Роль слабосвязанных с клеточной стенкой пероксидаз в устойчивости картофеля при инфицировании кольцевой гнилью

Автор: Граскова И.А., Эпова К.Ю., Кузнецова Е.В., Колесниченко А.В., Войников В.К.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 1 т.4, 2008 года.
Бесплатный доступ
Выявлена роль слабосвязанной с клеточной стенкой пероксидазы в определение устойчивости картофеля к действию патогена. Показано, что добавление к суспензионным клеткам картофеля устойчивого сорта Луговской кроличьих антител к пероксидазе хрена приводило к блокировке активности слабо-связанных с клеточной стенкой пероксидаз и в результате клетки погибали при культивировании с патогеном. Полученные данные позволяют предположить, что устойчивость клеток картофеля на заражение бактериальным патогеном зависит от активности слабосвязанных с клеточной стенкой пероксидаз.
Слабосвязанные с клеточной стенкой пероксидазы, картофель, устойчивость
Короткий адрес: https://sciup.org/14323463
IDR: 14323463
Weak-associated with cell wall peroxidase during the root infection
The role of weak-associated potato cell wall peroxidase in potato resistance to ring rot pathogen is shown. The data obtained showed that an addition of anti-horseradish peroxidase antibodies to resistant to this pathogen potato cv Lugovskoy cell culture caused block of weak-associated potato cell wall peroxidase activity and cell death after addition of pathogen. These data allows us to suppose that the resistance of potato to ring rot pathogen significantly depends on activity of weak-associated potato cell wall peroxidase.
Текст научной статьи Роль слабосвязанных с клеточной стенкой пероксидаз в устойчивости картофеля при инфицировании кольцевой гнилью
В современной биологии наиболее важной проблемой является изучение механизмов иммунитета растений, в частности при действии различных патогенов. Развитие заболевания и его последствия зависят от состояния растения-хозяина и активности его защитных реакций. Среди ферментов, участвующих в защитных ответах растения на атаку патогенов, важную роль играют пероксидазы (Андреева, 1988). Этот фермент реагирует на стрессовые ситуации повышением активности и изменением количества молекулярных форм (Газарян и др, 2006). Существуют пероксидазы, которые связаны ионными или ковалентными связями с полимерами клеточной стенки растения (Минибаева, Гордон, 2003). В последние годы появились сообщения о роли пероксидаз клеточной стенки при патогенезе, где они ответственны за продукцию перекиси водорода в ходе окислительного взрыва в ответ на действие элиситора, продуцируемого патогеном (Bolwell et all, 1999). Было показано, что на поверхности клеток растений существуют пероксидазы, которые способны легко отделяться от клеточной стенки и циркулировать по апопласту целого растения, инициируя в месте контакта с патогеном «иммунный ответ» (Минибаева, Гордон, 2003).
Ранее при изучении динамики активности слабосвязанных с клеточной стенкой пероксидаз картофеля при патогенезе Clavibacter michiganensis subsp . sepedonicus было установлено, что активность исследуемых пероксидаз у устойчивого сорта картофеля выше, чем у восприимчивого, как в контроле, так и после инфицирования (Граскова и др, 2003). У устойчивого сорта картофеля резкое повышение активности слабосвязанной с клеточной стенкой пероксидазы происходит в первые минуты инфицирования, и это увеличение активности не зависит от синтеза фермента de novo, а обусловлено изменением его кинетических параметров (Граскова и др, 2006).
Предполагается, что слабосвязанные с клеточной стенкой пероксидазы важны в формировании устойчивости картофеля к бактериальным патогену, вызывающему кольцевую гниль. Однако фактов, подтверждающих это предположение пока нет. Поэтому целью настоящей работы явилось выявление роли слабосвязанной с клеточной стенкой пероксидазы в определение устойчивости картофеля к действию патогена. Для достижения этой цели использовали иммунохимические методы.
МАТЕРИАЛЫ И МЕТОДЫ
В работе использовали суспензионные культуры клеток сортов картофеля (Solanum tuberosum L.) Луговской (устойчивый к патогену) и Лукьяновский (восприимчивый к патогену). Для получения суспензионных культур клеток картофеля 2-5 мг каллусной листовой ткани помещали в 100 мл жидкой питательной МС- среды с добавлением гормонов и витаминов (Бутенко и др, 1984) и выращивали при непрерывном встряхивании до получения суспензионных культур клеток.
Для заражения клеток использовали штамм 5369 (вирулентный, агрессивный) Clavibacter michiganensis subsp. sepedonicus (Spieck. et Kotth.) Skapt et Burkh, полученный из НИИ картофельного хозяйства (пос. Коренево, Московская обл.). Заражение проводили суспензией бактерий патогена (титр 2 х 108 кл/мл). Суспензионные клетки картофеля устойчивого сорта Луговской культивировали в течение суток в следующих вариантах: 1 – суспензионные клетки с добавлением стерильной среды культивирования (контроль); 2 – суспензионные клетки с добавлением суспензии бактериальных клеток; 3 – суспензионные клетки с добавлением кроличьих антител к пероксидазе хрена; 4 – суспензионные клетки с добавлением патогена и антител. Через сутки культивирования суспензионные ткани фильтровали и определенную массу клеток заливали 0.5% раствором ТТХ (2,3,5-трифенилтетразолий хлорид) и выдерживали 12 ч в темноте при 260С в статичных условиях. Затем, образовавшийся в клетках картофеля формазон извлекали этанолом и полученный раствор колориметрировали. Экстинцию расчитывали на 1 г сырого веса (Еникеев и др, 1995).
Пероксидазную активность анализировали в суспензионных культурах клеток картофеля. Для экстракции внеклеточных пероксидаз навески по 1 г суспензионных тканей картофеля помещали в шприц, заливали 10 мл холодного цитратно-фосфатного буфера (0.1 М, рН 6.2- устойчивый сорт, рН 5.4 – восприимчивый сорт) и дважды по 1 мин выдерживали при вакуумной фильтрации. В фильтрате определяли пероксидазную активность (Граскова и др, 2003). Для этого измеряли изменение оптической плотности при

Рис. 1. Реакция преципитации в агаре. В среднюю лунку внесен раствор, содержащий слабосвязанные с клеточной стенкой пероксидазы, выделенные из суспензионных клеток картофеля устойчивого сорта Луговской. В другие лунки внесен раствор кроличьих антител к пероксидазе хрена.
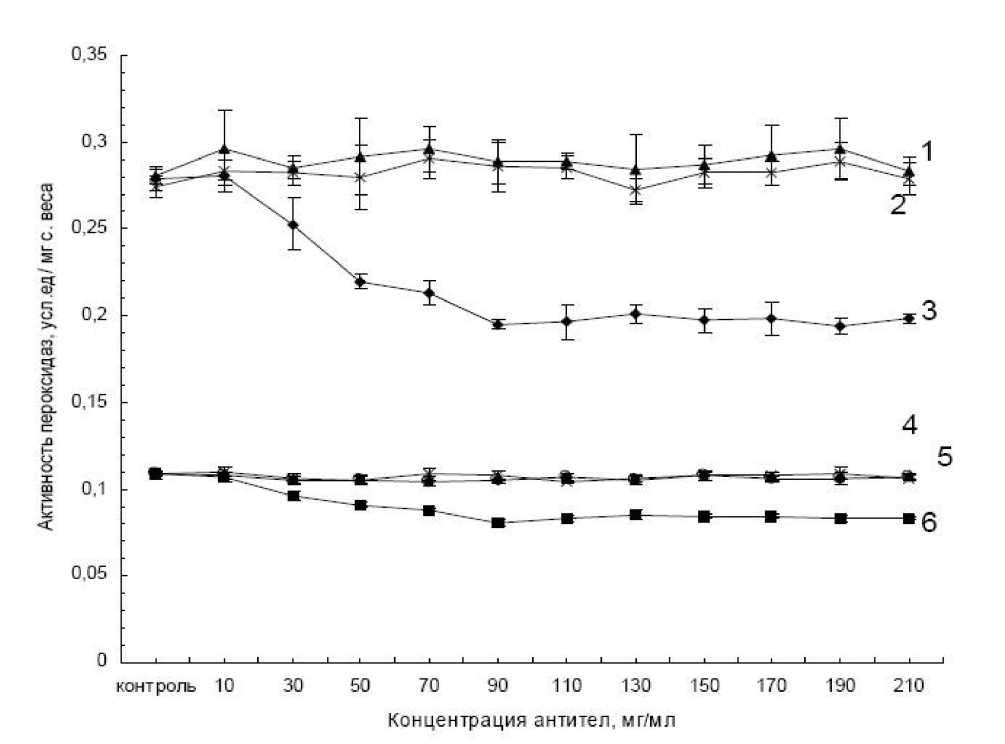
Рис.2. Влияние концентрации кроличьих антител к пероксидазе хрена на активность слабосвязанных с клеточной стенкой пероксидаз, выделенных из суспензионных клеток картофеля сортов Луговской и Лукьяновский. Активность фермента выражена в условных единицах на мг сырой массы.
1 – Луговской (контроль); 2 – Луговской с добавлением кроличьих не иммунных антител к пероксидазе хрена; 3 – Луговской с добавлением кроличьих антител к пероксидазе хрена; 4 – Лукьяновский (контроль); 5 – Лукьяновский с добавлением кроличьих неиммунных антител к пероксидазе хрена; 6 – Лукьяновский с добавлением кроличьих антител к пероксидазе хрена.
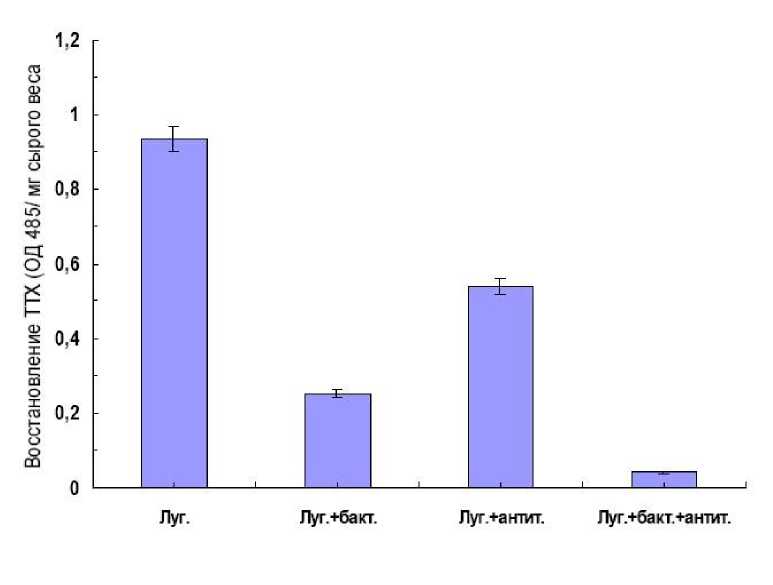
Рис. 3. Выживаемость суспензионных клеток картофеля устойчивого сорта Луговской.
1- жизнеспособность контрольных суспензионных клеток; 2 – клетки с добавлением патогена; 3 – клетки с добавлением антител к пероксидазе хрена; 4 – клетки с добавлением патогена и антител к пероксидазе хрена
580 нм в реакционной смеси из 0.5 мл 0.1 М цитратно-фосфатного буфера (рН 6.2 и 5.4), 0.5 мл 0.3% Н 2 О 2 и 0.5 мл (0.035%, 0.04%, 0.045%, 0.05%,0.055%) гваякола (“Sigma”). Активность пероксидазы определяли при 25оС. Активность фермента рассчитывали по методу Бояркина с модификациями (Граскова и др, 2003). На рисунках указаны средние значения и стандартные ошибки. Каждый вариант опыта проведен в трех-пяти биологических повторностях.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Была проведена иммунизация кроликов препаратом пероксидазы хрена. Полученные антитела проверяли на преципитацию в плоском слое агара, находящегося в чашке Петри. В агаре делали лунки, в одну из которых заливали исследуемые слабосвязанные с клеточной стенкой пероксидазы, выделенные из клеток картофеля устойчивого сорта, а в другие препарат антигена. Антигены и антитела, диффундируя из лунок, встречаются и образуют линии преципитации, которые соответствуют парам антиген-антитело, имеющимся в исследуемой системе (Иммунохимический анализ, 1968). На рисунке 1 наблюдается реакция идентичности: полосы преципитации, образованные антигенами и антисывороткой к ним, сливаются, что указывает на серологическое тождество сравниваемых антигенов.
Для подборки оптимальной концентрации кроличьих антител к пероксидазе хрена варьировали количество антител, выбирая такое их количество, при котором активность слабосвязанной с клеточной стенкой пероксидазы была наименьшей. На рисунке 2 приведены результаты конкурентного анализа кроличьих антител к пероксидазе хрена на активность слабосвязанных с клеточной стенкой пероксидаз, выделенных из суспензионных клеток устойчивого и восприимчивого сортов картофеля: Луговской и Лукьяновский. Активность фермента не изменялась, когда добавляли разные концентрации неиммунной сыворотки. Добавление кроличьих антител к пероксидазе хрена привело к уменьшению активности (рис. 2). Концентрация антител 1.2 мг/ мл наиболее сильно снижала активность исследуемой пероксидазы, выделенной из клеток как устойчивого, так и восприимчивого сортов картофеля. В дальнейших опытах использовали именно эту концентрацию антител.
При изучении роста культур клеток картофеля на воздействие патогена необходимо оценивать их жизнеспособность, которая определяется как соотношение живых и мертвых клеток. Жизнеспособность суспензионных культур можно определить с помощью «прижизненных» красителей с последующим подсчетом окрашенных и неокрашенных клеток (Еникеев и др, 1995, Bordnman et al., 1985). Использовали метод прижизненного окрашивания растительной клетки красителем 2,3,5-трифенилтетразолий хлоридом (ТТХ). Живые клетки способны восстанавливать ТТХ в водонерастворимый формазон, который извлекается органическими растворителями и колориметрируется (Еникеев и др, 1995). Восстанавливающая способность живых клеток существенно зависит от их метаболитической активности, которая резко меняется в ходе культивирования и под влиянием различных стрессовых воздействий. Полученные
Список литературы Роль слабосвязанных с клеточной стенкой пероксидаз в устойчивости картофеля при инфицировании кольцевой гнилью
- Андреева В.А. (1988) Фермент пероксидаза. М.: Наука, 128.
- Бутенко Р.Г., Хромова Л.М., Седнина Г.В. (1984) Методические указания по получению вариантных клеточных линий и растений у разных сортов картофеля. М.: ВАСХНИЛ, 28.
- Газарян И.Г., Хушпульян Д.М., Тишков В.И. (2006) Особенности структуры и механизма действия пероксидаз растений. Успехи биологической химии. 46, 303-322.
- Граскова И.А., Боровский Г.Б., Колесниченко А.В., Войников В.К. (2003) Различия в механизмах изменения активности пероксидазы устойчивого и восприимчивого сортов картофеля при патогенезе кольцевой гнили. Вестник ХНАУ. Серия Биология. 3(2), 70-71.
- Граскова И.А., Антипина И.В., Войников В.К. (2006) Изменение кинетических параметров слабосвязанной с клеточной стенкой пероксидазы суспензионных клеток картофеля в начальный период инфицирования. ДАН. 409(4), 566-569.
- Еникеев А.Г., Высоцкая Е.Ф., Леонова Л.А., Гамбург К.З. (1995) Об использовании 2,3,5-трифенилтетразолий хлорида для оценки жизнеспособности культур растительных клеток. Физиология растений. 42(3), 423-426.
- Иммунохимический анализ (1968). М.: Медицина, 300.
- Минибаева Ф.В., Гордон Л.Х. (2003) Продукция супероксида и активность внеклеточной пероксидазы в растительных тканях при стрессе. Физиология растений. 50, N3, 459-464.
- Bolwell G.P., Blee K.A., Butt V.S., Davies D.R., Gardner S.L., Gerrich C., Minibayeva F., Rowntree E.G., Wojtaszek P. (1999) Recent Advances in Understanding the Origin of the Apoplastic Oxidative Burst in Plant Cells. Free Radical Res. 31, 137-145.
- Bordnman C.H., Bordnman J.F. (1985) Plant Protoplast Viability//The Physiological Properties of Plant Protoplasts/Ed. Pilet P.E. Berlin; Heedelberg: Springer-Verlag, 29.Berlin; Heedelberg: Springer-Verlag, 29.
