Роль полиаминов в ограничении проницаемости внешней мембраны Escherichia coli для антибиотиков

Автор: Ахова А.В., Ткаченко А.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 3, 2020 года.
Бесплатный доступ
Многие гидрофильные антибиотики поступают в клетку грамотрицательных бактерий через пориновые каналы, которые могут блокироваться полиамином кадаверином. В данной работе исследованы скорость поринового транспорта и внутриклеточная концентрация основных биогенных полиаминов в зависимости от времени экспозиции клеток Escherichia coli с фторхинолоновыми антибиотиками. Пориновая проницаемость резко снижалась в первые 2 ч. воздействия антибиотиков, после чего темп падения скорости транспорта замедлялся. Внутриклеточное содержание всех полиаминов повышалось ко 2-му ч. воздействия антибиотиков, далее концентрация путресцина и спермидина снижалась, а количество кадаверина продолжало увеличиваться. Активация экспрессии гена micF, продукт которого участвует в отрицательной регуляции количества пориновых белков OmpF, наблюдалась после 2-го ч. воздействия. Это предполагает, что ограничение транспорта антибиотиков за счет блокирования пориновых каналов является первичным защитным ответом, после чего запускаются механизмы регуляции количества поринов. Обсуждается возможная специализация полиаминов в процессе регуляции ограничения транспорта антибиотиков через пориновые каналы.
Кадаверин, путресцин, спермидин, фторхинолоны, порины
Короткий адрес: https://sciup.org/147229655
IDR: 147229655 | УДК: 579.22 | DOI: 10.17072/1994-9952-2020-3-204-209
Role of polyamines in reducing the outer membrane permeability of Escherichia coli to antibiotics
Various hydrophilic antibiotics enter the cells of gram-negative bacteria through porin channels, which can be blocked by polyamine cadaverine. In this paper, we studied the rate of porin-mediated transport and the intracellular concentration of the main biogenic polyamines of Escherichia coli cells subjected to fluoroquinolone antibiotics depending on the exposure time. Porin permeability sharply decreased in the first two hours of antibiotic exposure, after that the transport rate continued to decline, but at a lower rate. Free polyamines accumulated by the second hour of the exposure, then putrescine and spermidine declined, and the concentration of cadaverine continued to grow. Activation of micF gene, a product of which is involved in the negative control of ompF porin gene expression, was observed after the two hours of antibiotic exposure. It could be supposed that the inhibition of porin-mediated transport of antibiotics by blocking pores precedes the mechanisms regulating the number of porins. The paper discusses the probable specialization of polyamines in the regulation of porin-mediated transport of antibiotics.
Текст научной статьи Роль полиаминов в ограничении проницаемости внешней мембраны Escherichia coli для антибиотиков
цилиндр из 16 антипараллельных тяжей, окружающих обводненный канал. Характерной особенностью поринового канала является выступающая во внутреннюю часть L3 петля, формирующая сужение просвета канала в центральной части. Этот участок, называемый «глазок», играет ключевую роль в определении селективности и проницаемости канала [delaVega, Delcour, 1996; Achouak, Heulin, Pagès, 2001]. Скорость поринового транспорта определяется количеством и составом пори-новых белков, а также проницаемостью каналов.
Одной из существенных составляющих эффективности антибактериального действия препаратов является скорость их проникновения в микробную клетку. Поскольку многие водорастворимые антибиотики (фторхинолоны, β-лактамы) транспортируются в бактериальные клетки через пориновые каналы [Nikaido, 2003; Delcour, 2009], регуляция пориновой проницаемости может вносить вклад в снижение чувствительности микроорганизмов.
Ограничение проницаемости внешней мембраны широко используется грамотрицательными микроорганизмами как средство адаптации ко многим видам стресса. Порин OmpF образует поры большего размера, по сравнению с OmpC, соответственно, его транспортная активность выше. Поэтому при наличии в среде ксенобиотиков, а также в условиях высокой осмолярности и низкого рН среды, клетка использует регуляторные механизмы, направленные на ограничение содержания или транспортной активности, в первую очередь, именно этого порина [Nikaido, 2003].
Регуляция транскрипции ompF и ompC осуществляется при участии двухкомпонентной сиг-нал-проводящей системы EnvZ/OmpR. Получая сигнал из окружающей среды, EnvZ аутофосфори-лируется и передает фосфат на белок OmpR. В фосфорилированной форме OmpR выступает в качестве репрессора ompF , одновременно индуцируя транскрипцию ompC [Pratt et al., 1996; Nikaido, 2003]. Вклад в ограничение проницаемости клеточной стенки также вносит активатор генов общего стрессового ответа - RpoS, который отрицательно регулирует экспрессию ompF [Liu, Ferenci, 2001]. Посттранскрипционный уровень регуляции основан на образовании двуспиральной структуры между мРНК белка порина ompF и антисмысловой РНК micF , в результате чего мРНК становится недоступной для рибосом, и процесс трансляции блокируется [Delihas, Forst, 2001].
Важную роль в ограничении пориновой проницаемости играют полиамины, которые у грамотри-цательных организмов представлены путресцином, кадаверином и спермидином [Tabor, Tabor, 1985; delaVega, Delcour, 1996; Samartzidou, Delcour, 1999]. Эффект полиаминов реализуется в блокировке пор и стабилизации их закрытого состояния за счет взаимодействия с отрицательно заряженными участками поринов [Iyer, Delcour, 1997]. Проведенные ранее исследования показали, что эффективность экзогенных полиаминов зависит от количества аминогрупп в их молекуле и увеличивается в ряду: путресцин<кадаверин<спермидин [Tkachenko, Pozhidaeva, Shumkov, 2006]. В то же время, в естественных условиях при эндогенном образовании полиаминов основная роль в регуляции поринового транспорта отводится кадаверину [Delcour, 2009], в то время как путресцин и спермидин участвуют в регуляции экспрессии генов [Шумков и др., 2010; Igarashi, Kashiwagi, 2018].
Цель работы – изучение роли биогенных полиаминов в регуляции пориновой проницаемости E. coli в зависимости от времени экспозиции с антибиотиком.
Материалы и методы исследования
Микроорганизмы и условия культивирования . В качестве объектов использованы штаммы Escherichia coli RO91 (MC4100 λ RZ5:rpoS742::lacZ [hybr]) [Muffler et al., 1997], трансформированные плазмидой pBR322, и N9212 (GC4468 λ :micF::lacZ ) [Martin, Gillette, Rosner, 2000]. Штамм RO91 любезно предоставлен профессором Р. Хенгге (Берлинский университет им. Гумбольдта, Германия). Штамм N9212 любезно предоставлен профессором Р. Мартином (США).
Клетки E. coli , сохраняемые на скошенном Lu-ria-Bertani-агаре (“Sigma”, США), переносили в пробирки с 5 мл Luria-Bertani-бульона (“Sigma”, США) и культивировали 4‒6 ч. при 37ºC без перемешивания. Затем бактериальные клетки переносили в колбы Эрленмейера (объемом 250 мл), содержащие 50 мл LB-бульона, в соотношении 1:1000 и культивировали при 37ºC с перемешиванием 100 об/мин (термостатируемый шейкер GFL 1092 (“GFL”, Германия)) в течение 14‒16 ч. Полученную культуру разводили в свежей питательной среде до оптической плотности 0.1 и культивировали в описанных выше условиях. Исследуемые антибиотики добавляли по достижении культурой оптической плотности 0.3.
Оптическую плотность бактериальных культур (А 600 ) измеряли по величине абсорбции на 600 нм с использованием спектрофотометра UV-1650PC ("Shimadzu”, Япония).
Транспортную активность пориновых каналов внешней мембраны E. coli определяли модифицированным методом Циммермана и Росселета [Nikaido, Rosenberg, Foulds, 1983], основанным на фотометрическом измерении концентрации цефалоспоринов в среде по мере их прохождения через пориновые каналы внешней мембраны грамотри- цательных микроорганизмов и расщепления пе-риплазматической β-лактамазой.
Клетки, трансформированные плазмидой pBR322, осаждали центрифугированием (3000g, 8 мин., 0°С), удаляли надосадочную жидкость, осадок отмывали в 3 мл 10 мМ холодного фосфатного буфера, содержащего 5 мМ CaCl 2 (рН 6.0), и повторно центрифугировали в том же режиме. Полученный осадок ресуспендировали в 6 мл фосфатного буфера. Половину полученной суспензии подвергали ультразвуковой дезинтеграции (22 кГц, 10 мА, 4×15 с, на холоде), другую половину оставляли для определения скорости гидролиза цефазоли-на клетками. Озвученный экстракт центрифугировали (16000g, 20 мин, 0°С) и в количестве, эквивалентном 0.15 мг/мл сухого веса (АСБ), помещали в кювету с длиной оптического пути 1 мм, содержащую фосфатный буфер и цефазолин в концентрации 1 мМ (конечный объем – 0.4 мл). Измерение поглощения производили на спектрофотометре UV-1650 PC («Shimadzu», Япония) на длине волны 270 нм.
Коэффициент проницаемости Р (×10-5 см/с), характеризующий транспортную активность пори-новых каналов, рассчитывали, как описано ранее [Tkachenko, Pozhidaeva, Shumkov, 2006].
Количественный анализ полиаминов проводили методом ТСХ [Чудинов, Чудинова, Коробов, 1984] их дансил-производных. Полиамины экстрагировали 0.4 N хлорной кислотой из клеток E. coli , сконцентрированных центрифугированием (2 мин., 16000g). К 100 мкл хлорнокислого экстракта, доведенного до рН 9.0 раствором Na 2 CO 3 (насыщенный), добавляли 100 мкл раствора 1-диметиламинонафталин-5-сульфонилхлорида (дансил-хлорид) («Sigma», США) в ацетоне (2.7 мг/мл) и инкубировали в течение 2 ч. при 37ºС в темноте. Смесь выпаривали в концентраторе Concentrator 5301 («Eppendorf», Германия) и экстрагировали бензолом. Бензольные экстракты количественно наносили на силикагелевые пластины для тонкослойной хроматографии («Sorbfil», Россия) размером 100 x 100 мм и последовательно разделяли в двух системах растворителей: I – бензол – триэтиламин (20:2); II – бензол – метанол (10: 0.45). Высушенные хроматограммы фотографировали на цифровую камеру Olimpus С-3040 (Япония) в ультрафиолетовом свете, возбуждающем сине-зеленое свечение пятен дансил-полиаминов, величина и яркость которых были пропорциональны их концентрации. По результатам компьютерного денситометрирования снимков рассчитывали концентрацию полиаминов. Расчет количества полиаминов проводили относительно внешнего стандарта.
Уровень экспрессии генов оценивали по активности β-галактозидазы в клетках, несущих слияние промотора исследуемого гена со структурной частью гена lacZ . Активность определяли в клетках, предварительно обработанных смесью додецилсульфата натрия и хлороформа, с добавкой ор-то-нитрофенил-β-галактопиранозида (“Sigma”, США) в качестве субстрата [Miller, 1992].
Статистическая обработка результатов проведена с использованием пакета программ Statisti-ca 6.0 (“StatSoft Inc.”, США). На рисунках представлены средние значения (столбцы, символы) и стандартные ошибки (отрезки).
Результаты и их обсуждение
В данной работе проницаемость клеточной стенки бактерий оценивали по скорости транспорта β-лактама цефазолина через внешнюю мембрану бактериальных клеток, подвергнутых воздействию субтерапевтических доз фторхинолоновых антибиотиков (пефлоксацин, левофлоксацин), которые не приводили к быстрой гибели микроорганизмов.
На второй час воздействия фторхинолоновых антибиотиков происходило существенное снижение пориновой проницаемости (рис. 1, 2). Дальнейшая экспозиция с фторхинолоном сопровождалась еще большим ингибированием проницаемости, но темп снижения замедлялся по сравнению с первыми часами воздействия антибиотика.
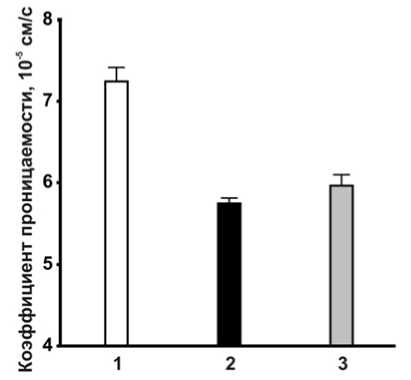
Рис. 1 . Изменение проницаемости клеточной стенки E. coli RO91 на второй час после добавки антибиотиков:
1 – контроль в отсутствие антибиотика, 2 – 0.012 мкг/мл левофлоксацина, 3 – 0.036 мкг/мл пефло-ксацина. * – отсутствие статистически значимых отличий от контроля (t-критерий, р ≤ 0.05)
Воздействие фторхинолонов сопровождалось увеличением внутриклеточного содержания основных полиаминов (рис. 2). Концентрации свобод- ных путресцина и спермидина возрастали в первые 2 ч. воздействия антибиотика, а затем наблюдалось снижение содержания данных полиаминов. В то же время, накопление кадаверина происходило на протяжении всего времени наблюдения и не имело тенденции к снижению по мере увеличения времени экспозиции с антибиотиком. Существенное опережение выхода пиковых концентраций путресцина и спермидина, по сравнению с кадаверином, после внесения антибиотиков может свидетельствовать о существовании функциональной специализации разных полиаминов при адаптивном ответе клетки на действие антибиотиков.
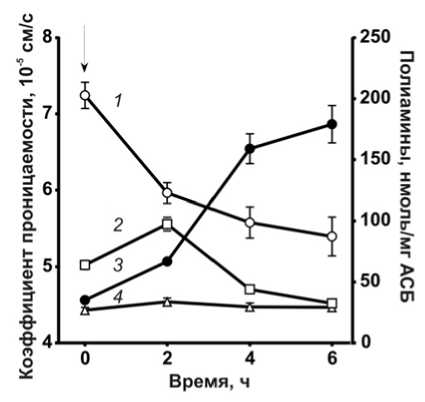
Рис. 2. Изменение проницаемости клеточной стенки (1), содержания путресцина (2), кадаверина (3) и спермидина (4), вызванное воздействием 0.036 мкг/мл пефлоксацина.
Стрелкой обозначен момент внесения антибиотика
Корреляционный анализ выявил наличие обратной зависимости между пориновой проницаемостью клеточной стенки и внутриклеточной концентрацией кадаверина (R = ‒0.485, р = 0.03). Подобной корреляции между проницаемостью и содержанием других полиаминов выявлено не было.
Интенсивность потока ксенобиотиков в клетку определяется не только транспортной активностью (проницаемостью) пориновых каналов, но и их количеством. Известно, что транспорт антибиотиков осуществляется, преимущественно, через белковые каналы OmpF, количество которых в клетке находится под строгим контролем. Один из механизмов, отрицательно регулирующих содержание OmpF на уровне синтеза белка, основан на взаимодействии антисмысловой РНК micF с мРНК ompF , что блокирует процесс ее трансляции.
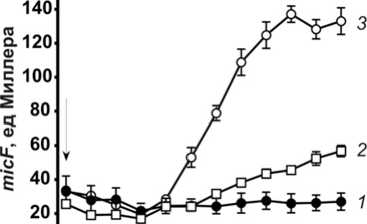
В условиях воздействия фторхинолоновых антибиотиков уровень экспрессии micF значительно возрастал (рис. 3). Стоит отметить, что активация гена micF наблюдалась после второго часа воздействия антибиотика, в то время как пориновая про- ницаемость к этому времени была уже значительно снижена. Можно предположить, что на первых этапах воздействия антибиотиков снижение проницаемости клеточной стенки происходит за счет блокирования просвета канала, а далее подключаются механизмы регуляции количества поринов.
о------------------------------------ 0 1 2 3 4 5 6
Время, ч
Рис. 3 . Изменение экспрессии гена micF в клетках E. coli N9212 в ответ на воздействие антибиотиков:
1 – контроль в отсутствие антибиотика, 2 – 0.012 мкг/мл левофлоксацина, 3 – 0.042 мкг/мл пефло-ксацина. Стрелкой обозначен момент внесения антибиотиков
Момент активации экспрессии micF совпадал с максимальным уровнем накопления путресцина и спермидина. Известно, что физиологические функции и зоны локализации полиаминов в микробной клетке различаются. Кадаверин, по большей части, локализуется в районе клеточной оболочки, тогда как путресцин и спермидин обнаруживаются в цитоплазме, как в свободной форме, так и связанными с нуклеиновыми кислотами. Специфическая локализация и большее сродство к нуклеиновым кислотам определяют преимущественное участие путресцина и спермидина, а не кадаверина в регуляции генной экспрессии [Igarashi, Kashiwagi, 2018]. Поскольку известно, что micF находится под положительным контролем полиаминов [Tkachenko, Pozhidaeva, Shumkov, 2006], обнаруженная нами индукция данного гена в ответ на действие антибиотиков (рис. 3) может быть обусловлена, в том числе, накоплением данных поликатионов в клетке. Кроме того, известно, что полиамины, в частности путресцин, положительно модулируют экспрессию транскрипционного регулятора RpoS [Tkachenko, Shumkov, 2004], под отрицательным контролем которого находится экспрессия гена ompF [Liu, Ferenci, 2001].
Таким образом, полиамины задействованы в снижении проницаемости клеточной стенки E. coli для антибиотиков. Этот эффект может быть обусловлен, по меньшей мере, тремя различными механизмами: прямым отрицательным действием на транспортную активность пориновых каналов, ограничением синтеза пориновых белков на уровне транскрипции, опосредованным RpoS, и снижением экспрессии ompF на уровне трансляции посредством положительной модуляции экспрессии micF.
Работа выполнена в рамках государственного задания, номер госрегистрации темы: АААА-А19-119112290009-1.
Список литературы Роль полиаминов в ограничении проницаемости внешней мембраны Escherichia coli для антибиотиков
- Чудинов А.А., Чудинова Л.А., Коробов В.П. Метод определения низкомолекулярных олигоаминов в различном биологическом материале // Вопросы медицинской химии. 1984. T. 30, № 4. C. 127-132.
- Шумков М.С. и др. Изменение экспрессии ldcC Escherichia coli как фактор адаптации к антибиотикам // Вестник Пермского университета. Сер. Биология. 2010. Вып. 1. С. 36-40.
- Achouak W., Heulin T., Pages J. Multiple facets of bacterial porins // FEMS Microbiology Letters. 2001. Vol. 199, № 1. P. 1-7.
- delaVega A., Delcour A. Polyamines decrease Escherichia coli outer membrane permeability // Journal of Bacteriology. 1996. Vol. 178. P. 3715-3721.
- Delcour A.H. Outer membrane permeability and antibiotic resistance // Biochimica et Biophysica Acta. 2009. Vol. 1794, № 5. P. 808-816.
- Delihas N., Forst S. MicF: an antisense RNA gene involved in response of Escherichia coli to global stress factors // Journal of Molecular Biology. 2001. Vol. 313, № 1. P. 1-12.
- Igarashi K., Kashiwagi K. Effects of polyamines on protein synthesis and growth of Escherichia coli // Journal of Biological Chemistry. 2018. Vol. 293, № 48. P. 18702-18709.
- Iyer R., Delcour A. Complex inhibition of ompF and ompC bacterial porins by polyamines // Journal of Biological Chemistry. 1997. Vol. 272, № 30. P. 18595-18601.
- Liu X., Ferenci T. An analysis of multifactorial influences on the transcriptional control of ompF and ompC porin expression under nutrient limitation // Microbiology. 2001. Vol. 147, № 11. P. 29812989.
- Martin R., Gillette W., Rosner J. Promoter discrimination by the related transcriptional activators MarA and SoxS: differential regulation by differential binding // Molecular Microbiology. 2000. Vol. 35, № 3. P. 623-634.
- Miller J.H. Experiments in molecular genetics. N.Y.: Cold Spring Harbor Laboratory, 1992. 466 p.
- Muffler A. et al. Heat shock regulation of sigmaS turnover: a role for DnaK and relationship between stress responses mediated by sigmaS and sigma32 in Escherichia coli // Journal of Bacteriology. 1997. Vol. 179, № 2. P. 445-452.
- Nikaido H. Molecular basis of bacterial outer membrane permeability revisited // Microbiology and Molecular Biology Reviews. 2003. Vol. 67, № 4. P. 593-656.
- Nikaido H., Rosenberg Y.E., Foulds J. Porin channels in Escherichia coli: studies with ß-lactams in intact cells // Journal of Bacteriology. 1983. Vol. 153. P. 232-240.
- Pratt L. et al. From acids to osmZ: multiple factors influence synthesis of the OmpF and OmpC porins in Escherichia coli // Molecular Microbiology. 1996. Vol. 20, № 5. P. 911-917.
- Samartzidou H., Delcour A. Excretion of endogenous cadaverine leads to a decrease in porin-mediated outer membrane permeability // Journal of Bacteriology. 1999. Vol. 181, № 3. P. 791-798.
- Tabor C. W., Tabor H. Polyamines in microorganisms // Microbiological Reviews. 1985. Vol. 49. P. 8199.
- Tkachenko A., Pozhidaeva O., Shumkov M. Role of polyamines in formation of multiple antibiotic resistance of Escherichia coli under stress conditions // Biochemistry (Mosc). 2006. Vol. 71, № 9. P. 1042-1049.
- Tkachenko A., Shumkov M. Role of putrescine in regulation of the sigmaS subunit of RNA polymerase in Escherichia coli cells on transition to stationary phase // Biochemistry (Mosc). 2004. Vol. 69, № 8. P. 876-882.
