Роль ежа обыкновенного (Erinaceus europaeus Linnaeus, 1758) в прокормлении иксодовых клеще
 в прокормлении иксодовых клеще")
Автор: Юрий Валерьевич Глазунов, Лариса Александровна Глазунова
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Ветеринария и зоотехния
Статья в выпуске: 10, 2021 года.
Бесплатный доступ
Иксодовые клещи представляют серьезную опасность для человека и животных, так как являются переносчиками и резерватами возбудителей многих заразных болезней. Для поддержания своей численности в природе иксодидам необходимы животные прокормители. Целью исследований явилось изучение уровня трофических связей ежа обыкновенного с иксодовыми клещами всех фаз развития, обитающих в Северном Зауралье. Исследовательскую работу проводили в период с мая по сентябрь 2014–2019 гг. в таежно-лесной и лесостепной зонах Северного Зауралья. Всего собрано 35 особей ежа обыкновенного (Erinaceus eurоpaeus Linnaeus). Установлено, что в таежно-лесной и лесостепной зоне Северного Зауралья еж обыкновенный является оптимальным прокормителем для всех фаз развития иксодовых клещей, обитающих на этой территории. Имаго, нимфы и личинки иксодовых клещей I. persulcatus и D. reticulatus могут одновременно и комфортно питаться на одном и том же прокормителе. Более половины всех собранных с ежей иксодид находились в стадии имаго (52,40 %), в меньшей степени обнаруживали нимф (29,08 %) и личинок (18,52 %). Доминирующим видом иксодид в обследуемых стациях являлся I. persulcatus, ИД которого составил 62,68±4,74 % в подзоне северной лесостепи и 88,16±1,58 % в подзоне подтайги. Обилие клещей всех фаз развития составило 50,19±1,66 особей в лесостепной и 42,22±1,72 % в таежно-лесной зоне. Среди фаз развития, паразитирующих на ежах во всех природно-климатических зонах, преобладала имагинальная (53,03 % в лесостепной и 50,26 % в таежно-лесной).
Лесостепная зона, таежно-лесная зона, иксодовые клещи, еж обыкновенный, имаго, нимфы, личинки.
Короткий адрес: https://sciup.org/140257843
IDR: 140257843 | УДК: 619:616.993:595.421: 599.365 | DOI: 10.36718/1819-4036-2021-10-114-120
The common hedgehog (Erinaceus europaeus Linnaeus, 1758) role in feeding ixodid ticks
Ixodid ticks pose a serious danger to humans and animals, as they are carriers and reserves of pathogens of many infectious diseases. To maintain their numbers in nature, ixodids need animal hosts. The aim of research was to study the level of trophic relationships between the common hedgehog and ixodid ticks of all developmental phases inhabiting the Northern Trans-Urals. The research work was carried out from May to September 2014 to 2019 in the taiga-forest and forest-steppe zones of the Northern Trans-Urals. A total of 35 specimens of the common hedgehog (Erinaceus europaeus Linnaeus) were collected. It has been established that in the taiga-forest and forest-steppe zones of the Northern Trans-Urals, the common hedgehog is the optimal host for all phases of development of ixodid ticks inhabiting this territory. The adults, nymphs, and larvae of the ixodid ticks I. persulcatus and D. reticulatus can simultaneously and comfortably feed on the same host. More than half of all ixodids collected from hedgehogs were in the adult stage (52.40 %); nymphs (29.08 %) and larvae (18.52 %) were found to a lesser extent. The dominant ixodid species in the studied stations was I. persulcatus, whose ID was 62.68±4.74% in the subzone of the northern forest-steppe and 88.16±1.58 % in the subzone of the subtaiga. The abundance of ticks at all stages of development was 50.19±1.66 individuals in the forest-steppe and 42.22±1.72 % in the taiga-forest zone. Among the developmental phases parasitizing on hedgehogs in all natural and climatic zones, the imaginal (53.03 % prevailed in the forest-steppe and 50.26 % in the taiga-forest).
Текст научной статьи Роль ежа обыкновенного (Erinaceus europaeus Linnaeus, 1758) в прокормлении иксодовых клеще
Введение . Иксодовые клещи (Ixodidae) являются относительно небольшой группой членистоногих, которые в большинстве своем в качестве прокормителей выбирают наземных позвоночных животных [1]. Представители этого семейства широко распространены во всем мире и зачастую играют роль векторов возбудителей инфекционных и инвазионных болезней [2].
В границы Северного Зауралья входят Курганская, Тюменская область и автономные округа – Ямало-Ненецкий и Ханты-Мансийский. В Тюменской области встречается пять природно-климатических зон из шести, характерных для Западной Сибири, что оказывает существенное влияние на климатические условия и отражается на функционировании биологических систем. В Тюменской области резко континентальный климат, но это не помешало организмам, ведущим паразитический образ жизни, адаптироваться к ним и даже расширять свои ареалы на север [3, 4]. Распространение иксодид тесно связано с характеристиками природных стаций, кроме климатических особенностей и рельефа местности, большое значение имеет фауна млекопитающих, которая выполняет роль их прокормителей [5– 11]. Возможных участников трофических и фори-ческих связей в жизненных циклах иксодовых клещей в Северном Зауралье достаточно. В большинстве своем это мелкие насекомоядные и грызуны для преимагинальных фаз и крупные позвоночные для имаго иксодид [12–14]. Учитывая треххозяинность иксодовых клещей, обитающих в регионе, и универсальность некоторых прокормителей, необходимо уточнить участие отдельных видов животных в жизненных циклах иксодид.
Цель исследования. Изучение уровня трофических связей ежа обыкновенного с иксодо-выми клещами всех фаз развития в Северном Зауралье.
Материалы и методы исследования. Экспериментальная часть работы выполнена в лаборатории акарологии ВНИИВЭА – филиале ТюмНЦ СО РАН по программе № 1210420000666 «Изучение и анализ эпизоотического состояния по болезням инвазионной этиологии сельскохо- зяйственных и непродуктивных животных, пчел и птиц, изменения видового состава и биоэкологи-ческих закономерностей цикла развития паразитов в условиях смещения границ их ареалов», а также на кафедре инфекционных и инвазионных болезней ФГБОУ ВО ГАУ Северного Зауралья. Сбор ежей проводили в таежно-лесной и лесостепной зонах Северного Зауралья в период с мая по сентябрь 2014–2019 гг. Потенциального прокормителя иксодовых клещей – ежа обыкновенного (Erinaceus eurоpaeus Linnaeus) отлавливали по мере встречаемости в лесных массивах [15]. Отлов проводили в основном в районах летних пастбищ, во время маршрутных обследований территорий. Всего за период исследования отловили 35 ежей, с которых сняли 1682 ик-содовых клещей различных фаз развития. В обработке и анализе материала использовали стандартные индексы: индекс обилия (ИО) и индекс доминирования (ИД) [16].
Наши наблюдения были сосредоточены в таежно-лесной зоне на территории Сорокинско-го и Викуловского районов, в лесостепной зоне в Голышмановском и Ишимском районах Тюменской области.
Для визуального обследования ежей обездвиживали с помощью эфира. Проводили осмотр брюшной поверхности тела, так как в этом месте иглы отсутствуют и заменены длинными и грубыми волосами, что является комфортным местом для присасывания клещей.
Результаты исследования и их обсуждение. В Северном Зауралье иксодовые клещи представлены шестью видами – это Ixodes persulcatus Schulze, 1930, Dermacentor reticulatus Fabricius 1794 (D. pictus, Hermann, 1804), D. marginatus Sulz, 1776, I. apronophorus P. Sch., 1924, I. (Exopalpiger) trianguliceps Bir., 1895 и I. (Ceratixodes) plumbeus, Kirsch., 1936. Наиболее широкое распространение и ветеринарное значение в регионе имеют три вида клещей: I. per-sulcatus, D. reticulatus и D. marginatus [17, 18].
Замечено, что у личинок и нимф иксодовых клещей, обитающих на территории Северного Зауралья, отсутствует филогенетическая специфичность. Определяющим фактором является биотический, который обусловливает участие в прокормлении ювенильных форм иксодид животных, приспособленных к конкретным экологическим условиям. Изменение условий существования для прокормителей может существенно влиять на популяционные показатели иксодо-вых клещей [19].
Из отловленных 35 ежей 26 обитали в подзоне северной лесостепи и 9 в подзоне подтайги.
Результаты осмотра ежей представлены в таблице.
Во всех случаях на теле ежей одновременно обнаруживали все фазы развития иксодид в процессе питания: личинок, нимф и имаго. Вероятно, что еж обыкновенный является универсальным прокормителем, так как его этологические особенности удовлетворяют требования всех фаз развития иксодид.
Заклещеванность ежа обыкновенного ( Erinaceus eurоpaeus Linnaeus) иксодовыми клещами в лесостепной и таежно-лесной зоне Северного Зауралья в период с 2014 по 2019 г.
|
Зона (подзона) |
Жизненная форма клеща |
Количество осмотренных животных |
Собрано, особей иксодид |
ИО, особей |
|
Лесостепная / северная лесостепь |
Личинка |
26 |
241 |
9,27±0,66 |
|
Нимфа |
372 |
14,31±0,80 |
||
|
Имаго |
692 |
26,12±1,12 |
||
|
Итого по подзоне |
1305 |
50,19±1,66 |
||
|
Таежно-лесная / подтаежная |
Личинка |
9 |
71 |
7,89±0,65 |
|
Лимфа |
118 |
13,11±0,86 |
||
|
Лмаго |
191 |
21,22±1,00 |
||
|
Итого по подзоне |
380 |
42,22±1,72 |
||
|
ИТОГО |
1685 |
48,14±1,68 |
||
При осмотре 26 млекопитающих, отловленных в подзоне северной лесостепи, было снято 1305 особей иксодовых клещей всех фаз развития. Обилие иксодид на одном прокормителе составило 50,19±1,66 особей. Более половины (53,03 %) всех питающихся иксодовых клещей составили имаго – 692 особи, 18,47 % – личинки и 28,50 % – нимфы. В таежно-лесной зоне осмотрено 9 ежей, с которых снято 380 иксодовых клещей всех фаз развития. Обилие иксодовых клещей всех фаз развития составило 42,22±1,72 %. Обилие иксодовых клещей всех фаз развития – 42,22±1,72 %. Половину (191 особь) всех собранных особей составили имаго, наименее представленной фазой развития оказалась личиночная (71 особь).
При видовой идентификации иксодовых клещей установлено, что в зависимости от природно-климатической зоны индекс доминирования иксодид варьировал (рис. 1 и 2).
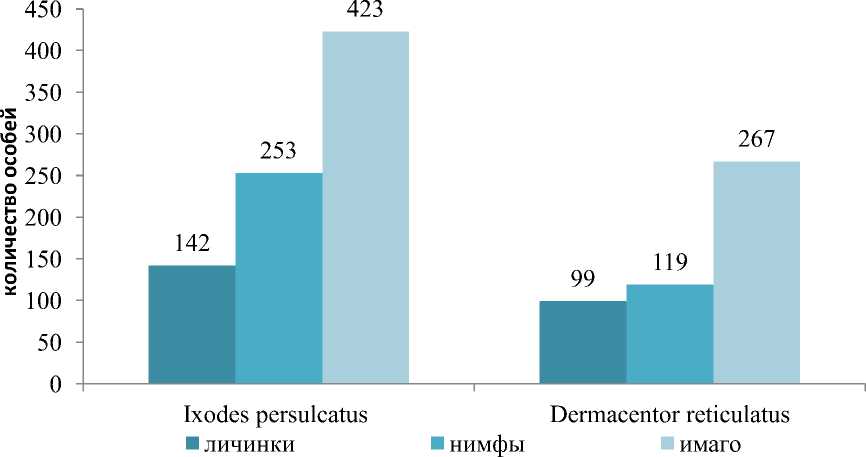
Рис. 1. Заклещеванность ежа обыкновенного в лесостепной зоне (подзоне северной лесостепи) в период с 2014 по 2019 г.
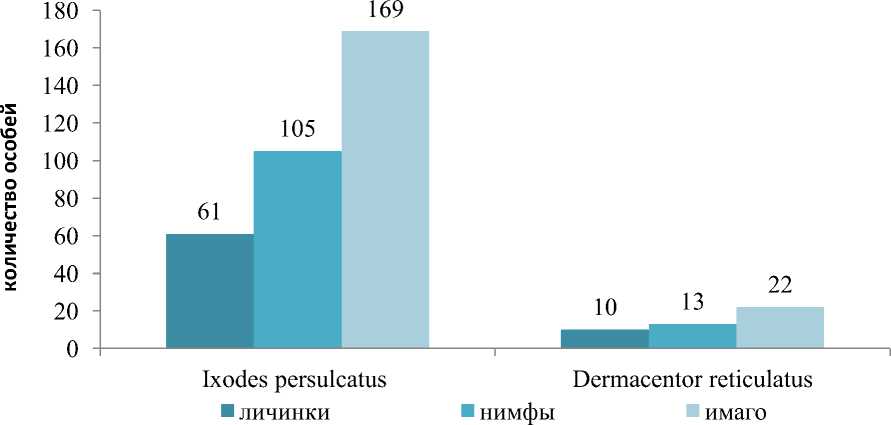
Рис. 2. Заклещеванность ежа обыкновенного в лесотепной зоне (подзоне северной лесостепи) в период с 2014 по 2019 г.
Как видно из рисунка 1, в подзоне северной лесостепи, несмотря на универсальность, наиболее часто фиксировали питание имаго клещей. Так, более половины всех собранных иксодид с ежей находились в имагинальной стадии развития (52,40 %), в меньшей степени обнаруживали нимф (29,08 %) и личинок (18,52 %). Кроме того, замечено, что доминирующим видом иксодид в обследуемых стациях являлся I. persulcatus, ИД которого составил 62,68±4,74 % в подзоне северной лесостепи и 88,16±1,58 % в подзоне подтайги (см. рис. 2). Выявлено, что в подзоне северной лесостепи обилие иксодид во всех стадиях развития по числовым показателям превосходило таковые в подзоне подтайги.
При видовой идентификации клещей, паразитирующих на ежах в подзоне северной лесостепи, определяли иксодид трех видов. Редко встречающимся видом оказался D. marginatus , представителей которого собрали всего две особи в фазе имаго. В наибольшей степени встречались клещи вида I. persulcatus, которые преобладали по всем генерациям. Наиболее представленной оказалась нимфальная стадия, особи которой составили 68,01 % всех нимф, собранных в подзоне северной лесостепи. ИД D. reticulatus составил 37,17±4,70 %, основной фазой развития явились личинки, которые составили 41,08 % всей генерации клещей этого вида.
В таежно-лесной зоне половину (191 особь) всех собранных особей составили имаго, наименее представленной фазой развития оказалась личиночная (71 особь). В указанной подзоне встречались клещи двух видов I. persulcatus и D. reticulatus. Таежный клещ преобладал во всех фазах развития, но наименьшее количество особей снимали в личиночной фазе. Особи D. reticulatus составили лишь 11,84±2,05 %, учитывая низкую распространенность клещей этого вида, достоверных отличий в количественном соотношении фаз развития не установлено.
Заключение. Установлено, что в таежнолесной и лесостепной зоне Северного Зауралья еж обыкновенный является универсальным прокормителем для всех фаз развития иксодовых клещей, обитающих на этой территории. Имаго, нимфы и личинки иксодовых клещей I. persu-lcatus и D. reticulatus могут одновременно и комфортно питаться на одном и том же прокормите- ле. Более половины всех собранных с ежей ик-содид находились в имагинальной стадии развития (52,40 %), в меньшей степени обнаруживали нимф (29,08 %) и личинок (18,52 %). Доминирующим видом иксодид в обследуемых стациях являлся I. persulcatus, ИД которого составил 62,68±4,74 % в подзоне северной лесостепи и 88,16±1,58 % в подзоне подтайги. Обилие клещей всех фаз развития составило 50,19±1,66 особей в лесостепной и 42,22±1,72 % в таежнолесной зоне.
Список литературы Роль ежа обыкновенного (Erinaceus europaeus Linnaeus, 1758) в прокормлении иксодовых клеще
- Балашов Ю.С. Иксодовые клещи – парази-ты и переносчики инфекций. СПб.: Наука, 1998. 287 с.
- Альмякова Е.Г., Донкова Н.В. Динамика заболеваемости собак бабезиозом в городе Красноярске // Вестник КрасГАУ. 2020. № 6 (159). С. 194–198.
- Григор Г.Г., Земцов А.А. Природное рай-онирование Западной Сибири // Природное и сельскохозяйственное районирование СССР. М.: Географгиз, 1961. С. 82–89 (Вопросы географии. Сб. 55).
- Каретин Л.Н. Почвы Тюменской области. – Новосибирск: Наука, 1990. 281 с.
- Глазунов Ю.В., Глазунова Л.А. Биотические связи имаго пастбищных клещей в Север-ном Зауралье // Современные проблемы науки и образования. 2015. № 4. С. 536.
- Глазунов Ю.В. Особенности биологии и методы ограничения численности иксодо-вых клещей в Северном Зауралье // Мир Инноваций. 2017. № 2. С. 20–24.
- Глазунов Ю.В., Глазунова Л.А. Взаимоотно-шения личинок и нимф иксодовых клещей (Ixodidae, Parasitiformes) с мелкими млекопи-тающими в Зауралье // Фундаментальные исследования. 2015. № 2-25. С. 5588–5593.
- Глазунов Ю.В., Домацкий В.Н., Глазуно-ва Л.А. Биофенология прокормителей личи-нок и нимф иксодовых клещей в лесостепной и таежно-лесной зоне Зауралья // Вестник АПК Ставрополья. 2014. № 3(15). С. 95–99.
- Глазунов Ю.В., Глазунова Л.А. Роль диких млекопитающих в прокормлении преимаги-нальных фаз иксодовых клещей в Тюмен-ской области // Фундаментальные исследо-вания. 2013. № 4-2. С. 371–374.
- Панов В.В. Мелкие млекопитающие лесо-парковой зоны ННЦ, численность и роль в прокормлении преимагинальных стадий ик-содовых клещей // Вестник ИрГСХА. 2017. № 83. С. 125–131.
- Собанский Г.Г. Трансформация фауны мле-копитающих Горного Алтая продолжается // Бюл. Московского общества испытателей природы. Отдел биологический. 2009. Т. 114, № 6. С. 3–8.
- Балашов Ю.С. Специфичность паразито-хозяинных связей членистоногих с назем-ными позвоночными // Паразитология. 2001. № 5(35). С. 473–489.
- Глазунов Ю.В. Некоторые аспекты феноло-гии иксодовых клещей на юге Тюменской области // Современные проблемы науки и образования. 2013. № 6. С. 746.
- Глазунов Ю.В. Особенности биологии и методы ограничения численности иксодо-вых клещей в Северном Зауралье // Мир Инноваций. 2017. № 2. С. 20–24.
- Соколов В.Е. Систематика млекопитающих: учеб. пособие для вузов. М.: Высш. шк., 1973. С. 432.
- Беклемишев В.Н. Термины и понятия, необ-ходимые при количественном изучении по-пуляции эктопаразитов и нидиколов // Зоол. журнал. 1961. Т. 40, вып. 2. С. 149–158.
- Столбов Н.М. Обнаружение клещей Ixоdes plumbeum Leach в Тюменской области // Сб. науч. работ Тюменского НИИ краевой ин-фекционной патологии. Тюмень, 1965. С. 163–164.
- Глазунов Ю.В., Глазунова Л.А. Иксодовые клещи (биология, экология, методы ограни-чения численности). Тюмень, 2011.
- Романенко В.Н. Эколого-этологические ас-пекты иксодовых клещей (Parasitiformes, Ixodidae) различных ландшафтов: автореф. дис. ... д-ра биол. наук. Томск, 2007. 44 с.
