Роль ГАМК А-рецепторов комплекса Бетцингера в регуляции дыхания у крыс

Автор: Маньшина Надежда Геннадьевна, Ведясова Ольга Александровна
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Нормальная и патологическая физиология
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
В острых опытах на наркотизированных половозрелых крысах изучены изменения паттерна внешнего дыхания и биоэлектрической активности инспираторных мышц при микроинъекциях растворов (10 -6 М) мусцимола и бикукуллина в комплекс Бетцингера (КБ). Установлено, что инъекции мусцимола в КБ укорачивают вдох и выдох, повышая частоту дыхания, при этом дыхательный объем и легочная вентиляция увеличиваются, что совпадает с изменениями частотно-амплитудных параметров электромиограмм инспираторных мышц. При введении бикукуллина в КБ частота дыхания снижается, а объемные параметры спирограмм закономерно не меняются. Наблюдаемые реакции свидетельствуют о различной роли ГАМК А-рецепторов области КБ в механизмах регуляции ритма и объемных параметров дыхания.
Комплекс бетцингера, паттерн дыхания, гамк а-рецепторы, gaba а-receptors, мусцимол, бикукуллин
Короткий адрес: https://sciup.org/14112956
IDR: 14112956 | УДК: 612.282:612.285
Role of the Betzinger complex GABA A-receptors in respiratory control in rats
In adult anaesthetized rats the reactions of breathing and inspiratory muscles activity to microinjections of muscimol (10 -6 M) andbicuculline (10 -6 M) into the Betzinger complex (BC) were investigated. It was shown, that muscimol microinjections into the BC shortened inspiratory and expiratory time and increased respiratory frequency, under this conditions the tidal volume and lung ventilation were increased. These responses corresponded to responses of temporal and amplitude parameters of electromiogramms of inspiratory muscles activity. Аdministrations of bicuculline into the BC decreased respiratory frequency, while volumetric parameters of breathing was not changed essentially. It is suggested that the reactions observed demonstrate the various contribution of GABA A-receptors within BC, in control of the temporal and volumetric parameters of breathing pattern.
Текст научной статьи Роль ГАМК А-рецепторов комплекса Бетцингера в регуляции дыхания у крыс
Введение. Формирование дыхательного ритма и его регуляция обеспечиваются сложными межнейронными взаимодействиями с участием многих возбуждающих, а также тормозных нейромедиаторов [10, 16, 17, 19]. Важную роль как в формировании нормоп-ноэ, так и в возникновении патологических паттернов дыхания играет гамма-аминомасляная кислота (ГАМК), являющаяся тормозным нейропередатчиком, широко распространенным в центральной респираторной нейросети [1, 6, 24]. Свое действие в различных отделах дыхательного центра ГАМК реализует через две основные группы мембранных рецепторов: ионотропные рецепторы класса ГАМКА и метаботропные рецепторы класса ГАМКВ [20, 22, 23]. В настоящее время актуальными аспектами нейрохимии дыхания являются изучение значения ГАМКергической медиации и анализ роли разных классов ГАМК-рецепторов, представленных на уровне функционально специфических отделов дыхательного центра, в меха- низмах модуляции центральной инспираторной и экспираторной активности, регуляции паттерна дыхания [2, 4, 12]. В этом плане особый интерес представляет комплекс Бет-цингера (КБ), образованный популяциями экспираторных и инспираторных нейронов. Среди нейронов КБ в функциональном отношении наиболее значимы экспираторные нейроны типа Е2 [13, 15, 25], которые участвуют в процессах ГАМКергического торможения в пределах всей респираторной нейросети и тем самым вносят вклад в механизмы формирования дыхательной ритмики [5, 26]. Отдельные физиологи на основании результатов введения в КБ агонистов и антагонистов ГАМКА-рецепторов полагают, что этот класс рецепторов непосредственно участвует в респираторном ритмогенезе [9, 20, 21]. Авторы других экспериментальных исследований считают, что опосредованное ГАМКА-рецепторами торможение в пределах КБ у взрослых млекопитающих не имеет существенного значения для генерации нормального дыхательного ритма [22]. Таким образом, на сегодняшний день многие стороны вопроса о роли ГАМКА-рецепторов в регуляции ритма и паттерна дыхания остаются спорными и требуют дальнейшего экспериментального изучения.
Цель исследования. Анализ респираторных реакций при блокаде и стимуляции ГАМК А -рецепторов области комплекса Бет-цингера в условиях in vivo у половозрелых крыс.
Материалы и методы. Эксперименты поставлены на 20 беспородных крысах массой 250–300 г, наркотизированных внутрибрюшинным введением уретана (1,5 г/кг массы), при строгом соблюдении биоэтических правил. Операционная подготовка начиналась с трахеотомии и введения в трахею пластиковой трубки, после чего животных переворачивали спиной вверх, препарировали шейные мышцы и осуществляли доступ к дорсальной поверхности продолговатого мозга через атланто-окципитальное отверстие. Затем на правом боку животного делали разрез кожи от II–III межреберий до нижнего края грудной клетки с целью доступа к дыхательным мышцам. Прооперированных крыс фиксировали в стереотаксической установке СЭЖ-3 и закрепляли голову в положении вентрального сгибания. Тепловое состояние животных поддерживали с помощью электрогрелки.
Блокаду и стимуляцию ГАМК А -рецепто-ров КБ осуществляли путем локального воздействия растворов мусцимола и бикукуллина, которые инъецировали через стеклянную микроканюлю (диаметр кончика 20–25 мкм), укрепленную на игле микрошприца МШ-1 (цена деления 0,02 мкл). Растворы веществ в концентрации 10-6 М готовили ex tempore разведением в искусственной спинномозговой жидкости и вводили в КБ в объеме 0,2 мкл по стереотаксическим координатам [11]. Кончик микроканюли удерживали в мозге в течение всего времени регистрации респираторных эффектов во избежание распространения раствора вверх по треку.
Реакции внешнего дыхания оценивали по изменениям спирограммы, которую регистрировали через трахеотомическую трубку при помощи электронного спирографа с ма- нометрическим датчиком. Выходные сигналы от спирографа подавались на аналоговоцифровой преобразователь, а затем на компьютер, где записывались в виде пневмотахограмм с последующим преобразованием их в спирограммы с помощью программы PowerGraph 3.2 Professional (ООО «Интероптика-С»). Одновременно регистрировали суммарную и интегрированную биоэлектрическую активность диафрагмы и наружных межреберных мышц (НММ), которую отводили с помощью биполярных стальных игольчатых электродов (межэлектродное расстояние 3 мм), подсоединенных к миографическому усилителю. Запись спирограммы и электромиограммы (ЭМГ) осуществляли непрерывно, в т.ч. до начала воздействия фармакологических веществ и в течение 60 мин после их микроинъекции в КБ.
На спирограммах анализировали длительность вдоха и выдоха (с), дыхательный объем (ДО, мл). Расчетным путем определяли частоту дыхания (ЧД, мин-1) и минутный объем дыхания (МОД, мл/мин). На суммарных и интегрированных ЭМГ диафрагмы и НММ определяли длительность залпов инспираторной активности (ДЗ, мс), длительность межзалповых интервалов (МЗИ, мс) и амплитуду залповых разрядов (отн. ед.). Статистическую обработку результатов проводили в программе SigmaStat 3.5 при помощи однофакторного парного t-теста, теста ANOVA, тестов Dunnett’s и Tukey; достоверными считали различия при p<0,05.
Результаты и обсуждение. Характерной реакцией на микроинъекции в КБ раствора ГАМК А -агониста мусцимола было изменение фазовой структуры дыхательного цикла за счет уменьшения времени обеих фаз дыхания. Максимальное уменьшение длительности вдоха регистрировалось в период с 5-й по 50-ю мин после инъекции (рис. 1), при этом вдох укорачивался от 185,55±32,95 (исходный уровень) до 150,37±40,63 мс, или на 18,9 % (p<0,05; ANOVA, Dunnett’s-test). В изменениях длительности выдоха тенденция укорочения имела большую выраженность и достигала статистической значимости уже с 1-й мин после микроинъекции мусцимола, что может отражать особую роль
ГАМКА-рецепторов в модуляции активности экспираторных нейронов КБ. Как видно из рис. 1, наибольшее изменение фазы выдоха было приурочено к 35-50-й мин экспозиции, когда параметр снижался от 779,06±170,58 (исходное значение) до 597,97±145,98 мс, а наблюдаемый при этом эффект удерживался на уровне 23,2 % (p<0,05; ANOVA, Tukey-test). То есть вызываемые мусцимолом отклонения фазы выдоха характеризовались большей устойчивостью во времени, сохраняясь до конца регистрации.
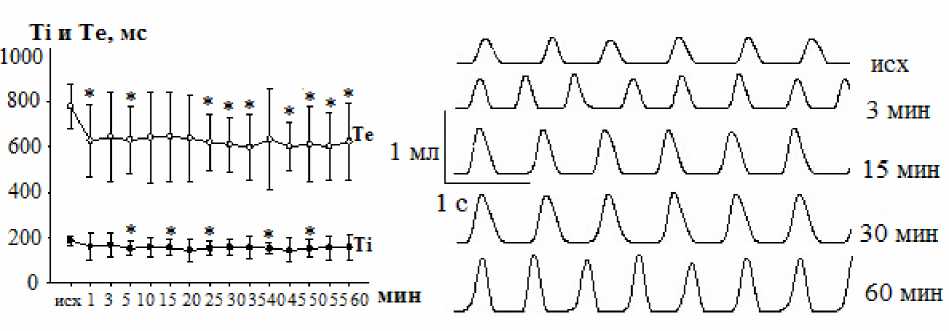
Рис. 1. Изменение длительности вдоха (Ti) и выдоха (Te) после микроинъекции 10-6 М раствора мусцимола в КБ крыс.
Справа – спирограммы.
Обозначения:
исх – исходный уровень; – достоверность отклонений от исходного уровня (p<0,05)
В результате уменьшения длительности вдоха и выдоха при инъекциях мусцимола в КБ был зарегистрирован некоторый рост ЧД. В целом ЧД увеличивалась от 71,10±5,24 (исходный фон) до 83,40±11,92 мин-1 (60-я мин экспозиции), что составляло 17,3 %. Динамика роста ЧД у животных в условиях активации ГАМК А -рецепторов области КБ мусцимолом наглядно представлена оригинальными спирограммами на рис. 1.
Анализ спирограмм показал, что микроинъекции мусцимола в КБ вызывали увеличение ДО. Исходное значение ДО равнялось 0,25±0,03 мл, на 1-й мин достигало 0,31±0,05 мл, что соответствовало приросту на 24,0 % (p<0,05; Dunnett’s-test; Tukey-test), в дальнейшем эффект мог увеличиваться. Сочетание увеличения ЧД и ДО определило направленность отклонений такого показателя, как МОД. До микроинъекции мусцимола в КБ величина МОД равнялась 17,65±4,63 мл/мин, на 1-й мин воздействия она возрастала до
23,45±7,65 мл/мин, а на 45-й мин – до 25,12±8,69 мл/мин. Выраженность эффекта при этом составляла соответственно 32,9 и 42,3 % (p<0,05; парный t-test, Dunnett’s-test).
В полном соответствии с увеличением глубины дыхания, начиная с 1-й мин воздействия ГАМК А -агониста на КБ, отмечалось повышение амплитуды активности инспираторных мышц. Причем на ЭМГ НММ амплитуда осцилляций увеличивалась на 25,4 % (p<0,05; ANOVA), что было почти в 2 раза сильнее, чем реакции диафрагмы. Изменения временных параметров ЭМГ совпадали с динамикой длительности фаз дыхания, т.е. при введении мусцимола в КБ происходило укорочение ДЗ (на 16,2 и 23,4 %; p<0,01) и МЗИ (на 27,6 и 28,1 %; p<0,01) на ЭМГ диафрагмы и НММ. Иллюстрацией наблюдаемых реакций инспираторных мышц служат оригинальные ЭМГ, зарегистрированные у крыс в исходном состоянии и в разные сроки после микроинъекции мусцимола в КБ (рис. 2).
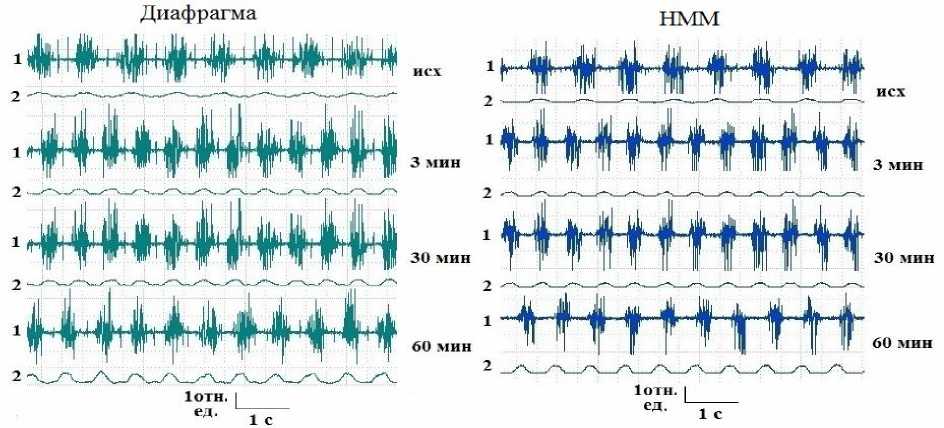
Рис. 2. Суммарные (1) и интегрированные (2) ЭМГ диафрагмы и НММ, зарегистрированные до и в разные сроки после микроинъекции 10-6 М раствора мусцимола в КБ крыс
При микроинъециях в КБ раствора ГАМК А -блокатора бикукуллина изменения большинства показателей паттерна дыхания, а также отдельных параметров ЭМГ инспираторных мышц имели характер, противоположный реакциям на введение ГАМК А -аго-ниста мусцимола. Главным отличием респираторных эффектов блокады ГАМК А -рецеп-торов КБ от эффектов их активации было пролонгирование инспираторной и экспираторной фаз дыхательного цикла.
Длительность вдоха достоверно увеличивалась с 1-й мин после микроинъекции бикукуллина, и этот эффект сохранялся в течение всего времени наблюдений. Именно на 1-й мин воздействия ГАМК А -блокатора отмечалась максимальная реакция, которая составляла 16,1 % (p<0,05; парный t-test) от исходного фона. В указанное время вдох увеличивался от 145,90±37,02 до 169,46±48,14 мс (рис. 3). Длительность выдоха после введения бикукуллина в КБ также увеличивалась, но этот эффект нарастал во времени. До введения бикукуллина длительность экспирации составляла 618,74±109,64 мс, на 30-й мин – 720,50±137,43 мс, а на 45-й мин достигала 774,44±133,48 мс, что соответствовало увеличению на 25,2 % (p<0,05; парный t-test) от исходного значения.
ЧД в ответ на инъекцию бикукуллина в КБ снижалась в среднем на 12,7 % (p<0,05; парный t-test) в интервале с 5-й по 30-ю мин (рис. 3). Отмечаемая реакция, как и в случае действия агониста, формировалась за счет преимущественного изменения длительности экспираторной фазы.
Изменения объемов дыхания на этом фоне имели бифазический характер. Так, в первые минуты после микроинъекции бикукуллина в КБ величины ДО и МОД отклонялись от исходных значений главным образом в сторону уменьшения (1–15-я мин), а в последующие сроки (20–60-я мин) эти параметры начинали меняться в сторону увеличения, причем наблюдаемые отклонения не достигали достоверного уровня. То есть выраженность изменений объемных параметров паттерна дыхания при блокаде ГАМК А -рецеп-торов КБ была заметно ниже, чем при их активации.
Амплитуда осцилляций на ЭМГ диафрагмы при инъекциях бикукуллина в КБ достоверно увеличивалась на 21,8 % (p<0,01; парный t-test). Что касается амплитуды ЭМГ НММ, то она, напротив, уменьшалась на 12,1 % (p<0,05; парный t-test). Также отсутствовало совпадение в характере изменений временных параметров ЭМГ диафрагмы и
НММ. Так, для ЭМГ диафрагмы было характерно нарастающее по мере действия ГАМКА-блокатора увеличение ДЗ и МЗИ, которые в начале наблюдений возрастали соответственно на 19,6 и 13,5 % (p<0,05; парный t-test). На ЭМГ межреберных мышц отмеча- лось укорочение ДЗ и МЗИ в среднем на 12,8 и 10,0 % (p<0,05; парный t-test), при этом изменения МЗИ сохранялись на достоверном уровне в течение всего времени действия ГАМКА-блокатора.
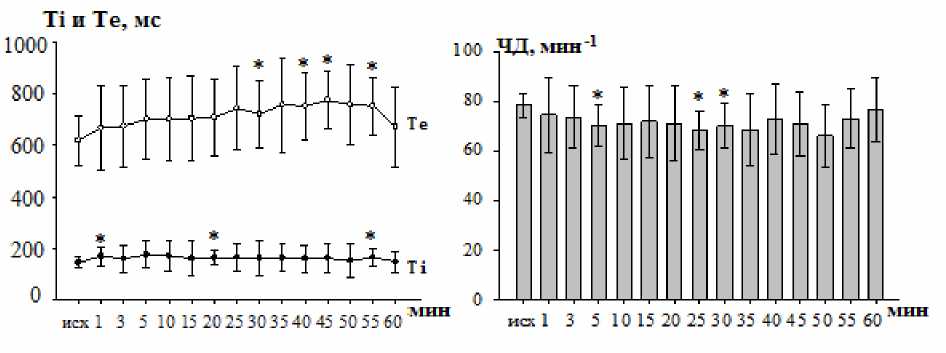
Рис. 3. Изменение длительности вдоха (Ti), выдоха (Te) и частоты дыхания после микроинъекции 10-6 М раствора бикукуллина в КБ крыс.
Обозначения: исх – исходный уровень; * – достоверность отклонений от исходного уровня (p<0,05)
Реакции внешнего дыхания и инспираторных мышц, наблюдаемые при активации и блокаде ГАМКА-рецепторов КБ, свидетельствуют об их участии на уровне указанного отдела дыхательного центра в опосредовании модулирующих влияний эндогенной ГАМК на частоту и глубину дыхания у половозрелых крыс. Обращает внимание, что отклонения большинства анализируемых параметров спирограмм и ЭМГ дыхательных мышц при изменении уровня активности ГАМКА-рецеп-торов КБ развивались с относительно короткими латентными периодами. Это, вероятно, объясняется принадлежностью ГАМКА-ре-цепторов к ионотропным мембранным сайтам, которые характеризуются высокой скоростью активации [3, 7, 8]. Также следует отметить, что ГАМКА-агонист мусцимол при инъекциях в КБ вызывал значительное укорочение фазы выдоха и коррелирующее с этим уменьшение межзалповых интервалов на ЭМГ инспираторных мышц. Это служит свидетельством важной роли ГАМКА-рецеп-торов КБ в регуляции ритмики дыхания посредством модуляции активности экспираторных нейронов, широко представленных в области КБ [15, 25, 26]. Тесная связь ГАМКА-рецепторов с регуляцией экспираторной фазы была ранее показана в условиях in vitro при регистрации инспираторной и экспираторной активности, отводимой от спинальных корешков у препаратов мозгового ствола – спинного мозга новорожденных крыс при воздействии бикукуллина и пикротоксина [18].
В плане конкретизации роли ГАМКА-рецепторов КБ в респираторном контроле представляют интерес результаты их блокады бикукуллином. Необходимо подчеркнуть, что выключение ГАМКА-рецепторов КБ не вызывало закономерных изменений объемных параметров паттерна внешнего дыхания, но в то же время приводило к достоверному изменению фаз дыхательного цикла. Этот эффект был противоположен действию мусцимола, т.е. проявлялся пролонгированием, причем особенно выраженно менялась фаза выдоха, что можно связывать с блокадой постсинаптических ГАМКА-рецепторов. Согласно литературными данным, у взрослых млекопитающих бикукуллин уменьшает величину тормозного постсинаптического потенциала во второй фазе экспирации (фазе Е2) у инспираторных и постинспираторных нейронов, во время всей экспирации – у поздних инспираторных нейронов, а во время инспирации – у экспираторных нейронов [17]. На- личие ГАМКА-рецепторов, чувствительных к бикукуллину и способных менять нейрональную возбудимость, было показано в опытах на препаратах мозгового ствола для экспираторных нейронов комплекса пре-Бетцин-гера [23].
На наш взгляд, наблюдаемое при инъекции бикукуллина в КБ увеличение длительности фаз вдоха и выдоха может быть обусловлено тем, что при блокаде ГАМК А -рецепторов меняется активность ритмогенерирующих респираторных нейросетей и интенсивность разрядов премоторных и моторных нейронов дыхательных мышц. Следует заметить, что другими исследователями факт участия ГАМК А -рецепторов в регуляции активности межреберных мышц у крыс был продемонстрирован в условиях in vitro [18]. В наших опытах это подтверждается снижением ЧД и характером отклонений амплитудных параметров ЭМГ диафрагмы и НММ. Надо подчеркнуть, что изменения амплитуды ЭМГ диафрагмы по направленности отклонений (увеличение) при действии бикукуллина на КБ совпадали с эффектом действия мусцимола, тогда как амплитуда ЭМГ НММ при введении бикукуллина и мусцимола менялась противоположным образом (уменьшалась и увеличивалась соответственно). В целом, полученные данные дают основание считать, что при блокаде ГАМК А -рецепторов КБ в большей степени меняется функциональное состояние бульбарных механизмов, управляющих активностью инспираторных межреберных мышц. Различный характер отклонений амплитуды залповых разрядов диафрагмы и НММ на фоне блокады ГАМК А -рецепторов может отчасти объяснить отсутствие закономерных реакций со стороны объемных параметров спирограмм.
Оценивая респираторные эффекты, вызываемые инъекциями мусцимола и бикукуллина, можно указать, что эффективность и результат их действия зависят как от состояния ионных каналов, в которых находятся ГАМКА-рецепторы [3], так и от набора субъединиц в рецепторах [8]. Субъединичный состав ГАМКА-рецепторов, а также их вне- и внутрисинаптическая локализация определяют физиологические свойства и кинетику мембранно-ионных процессов и, таким образом, конечный результат взаимодействия с агонистами и блокаторами [7, 14].
Заключение. На основании полученных данных можно заключить, что ГАМК А -ре-цепторы области КБ в большей степени включены в процессы формирования фазовой структуры дыхательного цикла и в меньшей степени – в механизмы регуляции глубины дыхания. С учетом того что при стимуляции и блокаде ГАМК А -рецепторов КБ какой-либо дезорганизации ритма дыхания у экспериментальных животных не наблюдалось, следует полагать, что у взрослых крыс действие лиганда через ионотропные ГАМК А -рецепто-ры КБ сводится не столько к формированию ритма, сколько к модуляции его частоты.
-
1. Александрова Н. П. Влияние гамма-аминомасляной кислоты на инспираторно-тормозящий рефлекс Геринга–Брейера / Н. П. Александрова, В. Г. Александров, Т. Г. Иванова // Рос. фи-зиол. журн. им. И. М. Сеченова. – 2008. – Т. 94, № 12. – С. 1356–1364.
-
2. Ведясова О. А. Реакции дыхания при микроинъекции ГАМК и пенициллина в различные отделы вентральной респираторной группы / О. А. Ведясова, А. М. Ковалев // Бюл. экспериментальной биологии и медицины. – 2012. – Т. 153, № 2. – С. 137–140.
-
3. Калуев А. В. Как организован хлорный ионофор ГАМК-А рецептора? / А. В. Калуев // Нейронауки. – 2006. – № 3 (5). – С. 31–43.
-
4. Реакции дыхания на введение агонистов ГАМКергических рецепторов / И. А. Тараканов [и др.] // Бюл. сибирской медицины. – 2005. – Т. 4 (прил. 1). – С. 47–48.
-
5. Alheid G. F. The chemical neuroanatomy of breathing / G. F. Alheid, D. R. McCrimmon // Respir. Physiol. Neurobiol. – 2008. – Vol. 164. – P. 3–11.
-
6. Association of GABA(B)R1 receptor gene polymorphism with obstructive sleep apnea syndrome / Y. A. Bayazit [et al.] // ORL. J. Otorhinolaryngol. Relat. Spec. – 2007. – Vol. 69, № 3. – Р. 190–197.
-
7. Banks M. I. Kinetic differences between synaptic and extrasynaptic GABA(A) receptors in CA1 pyramidal cells / M. I. Banks, R. A. Pearce // J. Neurosci. – 2000. – Vol. 20, № 3. – P. 937–948.
-
8. Bianchi M. T. Structural determinants of fast desensitization and desensitization-deactivation coupling in GABAa receptors / M. T. Bianchi, K. F. Haas, R. L Macdonald // J. Neurosci. – 2001. – Vol. 21, № 4. – P. 1127–1136.
-
9. Blockade of synaptic inhibition within the pre-Bötzinger complex in the cat suppresses respira-
tory rhythm generation in vivo / О. Pierrefiche [et al.] // J. Physiol (Lond.). – 1998. – Vol. 509. – Р. 245–254.
-
10. Bou-Flores C . Gap junctions and inhibitory synapses modulate inspiratory motoneuron synchronization / C. Bou-Flores, A. J. Berger // J. Neurophysiol. – 2001. – Vol. 85. – P. 1543–1551.
-
11. Chitravanshi V. C. Phrenic nerve responses to chemical stimulation of the subregions of ventral medullary neuronal group in the rat / V. C. Chitra-vanshi, H. N. Sapru // Brain Res. – 1999. – Vol. 821, № 2. – P. 443–446.
-
12. Differential ontogeny of GABA(B)-receptor-mediated pre- and postsynaptic modulation of GABA and glycine transmission in respiratory rhythmgenerating network in mouse / W. Zhang [et al.] // J. Physiol. – 2002. – Vol. 540. – P. 435–446.
-
13. Duffin J. Functional synaptic connections among respiratory neurons / J. Duffin, G.-F. Tian, J. N. Peever // Pespir. Physiol. – 2000. – Vol. 122. – P. 237–246.
-
14. Dynamic mobility of functional GABAA receptors at inhibitory synapses / P. Thomas [et al.] // Nat. Neurosci. – 2005. – Vol. 8. – P. 889–897.
-
15. Feldman J. L. Looking for inspiration: new perspectives on respiratory rhythm / J. L. Feldman, C. A. Del Negro // Nat. Rev. Neurosci. – 2006. – Vol. 7. – Р. 232–242.
-
16. Glycinergic pacemaker neurons in pre-Bötzinger complex of neonatal mouse / C. Morgado-Valle [et al.] // J. Neurosci. – 2010. – Vol. 30. – P. 3634–3639.
-
17. Haji A. Neuropharmacology of control of respiratory rhythm and pattern in mature mammals / A. Haji, R. Takeda, M. Okazaki // Pharmacol. Ther. – 2000. – Vol. 86. – P. 277– 304.
-
18. Iizuka M. GABAA and glycine receptors in regulation of intercostal and abdominal expiratory activity in vitro in neonatal rat / M. Iizuka
// J. Physiol. – 2003. – Vol. 551, № 2. – Р. 617–633.
-
19. Respiratory responses evoked by blockades of ionotropic glutamate receptors within the Bötzinger complex and the pre-Bötzinger complex of the rabbit / D. Mutolo [et al.] // Eur. J. Neurosci. – 2005. – Vol. 21. – Р. 122–134.
-
20. Respiratory responses induced by blockades of GABA and glycine receptors within the Bötzinger complex and the pre-Bötzinger complex of the rabbit / F. Bongianni [et al.] // Brain Res. – 2010. – Vol. 1344. – P. 134–147.
-
21. Ritter B. Early postnatal maturation of GABAA-mediated inhibition in the brainstem respiratory rhythm-generating network of the mouse / B. Ritter, W. Zhang // Eur. J. Neurosci. – 2000. – Vol. 12. – P. 2975–2984.
-
22. Role of inhibition in respiratory pattern generation / W. A. Janczewski [et al.] // J. Neurosci. – 2013. – Vol. 33, № 13. – P. 5454–5465.
-
23. Shao X. M. Respriatory rhythm generation and synaptic inhibition of expiratory neurons in pre-Botzinger complex: differential roles of glycinergic and GABАergic neural transmission / X. M. Shao, J. L. Feldman // J. Neurophysiol. – 1997. – Vol. 77. – P. 1853–1860.
-
24. St-Jacques R. Transient, reversible apnoea following ablation of the pre-Botzinger complex in rats / R. St-Jacques, W. M. St-John // J. Physiol. – 1999. – Vol. 520, № 1. – P. 303–314.
-
25. Sun Q. Firing patterns of pre-Botzinger and Botzinger neurons during hypocapnia in the adult rat / Q. Sun, A. K. Goodchild, P. M. Pilowsky // Brain Res. – 2001. – Vol. 903. – P. 198–206.
-
26. Tian G.-F. Botzinger-complex, bulbospinal expiratory neurons monosynaptically inhibit ventral-group respiratory neurones in the decerebrate rat / G.-F. Tian, J. H. Peever, J. Duffin // Exp. Brain. Res. – 1999. – Vol. 124. – P. 173–180.
ROLE OF THE BETZINGER COMPLEX GABAA-RECEPTORS IN RESPIRATORY CONTROL IN RATS
N.G. Manshina, O.A. Vedyasova
Samara State University
Список литературы Роль ГАМК А-рецепторов комплекса Бетцингера в регуляции дыхания у крыс
- Александрова Н. П. Влияние гамма-аминомасляной кислоты на инспираторно-тормозящий рефлекс Геринга-Брейера/Н. П. Александрова, В. Г. Александров, Т. Г. Иванова//Рос. физиол. журн. им. И. М. Сеченова. -2008. -Т. 94, № 12. -С. 1356-1364.
- Ведясова О. А. Реакции дыхания при микроинъекции ГАМК и пенициллина в различные отделы вентральной респираторной группы/О. А. Ведясова, А. М. Ковалев//Бюл. экспериментальной биологии и медицины. -2012. -Т. 153, № 2. -С. 137-140.
- Калуев А. В. Как организован хлорный ионофор ГАМК-А рецептора?/А. В. Калуев//Нейронауки. -2006. -№ 3 (5). -С. 31-43.
- Реакции дыхания на введение агонистов ГАМКергических рецепторов/И. А. Тараканов [и др.]//Бюл. сибирской медицины. -2005. -Т. 4 (прил. 1). -С. 47-48.
- Alheid G. F. The chemical neuroanatomy of breathing/G. F. Alheid, D. R. McCrimmon//Respir. Physiol. Neurobiol. -2008. -Vol. 164. -P. 3-11.
- Association of GABA(B)R1 receptor gene polymorphism with obstructive sleep apnea syndrome/Y. A. Bayazit [et al.]//ORL. J. Otorhinolaryngol. Relat. Spec. -2007. -Vol. 69, № 3. -Р. 190-197.
- Banks M. I. Kinetic differences between synaptic and extrasynaptic GABA(A) receptors in CA1 pyramidal cells/M. I. Banks, R. A. Pearce//J. Neurosci. -2000. -Vol. 20, № 3. -P. 937-948.
- Bianchi M. T. Structural determinants of fast desensitization and desensitization-deactivation coupling in GABAa receptors/M. T. Bianchi, K. F. Haas, R. L Macdonald//J. Neurosci. -2001. -Vol. 21, № 4. -P. 1127-1136.
- Blockade of synaptic inhibition within the pre-Bötzinger complex in the cat suppresses respiratory rhythm generation in vivo/О. Pierrefiche [et al.]//J. Physiol (Lond.). -1998. -Vol. 509. -Р. 245-254.
- Bou-Flores C. Gap junctions and inhibitory synapses modulate inspiratory motoneuron synchronization/C. Bou-Flores, A. J. Berger//J. Neurophysiol. -2001. -Vol. 85. -P. 1543-1551.
- Chitravanshi V. C. Phrenic nerve responses to chemical stimulation of the subregions of ventral medullary neuronal group in the rat/V. C. Chitravanshi, H. N. Sapru//Brain Res. -1999. -Vol. 821, № 2. -P. 443-446.
- Differential ontogeny of GABA(B)-receptor-mediated pre-and postsynaptic modulation of GABA and glycine transmission in respiratory rhythm-generating network in mouse/W. Zhang [et al.]//J. Physiol. -2002. -Vol. 540. -P. 435-446.
- Duffin J. Functional synaptic connections among respiratory neurons/J. Duffin, G.-F. Tian, J. N. Peever//Pespir. Physiol. -2000. -Vol. 122. -P. 237-246.
- Dynamic mobility of functional GABAA receptors at inhibitory synapses/P. Thomas [et al.]//Nat. Neurosci. -2005. -Vol. 8. -P. 889-897.
- Feldman J. L. Looking for inspiration: new perspectives on respiratory rhythm/J. L. Feldman, C. A. Del Negro//Nat. Rev. Neurosci. -2006. -Vol. 7. -Р. 232-242.
- Glycinergic pacemaker neurons in pre-Bötzinger complex of neonatal mouse/C. Morgado-Valle [et al.]//J. Neurosci. -2010. -Vol. 30. -P. 3634-3639.
- Haji A. Neuropharmacology of control of respiratory rhythm and pattern in mature mammals/A. Haji, R. Takeda, M. Okazaki//Pharmacol. Ther. -2000. -Vol. 86. -P. 277-304.
- Iizuka M. GABAA and glycine receptors in regulation of intercostal and abdominal expiratory activity in vitro in neonatal rat/M. Iizuka//J. Physiol. -2003. -Vol. 551, № 2. -Р. 617-633.
- Respiratory responses evoked by blockades of ionotropic glutamate receptors within the Bötzinger complex and the pre-Bötzinger complex of the rabbit/D. Mutolo [et al.]//Eur. J. Neurosci. -2005. -Vol. 21. -Р. 122-134.
- Respiratory responses induced by blockades of GABA and glycine receptors within the Bötzinger complex and the pre-Bötzinger complex of the rabbit/F. Bongianni [et al.]//Brain Res. -2010. -Vol. 1344. -P. 134-147.
- Ritter B. Early postnatal maturation of GABAA-mediated inhibition in the brainstem respiratory rhythm-generating network of the mouse/B. Ritter, W. Zhang//Eur. J. Neurosci. -2000. -Vol. 12. -P. 2975-2984.
- Role of inhibition in respiratory pattern generation/W. A. Janczewski [et al.]//J. Neurosci. -2013. -Vol. 33, № 13. -P. 5454-5465.
- Shao X. M. Respriatory rhythm generation and synaptic inhibition of expiratory neurons in pre-Botzinger complex: differential roles of glycinergic and GABАergic neural transmission/X. M. Shao, J. L. Feldman//J. Neurophysiol. -1997. -Vol. 77. -P. 1853-1860.
- St-Jacques R. Transient, reversible apnoea following ablation of the pre-Botzinger complex in rats/R. St-Jacques, W. M. St-John//J. Physiol. -1999. -Vol. 520, № 1. -P. 303-314.
- Sun Q. Firing patterns of pre-Botzinger and Botzinger neurons during hypocapnia in the adult rat/Q. Sun, A. K. Goodchild, P. M. Pilowsky//Brain Res. -2001. -Vol. 903. -P. 198-206.
- Tian G.-F. Botzinger-complex, bulbospinal expiratory neurons monosynaptically inhibit ventral-group respiratory neurones in the decerebrate rat/G.-F. Tian, J. H. Peever, J. Duffin//Exp. Brain. Res. -1999. -Vol. 124. -P. 173-180.
