быков/Морфология, 1996, № 6, с. 14 -19
- Марченко Я.В. Структурная организация микрососудистого русла лицевой области/Я.В. Марченко//Автореф. дис.. канд.мед. наук. Саранск, 2009, с.25
- Пермяков Н.К. Эндотоксин и система полиморфноядерного лейкоцита/Н.К. Пермяков, М.Ю. Яковлев, В.Н. Галанкин//Арх. патол. 1989, № 5, с.3-11
- Маянский А.Н., Маянскин Д.Н. Очерки о нейтрофиле и макрофаге//Новосибирск, 1983, 217 с.
- Пермяков Н.К. Иммунная оценка резервов связывания эндотоксина полиморфноядерными лейкоцитами // Н.К. Пермяков, И.А. Аниховская и др. // Арх. патол. 1995, № 2, с.4 - 7
- Лиходед В.Г., Аниховская И.А., Аполлонин А.В., Козлова Н.Н., Кудрявцев А.Е., Хабриев Р.У., Ющук Н.Д., Яковлев М.Ю. Fc -зависимое связывание эндотоксинов граммотрицательных бактерий полиморфно ядерными лимфоцитами крови человека/В.Г. Лиходед, И.А. Аниховская и др.//ЖМЭиИ, 1996, № 2, с. 76 -79
- Яковлев М.Ю. Системная эндотоксинемия в физиологии и патологии человека//Автореф. дис.. докт. мед.наук М. 1993, 55 с.
- Кошев В.И. Диапедезные кровотечения -основная причина развития стойких нарушений микроциркуляции при различных патологических процессах/В.И. Кошев, Е.С. Петров, И.И. Марков//Морфологические ведомости, 2009, № 1 -2, с. 23 -26
- Кошев В.И. Эндотелиальные механизмы развития постишемического развития феномена не восстановления микрогемо -илимфоциркуляции/В.И. Кошев, Е.С. Петров, И.И. Марков//Морфологические ведомости, 2009, № 1-2, с. 127 -129
