Роль света в реакции огурца на дневной кратковременный перепад температуры

Автор: Икконен Е.Н., Шибаева Т.Г., Титов А.Ф.
Журнал: Журнал стресс-физиологии и биохимии @jspb
Статья в выпуске: 2 т.13, 2017 года.
Бесплатный доступ
В производстве парниковых газов концепция снижения температуры стала важным инструментом для контроля удлинения стебля ряда декоративных видов растений, растений для подстилки и растительных пересадок. Эта концепция использует тот факт, что у многих видов удлинение ствола растения чувствительно к короткому суточному перепаду температуры утром или в конце ночи. Здесь была исследована роль света в растениях огурца (Cucumis sativus L.) на суточное падение температуры. Растения выращивали при относительно низкой интенсивности (120 мкмоль м-2 с-1 ППФД) или «высокой» (320 мкмоль м-2 с-1 ППФД) интенсивности света. Изучено влияние 2-часового падения температуры от 23 до 9 ° C либо в конце ночи (в темноте), либо в начале дня (в свете) роста растений, фотосинтеза и дыхательной активности. В экспериментах использовались светостойкие и устойчивые к оттенкам гибриды огурцов. Результаты показали дифференциальную температурную регуляцию морфологии растений и фотосинтетической активности в свете и темноте. Падение температуры в свете привело к более выраженному уменьшению биометрических и фотосинтетических параметров растений, чем падение температуры в темноте. Интенсивность света во время роста растений также изменяла реакцию растений на падение температуры. В частности, падение температуры в ночное время привело к снижению фотосинтетической активности только у растений, выращенных под «низким» светом. Напротив, дыхание листьев усиливалось в результате падения температуры в растениях, выращенных под «высоким» светом. Легкотяжелые и устойчивые к оттенкам гибриды имели в целом аналогичный ответ на падение температуры, но некоторые различия были также Таким образом, степень легкого торможения митохондриального дыхания снижалась из-за падения температуры только у листьев светопотребительных гибридов, а также демонстрировало большее снижение содержания фотосинтетического пигмента в ответ на падение температуры по сравнению с оттеноподобными растениями. Это говорит о том, что реакция растения на суточный кратковременный перепад температур зависит в значительной степени от времени обработки капель (в свете или темноте) и условий роста света. Степень спроса на свет растений также может быть важна.
Короткий адрес: https://sciup.org/14324012
IDR: 14324012
The role of light in cucumber plant response to a diurnal short-term temperature drop
In greenhouse production a temperature drop concept has become an important tool to control stem elongation of a number of ornamental plant species, bedding plants and vegetable transplants. This concept takes advantage of the fact that, in many species, plant stem elongation is sensitive to a short diurnal temperature drop in the morning or in the end of the night. Here, the role of light in cucumber plant ( Cucumis sativus L.) response to a diurnal temperature drop has been investigated. Plants were grown under relatively “low” (120 µmol m-2 s-1 PPFD) or “high” (320 µmol m-2 s-1 PPFD) light intensity. The effect of a 2 h temperature drop from 23° to 9°C, either in the end of the night (in darkness) or in the beginning of the day (in light) on plant growth, photosynthetic and respiratory activity was studied. Light-demanding and shade-tolerant cucumber hybrids were used in experiments. The results showed differential temperature regulation of plant morphology and photosynthetic activity in light and darkness. A temperature drop in the light led to more pronounced decrease in biometric and photosynthetic parameters of plants than a temperature drop in darkness. Light intensity during plant growth also modified plant response to a temperature drop. In particular, a temperature drop in the night led to a decrease in photosynthetic activity only in plants grown under "low" light. The respiration of the leaves, on the contrary, was intensified as a result of a temperature drop in plants grown under “high" light. Light-demanding and shade-tolerant hybrids had generally similar response to a temperature drop, but some differences were also revealed. Thus, the degree of light inhibition of mitochondrial respiration decreased due to a temperature drop only in the leaves of light-demanding hybrids. They also demonstrated greater reduction of photosynthetic pigment content in response to a temperature drop compared to shade-tolerant plants. This suggests that plant response to a diurnal short-term temperature drop depends to large extend on the timing of the drop treatment (in light or darkness) and light growing conditions. The degree of plant light demand may also be important.
Текст научной статьи Роль света в реакции огурца на дневной кратковременный перепад температуры
Сокращения: ДРОП – кратковременные ежесуточные понижения температуры (от англ. drop); СКП – световой компенсационный пункт; ФСА – фотосинтетический аппарат; A n – скорость видимого фотосинтеза; A g – скорость гросс-фотосинтеза; R d –митохондриальное дыхание в темноте; R l – митохондриальное дыхание на свету; α –видимый квантовый выход фотосинтеза.
Температура и свет являются ведущими экологическими факторами, которые в естественных условиях действуют на растения взаимосвязанно. Недостаток или избыток тепла и света вызывают у растений различные отклонения и/или нарушения, которые могут усиливаться за счет взаимного наложения эффектов этих факторов. Учитывая непостоянство свето-температурных условий в течение суток, время, когда растения подвергаются действию низких температур, также играет немаловажную роль. Например, известно о различиях в реакции растений на холод в присутствии света и в темноте в отношении их роста в высоту и удлинения междоузлий (Ueber, Hendriks, 1992; Grimstad, 1995; Grindal, Moe, 1995). Обусловлено это, как предполагается, тем, что экспрессия генов, ответственных за инактивацию гиббереллинов, контролирующих ростовые процессы, происходит только при снижении температуры на свету (Stavang et al., 2007, 2010). Характер и глубина структурно-функциональных изменений фотосинтетического аппарата (ФСА) растений, вызываемых действием холода, особенно у теплолюбивых видов, также в значительной степени зависит от световых условий. Как правило, действие холода на свету приводит к бóльшим функциональным нарушениям ФСА, чем действие этого фактора в темноте (Allen, Ort, 2001). Даже относительно слабый свет при холодовом воздействии может приводить к поглощению растениями не используемой в фотохимических реакциях световой энергии и вызывать фотоингибирование, проявляющееся в снижении скорости фотосинтеза, фотоокислении пигментов и разрушении структуры хлоропластов (Hetherington et al., 1989). Поскольку низкие температуры, прежде всего, снижают скорость темновых фотосинтетических реакций, у освещенных растений в условиях холода нарушается баланс между поглощением энергии и ее использованием. Поэтому, возможно, что на свету теневыносливые растения, имеющие бóльший размер светособирающего комплекса и меньшие скорости транспорта электронов, могут в бóльшей степени, чем светолюбивые быть подвержены фотоингибированию, вызванному понижением температуры. Однако реакция растений на охлаждение может зависеть не только от того, происходит оно на свету или в темноте, но также от световых условий их роста, предшествующих действию холода. Например, было показано, что интенсивность освещения, при которой растут растения, оказывает заметное влияние на температурную зависимость фотосинтеза (Huner et al., 1998), хотя это не характерно для процесса дыхания (Zaragoza-Castells et al., 2007). Подобные различия в реакции процессов фотосинтеза и дыхания на свето-температурные условия роста могут вызывать изменение соотношения этих двух процессов и, соответственно, отражаться на росте и развитии растений.
Следует отметить, что изучению влияния низкой положительной температуры на теплолюбивые растения в различных световых условиях посвящено значительное количество работ, но в подавляющем их большинстве рассматриваются эффекты ее продолжительного действия (сутки и более). Однако в природе нередко возникает ситуация, когда растения испытывают периодически повторяющиеся понижения температуры всего на несколько часов в суточном цикле, чаще в ночные или ранние утренние часы. В практике тепличного растениеводства ежесуточные непродолжительные (на 2-3 ч) снижения температуры на 5-15°С (ДРОП, от англ. drop – падение), обычно в конце ночи или рано утром, нашли применение в качестве агроприема, который широко используется для получения компактной рассады овощных культур, клумбовых и цветочных растений без применения ретардантов. При этом подавляющая часть имеющихся данных, касающихся эффективности ДРОП-воздействий в зависимости от времени суток (ночью - в темноте или днем - на свету) посвящена их влиянию на габитус растений и их биомассу (Myster, Moe, 1995; Moe, Heins, 2000). Сведений о влиянии ДРОП в разных световых условиях на фотосинтетическую и дыхательную активность растений крайне мало. Тем не менее, на примере огурца было показано, что в отличие от продолжительного воздействия низкой температуры, вызывающего необратимые нарушения ФСА (Zhou et al., 2007), ДРОП-воздействия в темноте могут, наоборот, повышать устойчивость фотосинтеза к охлаждению (Ikkonen et al., 2015), однако роль света в реакциях ФСА растений на ДРОП не изучалась.
Исходя из вышесказанного, в задачи данной работы входила оценка влияния (а) освещения растений в период кратковременного действия низкой температуры (ДРОП в темноте и на свету), (б) световых условий роста растений (уровень освещенности) и (в) требовательности растений к свету (светолюбивые, теневыносливые) на ответные реакции Cucumis sativus L. на ДРОП-воздействия.
MATERIALS AND METHODS
Условия выращивания
Эксперимент 1. Влияние световых условий выращивания на реакцию растений на ДРОП, осуществляемый в темноте или на свету.
Растения огурца ( Cucumis sativus L., гибрид Зозуля F1) выращивали в камере искусственного климата (Vötsch, Германия) при поливе питательным раствором (рН 6.2–6.4), температуре воздуха 23°С, фотопериоде 12 ч, влажности воздуха 70% и освещенности ФАР, равной 120 мкмоль/(м2 с) («низкая» освещенность) или 320 мкмоль/(м2 с) («высокая» освещенность). Когда первый лист достигал 60-70% от своей максимальной площади, растения в течение 8 суток подвергали 2-часовому действию температуры 9°C в конце ночного периода (вариант ДРОП ночь ) или в начале дневного периода (вариант ДРОП день ) в камере искусственного климата (Snijders Microclima 1750, Нидерланды), сохраняя прочие условия неизменными. Растения, не подвергающиеся ДРОП-воздействиям, были приняты за контроль.
Эксперимент 2. Сравнительная оценка влияния ДРОП на ФСА и дыхание у гибридов огурца, различающихся по требовательности к свету.
Растения огурца ( Cucumis sativus L., светолюбивые гибриды Кадриль F1, Кураж F1, Хасбулат F1 и теневыносливые Берендей F1,
Гирлянда F1, Рафаэль F1) выращивали в камере искусственного климата (Vötsch, Германия) при поливе питательным раствором (pH 6.2-6.4), температуре воздуха 25/20°С (день/ночь), освещенности 250 мкмоль/(м2 с) ФАР, фотопериоде 16 ч, влажности воздуха 70%. Растения подвергали действию ДРОП ночь или ДРОП день в течение 11 суток в период роста первого настоящего листа.
Анализ морфологических и физиологических показателей. На следующий день после завершения периода, когда растения подвергались ДРОП-воздействиям, анализировали морфологические и физиологические показатели. Определяли сухую биомассу растений и площадь листьев. Рассчитывали отношение сухой массы листьев к их площади (LMA, от leaf mass per area). Содержание хлорофилла (Хл) a и b и каротиноидов (Кар) в эксперименте 1 определяли с помощью спектрофотометра СФ-2000 (Спектр, Россия), экстрагируя их 96% этиловым спиртом, и рассчитывали по известным формулам (Lichtenthaler, Wellburn, 1983). В эксперименте 2 относительное содержание хлорофиллов a и b определяли с помощью измерителя уровня хлорофилла SPAD 502 Plus (Konica Minolta, Osaka, Япония). Флуоресценцию хлорофилла измеряли при помощи анализатора фотосинтеза с импульсно-модулированным освещением MINI-PAM (Walz, Германия). Максимальный квантовый выход фотохимической активности ФС II (Fv/Fm) определяли после 20-минутной темновой адаптации листьев. Эффективный квантовый выход фотохимической активности измеряли после 30-минутной световой адаптации листьев и рассчитывали как φII = ΔF/Fm’ = (Fm’- F)/Fm’. Газообмен листьев анализировали с помощью портативной фотосинтетической системы HCM-1000 (Walz, Германия) при температуре листа 23°С. Видимый фотосинтез (An) листьев измеряли при ФАР, равной 1000, 60, 40 и 20 мкмоль/(м2 с). При каждой интенсивности ФАР величину газообмена определяли после полной его стабилизации. По окончанию измерений газообмена листа на свету растение выдерживали в течение 30 мин в темноте и определяли скорость газообмена при 0 мкмоль/(м2 с) ФАР, принимая данную величину за дыхание листьев в темноте (Rd). Видимый квантовый выход фотосинтеза (α) и световой компенсационный пункт (СКП) вычисляли по начальному линейному отрезку световой кривой фотосинтеза, построенной по значениям скорости фотосинтеза при 60, 40 и 20 мкмоль/(м2 с) ФАР (Garmash, Golovko, 1997). Митохондриальное дыхание листьев на свету (Rl) определено согласно методу Kok (Kok, 1948) экстраполяцией на ось Y (An) прямой линии, аппроксимирующей значения An при 60, 40 и 20 мкмоль/(м2 с) ФАР. Cкорость истинного фотосинтеза (Ag) рассчитывали как сумму An при 1000 мкмоль/(м2 с) ФАР и Rl. Все измерения проводили на первом настоящем листе.
В таблице и на рисунке представлены средние значения по двум независимым опытам (4 и более биологических повторностей в каждом варианте отдельного опыта) и их стандартные ошибки. Достоверность различий между средними значениями определяли на основе дисперсионного анализа (LSD тест) при Р < 0.05 с использованием программного обеспечения Statistica (v. 8.0.550.0, “StatSoft, Inc.”).
RESULTS
Эксперимент 1. Влияние световых условий выращивания на реакцию растений на ДРОП, осуществляемый в темноте или на свету.
Полученные результаты показали, что световые условия роста растений оказали существенное влияние как на ростовые процессы, так и на реакции ФСА. Растения, выросшие в условиях «низкой» освещенности, характеризовались более низкой скоростью накопления биомассы, имели меньшую площадь листьев и удельный вес листьев (LMA) по сравнению с растениями, выросшими при «высокой» освещенности (табл. 1). Содержание Хл, Кар, значения φ II были выше в условиях «низкой» освещенности, а величина A n , СКП и R l - в условиях «высокой» освещенности. Уровень освещенности не влиял только на величину F v / F m и α.
Реакция растений на дневной и ночной ДРОП отличалась по ростовым показателям, состоянию и активности ФСА независимо от того, при каких условиях освещения они выросли (табл. 1). ДРОП вызывал большее снижение общей биомассы, площади листьев растений и LMA в случае его действия на свету, чем в темноте. Так, по сравнению с контролем сухая масса растений была ниже в вариантах ДРОПночь и ДРОПдень, соответственно, на 21 и 40% в условиях «низкой» освещенности и на 19 и 38% в условиях «высокой» освещенности. Площадь листьев в вариантах ДРОПночь и ДРОПдень снижалась, соответственно, на 26 и 39% при «низкой» освещенности и на 13 и 24% при «высокой» освещенности.
Независимо от световых условий роста растений дневной ДРОП в большей степени, чем ночной, инициировал изменения в пигментном комплексе. Общее содержание хлорофиллов в листьях вариантов ДРОП ночь и ДРОП день было ниже, чем в контроле соответственно на 8 и 28 % у растений, выросших при «низкой» освещенности и на 7 и 12 % у растений, выросших при «высокой» освещенности (табл. 1). В то время как ДРОП в темноте не влиял на содержание Кар в листьях, ДРОП на свету вызывал снижение их содержания на 20 и 10 % соответственно при «низкой» освещенности и «высокой» освещенности. При «низкой» освещенности у растений в вариантах ДРОП ночь и ДРОП день соотношение Хл/Кар было ниже, чем в контроле на 12%, а при «высокой» освещенности этот показатель был ниже контрольного только у растений в варианте ДРОП ночь .
Изменения в функциональной активности ФСА были также более существенными при ДРОП-воздействиях на свету. В отличие от ночного, дневной ДРОП вызывал снижение величин F v / F m и φ II , независимо от уровня освещенности, при котором росли растения (табл. 1). Видимый квантовый выход (α), отражающий количество связанного в процессе фотосинтеза СO 2 на один квант падающей на растение световой энергии, в вариантах ДРОП ночь и ДРОП день был ниже контрольных значений, соответственно, на 16 и 20 % при «низкой» освещенности и на 15 и 27 % при «высокой» освещенности. У растений, выросших при «низкой» освещенности, скорость A n понижалась в обоих вариантах ДРОП, но в большей степени в варианте ДРОП день . Растения, выросшие при
«высокой» освещенности, сохраняли высокий уровень фотосинтеза в варианте ДРОПночь, но существенно (более чем на 30%) снижали его в варианте ДРОПдень. Действие ДРОП не отразилось
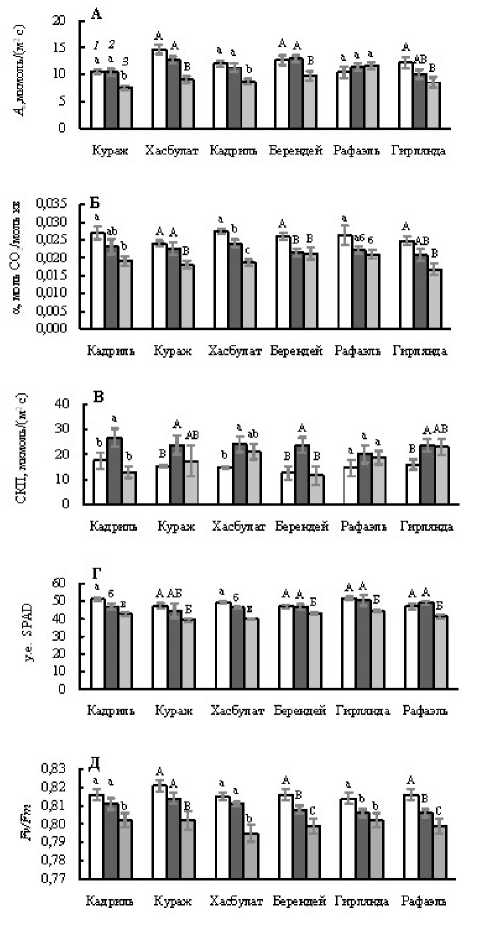
Figure 1. Видимый фотосинтез ( A n , А ) при ФАР, равной 1000 мкмоль/(м2 с), видимый квантовый выход фотосинтеза (α, Б ), световой компенсационный пункт (СКП, В ), относительное содержание хлорофилла (у. е. SPAD, Г ) и максимальный квантовый выход фотохимической активности ФС II ( F v / F m , Д ) у листьев светолюбивых (Кадриль, Кураж, Хасбулат) и теневыносливых (Берендей, Рафаэль, Гирлянда) гибридов огурца, не испытавших (Контроль, 1 ) и испытавших кратковременное ежесуточное понижение температуры до 9°С в темноте (ДРОП ночь , 2 ) и на свету (ДРОП день , 3). Измерения выполнены при температуре листа 23°С. В пределах каждого показателя и каждого гибрида разные буквы указывают на достоверность различий средних значений при Р < 0.05.
на значениях СКП и R l у растений, выросших при «низкой» освещенности, но вызывало их повышение у растений, выросших при «высокой» освещенности, однако, только в варианте ДРОП ночь .
1J0 гА

Кадриль Кураж Хасбупат Беревдей Рафаэль Гирлянда
1Д Б
Кадриль Кураж Хасбупат Берендей Рафаэль Гирлявда
Кадриль Кураж Хаббупат Берендей Рафаэль Гирлянда
Кадр!иль Кураж Хасбупат Берендей Рафаэль Гирлянда
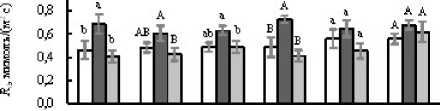
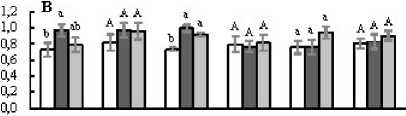
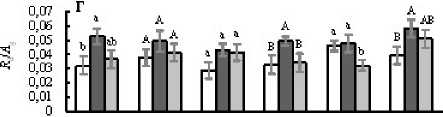
Figure 2. Митохондриальное дыхание листьев на свету ( R , А ) и в темноте ( R d , Б ), отношение R к R d ( R R d , В ), отношение R d к скорости истинного фотосинтеза A g ( R d A g , Г у листьев светолюбивых (Кадриль, Кураж, Хасбулат) и теневыносливых (Берендей, Рафаэль, Гирлянда) гибридов огурца, не испытавших (Контроль, 1 ) и испытавших кратковременное ежесуточное понижение температуры до 9°С в темноте (ДРОП ночь , 2 ) и на свету (ДРОП день , 3). Измерения выполнены при температуре листа 23°С. В пределах каждого показателя и каждого гибрида разные буквы указывают на достоверность различий средних значений при Р < 0.05.
Table 1. Морфо-физиологические показатели растений Cucumis sativus L, выросших в условиях «низкой» (120 мкмоль/(м2 с)) и «высокой» (320 мкмоль/(м2 с)) освещенности и не подвергавшихся (контроль) или подвергавшихся кратковременному (2 ч) ежесуточному воздействию температуры 9°С в темноте (ДРОП««.)
или на свету (ДРОПдень)
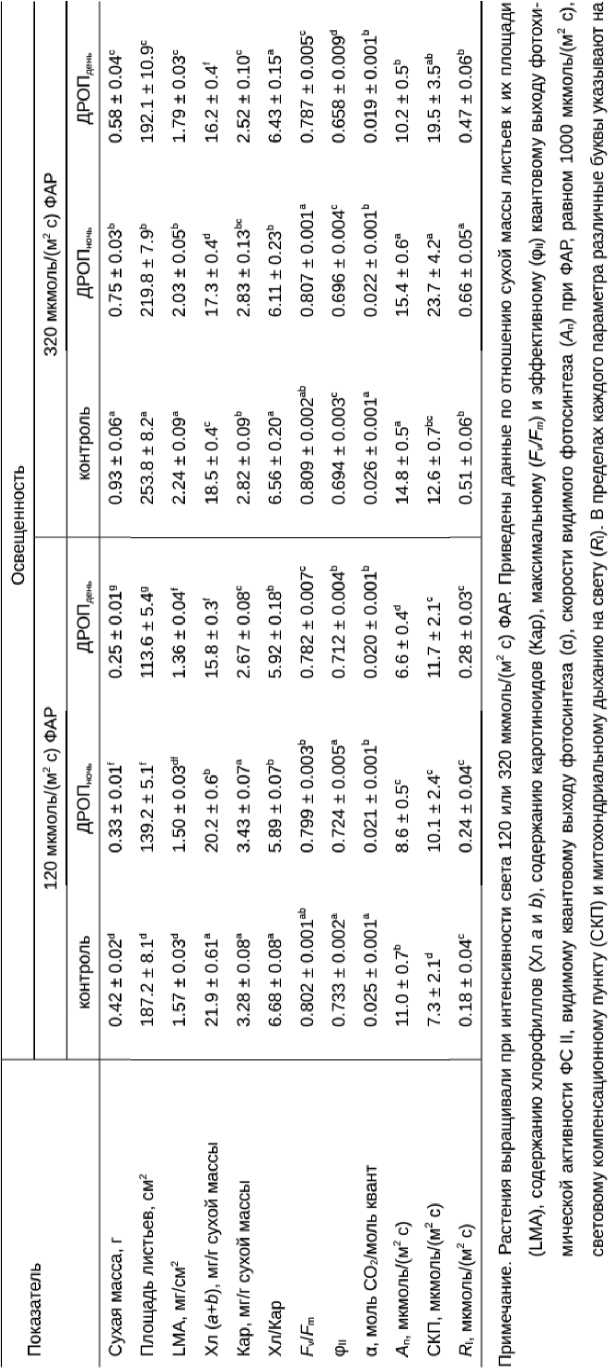
достоверность различий средних значений при уровне значимости Р < 0.05.
Эксперимент 2. Сравнительная оценка ДРОП-воздействий на реакцию гибридов огурца, различающихся по требовательности к свету.
Независимо от отношения гибридов огурца к свету, дневной ДРОП, в отличие от ночного, вызывал снижение скорости A n (рис. 1А). Исключение составил теневыносливый гибрид Рафаэль, оказавшийся малочувствительным к кратковременным понижениям температуры независимо от того, происходили они на свету или в темноте. В вариантах ДРОП ночь и ДРОП день величина α снижалась, соответственно, на 5 и 25% у светолюбивых и на 16 и 23% у теневыносливых гибридов (рис. 1Б). В отличие от дневного, ночной ДРОП вызывал повышение значений СКП у большинства исследованных гибридов независимо от их отношения к свету (рис. 1В).
Наряду с общими чертами в реакции на ДРОП разных по светотребовательности растений огурца, по ряду показателей между светолюбивыми и теневыносливыми гибридами выявлены определенные различия. Так, в варианте ДРОП день содержание Хл снижалось только у теневыносливых гибридов (рис. 1Г). В варианте ДРОП ночь у всех теневыносливых гибридов в отличие от светолюбивых наблюдалось снижение значений F v /F m , а в варианте ДРОП день уменьшение этого показателя происходило как у светолюбивых, так и теневыносливых гибридов (рис. 1Д).
В среднем величины R и Rd листьев светолюбивых растений, не подвергавшихся действию ДРОП, были соответственно на 16 и 12% ниже, чем у теневыносливых (рис. 2А, Б). В варианте ДРОПночь скорости R и Rd светолюбивых растений возрастали в среднем на 73 и 34%, соответственно. Тенденция к подобному усилению дыхания в варианте ДРОПночь прослеживается и у теневыносливых растений, однако достоверно оно только для гибрида Берендей. В варианте ДРОПночь соотношение R Rd не изменялось у теневыносливых и возрастало в среднем на 30% у светолюбивых растений, хотя и не у всех гибридов это повышение было статистически достоверным (рис. 2В). При ДРОП в темноте величина Rd Ag возрастала у всех гибридов независимо от их требовательности к свету, хотя и не у всех гибридов эти изменения подтверждены статистически (рис. 2Г). Также у всех гибридов, за исключением теневыносливого Рафаэля, прослеживалась тенденция к повышению величины Rd Ag при действии ДРОП на свету.
DISCUSSION
Различия в реакциях растений на световые условия роста, зафиксированные в наших опытах, соответствуют существующим представлениям об их адаптации к уровню освещенности (Garmash, Golovko, 1997; Zaragoza-Castells et al. , 2007), хотя в данной работе они не столь ярко выражены. По всей видимости, это связано с тем, что при использованных в работе уровнях ФАР, равных 120 мкмоль/(м2 с) («низкая» освещенность) и 320 мкмоль/ (м2 с) («высокая» освещенность), у растений не возникало недостатка или, наоборот, избытка света. Тем не менее, растения огурца, выросшие при более низкой освещенности, характеризовались меньшими значениями LMA и содержали больше хлорофилла. Однако, как это часто бывает у «теневых» листьев, повышенное содержание хлорофилла не сопровождалось увеличением скорости фотосинтеза, поэтому биомасса растений, выросших в условиях более низкой освещенности, оказалась меньшей. Помимо снижения скорости фотосинтеза, физиологическим откликом растений на более низкую освещенность было снижение скорости митохондриального дыхания и, как следствие, величины СКП, однако они отличались более эффективным использованием света в процессе фотосинтеза. Такая ситуация весьма характерна для листьев растений затененных мест обитания (Garmash, Golovko, 1997; Lewis et al. , 2000; Zaragoza-Castells et al. , 2007). Повышение A n у растений, выросших при более высокой освещенности (табл. 1), могло быть частично обусловлено большим количеством и массой фотосинтезирующих клеток на единицу площади листа, что подтверждается более высокими значениями LMA.
Наряду с этим результаты исследований показали, что эффекты ДРОП на растения могут различаться в зависимости от того, осуществляются
ДРОП-воздействия в присутствии света или в темноте. ДРОП на свету приводил к более выраженному снижению биометрических и фотосинтетических показателей у растений, чем ДРОП в темноте (табл. 1). Меньшая площадь листьев у растений, испытавших действие низких температур на свету, вероятно, связана со снижением активности гиббереллинов (GA1) в результате быстрого увеличения экспрессии одного из генов, деактивирующих гиббереллины PsGA2ox2 (Stavang et al. , 2007). Экспрессия этого гена происходит лишь при действии низкой температуры на свету. Однако, надо отметить, что механизм, связанный с активностью гиббереллинов участвует лишь в изменениях морфологии растений и не влияет на их фотосинтетическую и дыхательную способность (Stavang et al. , 2010). Видимо, снижение скорости фотосинтеза и количества фотосинтетических пигментов в варианте ДРОП день служит своеобразным механизмом «подстройки» ФСА к существующим свето-температурным условиям. Потеря хлорофилла в охлажденных на свету листьях в отличие от листьев, испытавших действие низкой температуры в темноте, может происходить, как предполагается, в результате деградации подверженного фотоингибированию комплекса ФС I (Hetherington et al. , 1989).
Световые условия, в которых выращивались растения, также повлияли на их реакцию на ДРОП по ряду изученных показателей (табл. 1). Так, действие ДРОП в темноте приводило к снижению фотосинтетической активности только у растений, выросших в условиях «низкой» освещенности. Можно предположить, что рост в условиях «низкой» освещенности не способствовал формированию более устойчивого даже к кратковременному охлаждению механизма фиксации СО2. Помимо этого световые условия выращивания растений повлияли на изменения в их дыхательном метаболизме при ДРОП-воздействиях. Усиление митохондриального дыхания в результате темнового ДРОП было характерно только для растений, выросших при «высокой» освещенности. Как известно, адаптация растений к низкой температуре заключается в ряде структурно-функциональных изменений, которые предполагают дополнительный запрос на интермедиаты и энергию, возникающие в процессе дыхания. Не исключено, что адаптационные изменения, проявившиеся в усилении дыхания под влиянием темнового ДРОП, могут реализоваться только в благоприятных для растения световых условиях роста. Повышение скорости дыхания листьев огурца при темновом ДРОП обусловило увеличение соотношения Rd/Ag на уровне листа, что не было характерно для растений, подвергавшихся действию ДРОП на свету (рис. 2Г). Таким образом, результаты данной работы служат дополнительным аргументом в пользу того, что низкая температура на свету и в темноте действует на физиологическое состояние растений по-разному. Возможно, что причина этих различий заключается в разных (или частично разных) «мишенях» воздействия низкой температуры в первом и втором случаях.
Светолюбивые виды растений поддерживают, как правило, более высокие скорости фотосинтеза, чем теневыносливые (Lewis et al., 2000), однако в нашем случае не выявлено достоверных различий по скорости фотосинтеза между светолюбивыми и теневыносливыми гибридами огурца (рис. 1А). Также не обнаружено явных различий в реакции ФСА на действие ДРОП у гибридов, различающихся по требовательности к свету. Обе группы гибридов были способны поддерживать высокую фотосинтетическую активность в условиях кратковременных охлаждений в темноте, но снижали скорость фотосинтеза, если ДРОП-воздействия проводились на свету. Интенсивность митохондриального дыхания на свету обычно ниже, чем в темноте вследствие ингибирования процесса светом, однако, как и дыхание в темноте, оно может адаптироваться к изменениям температуры (Atkin et al., 2000). Под влиянием ДРОПночь дыхание на свету в листьях светолюбивых гибридов огурца усилилось в большей степени, чем дыхание в темноте (рис. 2А, Б). Это означает снижение ингибирования светом дыхания листьев светолюбивых гибридов, что не проявилось у теневыносливых. Также светолюбивые гибриды демонстрировали несколько бóльшую чувствительность пигментного аппарата к действию ДРОП (рис. 1Г).
Эффективность использования света на фотосинтез снижалась под влиянием ДРОП независимо от требовательности растений к свету, от световых условий выращивания и от того, на свету или в темноте подвергались растения действию ДРОП (рис. 1Б, табл. 1). Известно, что видимый квантовый выход (α) влияет на скорость фотосинтеза преимущественно при малой и средней интенсивности света (Garmash, Golovko, 1997). Поскольку рост растений происходит чаще всего при свете, не достигающем насыщающих фотосинтез значений, величина α может определять скорость образования ими первичной продукции (Ehleringer, Björkman, 1977). Наши результаты показали связь величины видимого квантового выхода и накопления растениями сухой биомассы (табл. 1). Возможно, что снижение скорости ростовых процессов у растений огурца под влиянием ДРОП в определенной мере было обусловлено уменьшением эффективности использования ими световой энергии на фотосинтез. Уменьшение величины α возможно вследствие преобразования пигментного комплекса (Garmash, Golovko, 1997), которое, отражая структурнофункциональную реорганизацию ФСА, носит при умеренном стрессе адаптивный характер и является частью процесса повышения его устойчивости к низким температурам (Sofronova et al. , 2014). Видимо, уменьшение пигментного пула под влиянием ДРОП, способствует снижению эффективности использования света ФСА, что можно рассматривать как одну из защитных реакций, позволяющих избежать отрицательного влияния пониженной температуры на свету на клетки листьев растений.
Таким образом, результаты проведенных исследований показали, что световой фактор оказывает существенное влияние на реакцию растений на ДРОП-воздействия. Прежде всего эффекты ДРОП на растения различаются в зависимости от того осуществляются эти воздействия в присутствии света или в темноте. Действие ДРОП на свету вызывает у растений, как правило, более ощутимые изменения в ФСА, приводящие в итоге к снижению скорости накопления биомассы. Негативное влияние ДРОП в присутствии света скорее всего связано со светоиндуцированными нарушениями в ФС II. Однако, ввиду кратковременности (2 часа) действия ДРОП и ежесуточному возврату растений на 22 часа в нормальные температурные условия, происходящие отклонения и/или нарушения носят обратимый характер, о чем свидетельствуют зафиксированные нами значения Fv/Fm (рис. 1Д). Помимо этого важную роль в реакции растений огурца на кратковременные ежесуточные понижения температуры могут играть световые условия роста и светотребовательность растений.
ACKNOWLEDGEMENT
Работа выполнена с использованием оборудования Центра коллективного пользования научным оборудованием Института биологии КарНЦ РАН в рамках государственного задания (№ темы 0221-2014-0032) и частично поддержана РФФИ (проект № 14-04-00840_а).
Список литературы Роль света в реакции огурца на дневной кратковременный перепад температуры
- Allen D.J., Ort D.R. (2001) Impact of chilling temperatures on photosynthesis in warm-climate plants. Trends in Plant Science, 6(1), 36-42
- Atkin O.K., Evanrds E.J., Ball M.C., Lambers H., Pons T.L. (2000) Leaf respiration of snow gum in the light and dark. Interactions between temperature and irradiance. Plant Physiol., 122(3), 915-923
- Ehleringer J., Björkman O. (1977) Quantum yields for CO2 uptake in C3 and C4 plants. Dependence on temperature, CO2 and O2 concentration. Plant Physiol., 59(1), 86-90
- Garmash E. V., Golovko T. K. (1997) CO2 gas-exchange and growth in Rhaponticum carthamoides under the conditions of middle taiga subzone of northeastern Europe. 1. Dependence of photosynthesis and respiration on environmental factors. Russ. J. Plant Physiol., 44(6), 737-745
- Grimstad S.O. (1995) Low-temperature pulse affects growth and development of young cucumber and tomato plants. J. Hortic. Sci., 70(1), 75-80
- Grindal G., Moe R. (1995) Growth rhythm and temperature DROP. Acta Hort., 378, 47-52. Hetherington S.E., He J., Smillie R.M. (1989) Photoinhibition at low temperature in chilling-sensitive and -resistant plants. Plant Physiol., 90, 1609-1615
- Huner N.P.A., Öquist G., Sarhan F. (1998) Energy Balance and Acclimation to Light and Cold. Trends in Plant Science, 3(6), 224-230
- Ikkonen E.N., Shibaeva T.G., Titov A.F. (2015) Response of the Photosynthetic Apparatus in Cucumber Leaves to Daily Short-Term Temperature Drops. Russ. J. Plant Physiol., 62(4), 494-498
- Kok B. (1948) A critical consideration of the quantum yield of Chlorella-photosynthesis. Enzymologia, 13, 1-56
- Lewis J.D., McKane R.B., Tingey D.T., Beedlow P.A. (2000) Vertical gradients in photosynthetic light response within an old-growth Douglas-fir and western hemlock canopy. Tree Physiol., 20(7), 447-456
- Lichtenthaler H.K., Wellburn A.R. (1983) Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans., 603, 591-592
- Moe R., Heins R.D. (2000) Thermo-and photomorphogenesis in plants. In Stroemme, E. (ed.), Adv. Floricult. Res. Report №6. Agric. Univ. of Norway, Aas, Norway, pp. 52-64
- Myster J., Moe R. (1995) Effect of diurnal temperature alternation on plant morphology in some greenhouse crops -a mini review. Sci. Hort., 62(4), 205-215
- Sofronova V.E., Chepalov V.A., Dymova O.V., Golovko T.K. (2014) The role of pigment system of an evergreen dwarf shrub Ephedra monosperma in adaptation to the climate of central Yakutia. Russ. J. Plant Physiol., 61(2), 246-254
- Stavang J.A., Junttila O., Moe R., Olsen J.E. (2007) Differential temperature regulation of GA metabolism in light and darkness in pea. J. Exp. Bot., 58(11), 3061-3069
- Stavang J.A., Pettersen R.I., Wendell M., Solhaug K.A., Junttila O., Moe R., Olsen J.E. (2010) Thermoperiodic growth control by gibberellins does not involve changes in photosynthetic or respiratory capacities in pea. J. Exp. Bot., 61(4), 1015-1029
- Ueber E., Hendriks L. (1992) Intensity effects of a temperature drop on pelargoniums. Acta Hort., 378, 34
- Zaragoza-Castells J., Sanchez-Gomez D., Valladares F., Hurry V., Atkin O.K. (2007) Does growth irradiance affect temperature dependence and thermal acclimation of leaf respiration? Insights from a Mediterranean tree with long-lived leaves. Plant, Cell and Environment, 30(7), 820-833
- Zhou Y., Huang L., Zhang Y., Shi K., Yu J., Nogues S. (2007) Chill-Induced Decrease in Capacity of RuBP Carboxylation and Associated H2O2 Accumulation in Cucumber Leaves Are Alleviated by Grafting onto Figleaf Gourd. Ann. Bot., 100(4), 839-848
