Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений

Автор: Чудинова Лариса Алексеевна, Суворов Василий Иванович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2011 года.
Бесплатный доступ
Показано, что ключевым механизмом кросс-адаптации является накопление разнообразных низкомолекулярных соединений (моносахариды, пролин, полиамины), которые отличаются полифункциональностью и могут обслуживать неспецифические реакции растений на стрессоры различной природы.
Низкомолекулярные протекторы (пролин, глюкоза, полиамины), гипертермия, засоление, прогрев закаливающей температурой, перекрестная адаптация
Короткий адрес: https://sciup.org/147204526
IDR: 147204526 | УДК: 581.1:632.122.1
Role of some low-molecular compounds in the mechanism of cross adaptation of plants
The accumulation of various low-molecular compounds (monosaccharides, proline, polyamines) which differ polyfunctionality is a mechanizm of cross-country-adaptation and can by nonspecific reactions of plants on stressors the various nature.
Текст научной статьи Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений
Эксперименты по перекрестной адаптации являются убедительным доказательством функционирования общих механизмов устойчивости растений к действию различных стрессоров.
Показано, что у ряда растений тепловой шок оказывает протекторный эффект к последующему тепло-, холодо-, солевому воздействию и водному дефициту (Кузнецов и др., 1992; Таланова и др., 1993; Акимова и др., 2001).
Нами в целой серии экспериментов с разными растениями (горох, пшеница, гречиха, кукуруза и др.) было показано, что предварительный прогрев закаливающей температурой 35-37 ° С в течение 1.5–3 ч. снижает рост ингибирующий эффект действия последующего засоления или гипертермии (Чудинова, Суворов, Тюмина, 2005; Чудинова, Суворов, 2008, 2009). Развитие устойчивости было зафиксировано уже через 1–3 ч. Эффекты, вызываемые кратковременным нагревом растений, могут сохраняться на протяжении длительного времени (Акимова и др., 2001).
Локальное воздействие повышенной температуры только на корень индуцировало рост тепло- и солеустойчивости непрогретых листьев. Это указывает на передачу сигнала о тепловом воздействии в пространственно удаленные органы, что вызывает в них развитие неспецифических изменений, направленных на повышение общей устойчивости.
В настоящее время основное внимание уделяется изучению биохимических и молекулярногенетических аспектов устойчивости растений к неблагоприятным факторам, в частности, к одно- временному действию высокой температуры и засоления. Имеются указания на важное значение низкомолекулярных протекторов и осмопротекторов, таких как некоторые моносахариды, аминокислоты, полиамины. Однако в этом аспекте данный вопрос исследован недостаточно.
Целью нашей работы было изучение некоторых низкомолекулярных протекторов (НМП) растений в условиях комбинированного и раздельного действия засоления и гипертермии.
Объекты и методы исследований
Объект исследований – проростки пшеницы мягкой ( Triticum aestivum L . ) сорта Иргина. Трехдневные проростки неделю выращивали методом водной культуры на питательной смеси Кнопа (контроль), а затем подвергали их тепловому и солевому воздействиям.
Основные варианты опытов: 1) засоление 1.0% NaCl – 3 ч.; 2) тепловая обработка закаливающей температурой (37 ° С) - 3 ч.; 3) гипертермия (45 ° С) – 3 ч.
У 12-дневных проростков в надземной части и корнях анализировали: содержание пролина – по методу L.S. Bates (Bates, Waidren, Tear, 1973), полиаминов (путресцина и спермидина) – методом тонкослойной хроматографии (Чудинов, Чудинова, Коробов, 1984), содержание глюкозы определяли по методу Бертрана. Повторность эксперимента – трехкратная.
Результаты и их обсуждение
Считается, что среди всех наиболее вероятных кандидатов на участие в развитии устойчивости
растительных клеток к засолению и гипертермии является пролин, обладающий преимущественно защитной функцией. Аккумуляция пролина является одной из неспецифических реакций растений на действие различных стрессоров. Содержание этой аминокислоты возрастает в десятки и сотни раз в условиях засухи, засоления, действия высоких и низких температур и других повреждающих факторов (Кузнецов, Шевякова, 1999; Hare, Cress, Sta-den, 1999).
Сложилось мнение, что свободный пролин при стрессе – осмопротектор, обладает полифункцио-нальным биологическим эффектом, является стабилизатором макромолекул и мембран, дополнительным источником энергии и азота, антиоксидантом и др. (Кузнецов, Шевякова, 1999).
Ранее нами также была показана аккумуляция пролина в побегах и корнях гороха под влиянием высокой температуры (45○C) и засоления (Чудинова, Суворов, 2009). Как показывают данные, представленные на рис. 1, наибольшая аккумуляция пролина (в 4–5 раз по сравнению с контролем) наблюдалась при одновременном действии гипертермии и засоления. При этом предварительный прогрев закаливающей температурой незначительно менял динамику пролина.
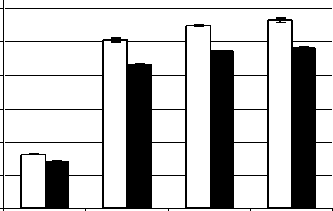
Вариант 1 Вариант 2 Вариант 3 Вариант 4
Рис. 1 . Накопление пролина в проростках пшеницы в ответ на комбинированное действие гипертермии и засоления (мг на г сухой массы): вариант 1 – контроль; вариант 2 – гипертермия и засоление без предварительного прогрева; вариант 3 - предварительный прогрев корней (37 ° C 3 ч.) и последующее воздействие гипертермии и засоления; вариант 4 – предварительный общий прогрев растений (37 ° C 3 ч.) и последующее воздействие гипертермии и засоления
Существует мнение (Кузнецов и др., 1990), что тепловой шок сам по себе не индуцирует синтез пролина, а лишь делает растения компетентными ответить на последующее засоление накоплением данного осмолита.
Локальный предварительный прогрев корней положительно сказывается на аккумуляции пролина в проростках пшеницы при последующем солевом и температурном воздействии, что, по-видимому, обусловлено развитием преимущест- венно неспецифических реакций (Чудинова, Суворов, 2009).
До сих пор остается неизвестной природа рецептора, с помощью которого растительная клетка воспринимает температурный сигнал. Предполагают, что при изменении вязкости мембран может происходить открывание кальциевых каналов (Карпец, Колупаев, 2009) и активация кальций зависимых путей трансдукции сигнала в геном (Трунова, 2007). Важными интермедиатами могут быть также активные формы кислорода и продукты перекисного окисления липидов.
Наши исследования показали (рис. 2), что предварительная обработка закаливающей температурой приводила к значительному возрастанию содержания глюкозы, особенно при высокой напряженности стрессового воздействия (вар. 4). Небольшое изменение количества глюкозы наблюдалось при монодействии засоления и гипертермии (вар. 5 и 6), однако при совместном действии этих стрессоров содержание глюкозы резко снижалось (вар. 7).
Корень Побег
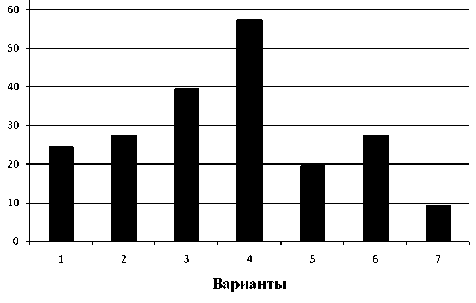
Рис. 2. Содержание глюкозы в проростках пшеницы (мг на г сырой массы):
1 - контроль; 2 - 37 ° С и засоление; 3 - 37 ° С и гипертермия; 4 - 37 ° С с последующей гипертермией и засолением; 5 – засоление; 6 – гипертермия; 7 – гипертермиия и последующее засоление
Повышенный уровень глюкозы у опытных растений может выполнять важную протекторную роль и наряду с другими факторами позволяет растениям сохранить достаточную интенсивность роста в условиях стресса. Этому способствуют такие свойства моносахаридов, как способность повышать стабильность биомембран путем формирования слабых взаимодействий с кислородными атомами фосфатов фосфолипидов, их антиденатура-ционное влияние на белки и антиоксидантное действие (Карпец, Колупаев, 2009). Накапливающиеся углеводы могут также обеспечивать поддержание осмотического статуса, не оказывая вредного влияния на ферменты и мембраны (Кафи, Стюарт, Борланд 2003). С недавних пор, помимо протекторных функций, некоторым углеводам отводят и сигнальную роль (Rolland, Sheen, 2005).
В настоящее время полиамины (путресцин, кадаверин, спермидин, спермин) рассматриваются в качестве низкомолекулярных мультифункциональ-ных регуляторов физиологических процессов, как новый класс вторичных посредников, а также маркеров центров с высокой метаболитической активностью. Установлено, что полиамины участвуют в дистанционной передаче стрессорного сигнала у растений (Ozturk, Demir, 2003; Кузнецов, Радюки-на, Шевякова, 2006).
Проведенный нами анализ содержания полиаминов в проростках пшеницы (рис. 3) показал значительное увеличение их количества при засолении и некоторое снижение в условиях гипертермии. Предварительный прогрев проростков закаливающей температурой способствовал поддержанию повышенного уровня полиаминов. Возможно, что это определяется высокой активностью их синтетических ферментов – декарбоксилаз, которые ха акте из ются си ьной ин ибе ьностью
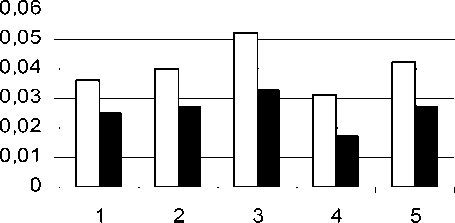
Путресцин Спермидин инов в сух. массы):
каливающая температура (37°C, 3 ч.) и последующее засоление; 4 - гипертермия; 5 - (37°C, 3 ч.) и последующая гипертермия
В целом, полученные нами результаты и данные в литературе дают основание считать, что ключевым механизмом кросс-адаптации является накопление разнообразных низкомолекулярных соединений, которые отличаются полифункциональностью и могут обслуживать неспецифические реакции растений на стрессоры различной природы. Активность ферментов, ответственных за накопление низкомолекулярных протекторов, может возрастать в результате изменения экспрессии, кодирующих их генов под влиянием вторичных мессенджеров (АФК, ионов кальция) или стрессовых гормонов (абсцизовой и салициловой кислот, этилена).
Синтез стрессовых белков также относится к малоспецифическим реакциям, поскольку одни и те же стрессовые белки накапливаются при гипер- и гипотермии, засолении, засухе и др. факторах, а изменение экспрессии генов стрессовых белков вызывают одни и те же посредники, в частности АФК (Суворов, Чудинова, 2009). Главная функция всех стресс-белков – это защита структуры функциональных белков, что важно при различных повреждениях. В то же время синтез определенных групп стресс-белков (например, поздних БТШ) может быть достаточно специфической реакцией на определенный фактор. В процессе адаптации соотношение между неспецифическими и специфическими реакциями изменяется в пользу последних.
Список литературы Роль некоторых низкомолекулярных соединений в механизме перекрестной адаптации растений
- Акимова Т.В. и др. Повышение теплоустойчивости листьев при локальном прогреве проростков//Физиол. раст. 2001. Т. 48, № 4. С. 584-588.
- Карпец Ю.В., Колупаев Ю.Е. Ответ растений на гипертермию: молекулярно-клеточные аспекты//Вестн. Харьков. нац. аграр. ун-та. Сер. биология. 2009. Вып. 1 (16). С. 19-38.
- Кафи М., Стюарт В.С., Борланд А.М. Содержание углеводов и пролина в листьях, корнях и апексах пшеницы, устойчивых и чувствительных к засолению//Физиол. раст. 2003. Т. 50, № 2. С. 174-182.
- Кузнецов Вл.В. и др. Общие системы устойчивости хлопчатника к засолению и высокой температуре: факты и гипотезы//Физиол. раст. 1990. Т. 37, № 5. С. 987-996.
- Кузнецов Вл.В. и др. Стрессовые белки и фитогормоны при адаптации растений Cucumis sativus L. к почвенной засухе//Докл. АН СССР. 1992. Т. 322. С. 204-207.
- Кузнецов Вл.В., Радюкина Н.Л., Шевякова Н.И. Полиамины при стрессе: биологическая роль, метаболизм и регуляция//Физиол. раст. 2006. Т. 53, № 5. С. 658-683.
- Кузнецов Вл.В., Шевякова Н.И. Пролин при стрессе: биологическая роль, метаболизм, регуляция//Физиол. раст. 1999. Т. 46, № 2. С. 321-336.
- Суворов В.И., Чудинова Л.А. Влияние раздельного и комбинированного действия гипертермии и засоления на активность некоторых антиоксидантных ферментов в проростках гороха//Экология в высшей школе: синтез науки и образования: материалы Всерос. науч.-практ. конф. Челябинск, 2009. Ч. 1. С. 115-118.
- Таланова В.В. и др. Раздельное и комбинированное действие засоления и закаливающих температур на растения//Физиол. раст. 1993. Т. 40, № 4. С. 584-588.
- Трунова Т.И. Растение и низкотемпературный стресс. М.: Наука, 2007. 55 с.
- Чудинов А.А., Чудинова Л.А., Коробов В.П. Метод определения низкомолекулярных олигоаминов в различном биологическом материале//Вопр. мед. химии. 1984. Т. 30, вып. 4. С. 127-132.
- Чудинова Л.А., Суворов В.И., Тюмина Е.Н. Влияние раздельного и комбинированного действия засоления и гипертермии на полиаминсинтезирующую систему проростков кукурузы//Вестн. Перм. ун-та. 2005. Вып. 6. Биология. С. 71-73.
- Чудинова Л.А., Суворов В.И. Влияние гипертермии на устойчивость проростков растений к последующему засолению//Материалы регион. науч.-практ. конф. «Проблемы экологии и экологического образования Уральского Федерального округа». Челябинск, 2008. С. 120-125.
- Чудинова Л.А., Суворов В.И. Влияние предварительного прогрева на аккумуляцию пролина в проростках гороха при последующем засолении и гипертермии//Ботанические исследования на Урале: материалы регион. науч. конф. Пермь, 2009. С. 377-380.
- Bates L.S., Waidren R.P., Tear J.D. Rapid determination of free proline for water stress studies//Plant and soil. 1973. Vol. 39, № 1. P. 205-210.
- Hare P.D., Cress W.A., Staden J.V. Prolin synthesis and degradation: a model for elucidating stressrelated signal transduction//J. Exp. Bot. 1999. Vol. 50. Р. 413-434.
- Ozturk L., Demir Y. Effects of putrescine and ethephon on some oxidative stress enzyme activates and prolin content in stressed spinach leaves//Plant Growth Regul. 2003. Vol. 40. Р. 89-95.
- Rolland F., Sheen J. Sugar sensing and signalling networks in plants//Biochemical Society Transaction. 2005. Vol. 33. Р. 269-271.
