Российский Дальний Восток: эволюция биоты, морфотектоника, новинки флоры

Автор: Урусов В.М., Варченко Л.И.
Журнал: Вестник Красноярского государственного аграрного университета @vestnik-kgau
Рубрика: Биологические науки
Статья в выпуске: 7, 2016 года.
Бесплатный доступ
Цель исследования - выделить главные зо-ны и причины видообразования и эндемизма высокогорий и побережья российского Даль-него Востока и оценить возможности и темп стрессируемой физиологии биоты этих зон перехода глобального уровня. Рассмотрена территория Дальнего Востока (ДВ) и приле-жащих субрегионов. Стрессируемость репро-дукционного процесса, учащение мутаций, особая жѐсткость популяционных волн - в зоне современных и древних, преодолѐнных по-гружением края Азии, высокогорий и на бере-гах существующих и исчезнувших морей. Вы-сокая радиация, химизм воздуха, осадков, верх-него слоя почв обеспечивают адаптивную эволюцию за миллионы и десятки миллионов лет. Эволюция глобального климата в значи-тельной мере определяется динамикой субре-гиональных структур рельефа, в частности гигантских морфоструктур центрального типа (МЦТ). Этап воздымания МЦТ обеспечи-вал изоляцию новообразований в горах мела-олигоцена, где сформированы как характер-ные высоким широтам виды, так и узкие эн-демы-стланцы, древние, но более молодые (Taxusnana, Microbiotadecussata, Sabinadavurica, S. sargentiiи т.д.). На последних этапах разви-тия МЦТ их центры занимают моря, берега которых становятся средой в т.ч. четвер-тичного (квартер) видообразования, включая и эндемизм полиплоидного генезиса. Однако в системе стадиал-межстадиал в квартере ро-тация климата с оборотом 90-110 тыс. лет недосягаема для адаптивной эволюции, заме-няющейся гибридообразованием. В зонах кон-такта глобального уровня и перехода форм уцелевания и организации биоты главным фактором эволюции является стрессирование физиологии генеративных процессов на уровне ценопопуляций видов в особых, а имен-но крайних, периферийных условиях среды. Находки новых эндемов будут приурочены к древним высокогорьям и их береговому пери-метру, новых для России видов - к убежищам от пожаров в южной зоне хвойно-широко-лиственных и темнохвойно-широколиствен-ных лесов.
Тектоника, морфост-руктуры центрального типа (мцт), арктиче-ская мцт 1-го порядка г.и. худякова, мцт 3-5-го порядков, адаптивная эволюция, зоны кон-такта глобального уровня, процент эндемов субальп и берегов, полиплоиды, гибридогенная эволюция системы стадиал-межстадиал, зоны поиска новых эндемов
Короткий адрес: https://sciup.org/14084744
IDR: 14084744 | УДК: 581.524/527:574(571.6)
Russian Far East: evolution of biota, morphotectonics, news flora
Research objective was to allocate the main zones and the reasons of speciation and an en-demism of highlands and the coast of the Russian Far East and to estimate opportunities and rate of stressed physiology of biota of these zones of tran-sition of global level. The territory of the Far East (FE) and adjacent subregions is considered. The Stress ability of reproductive process, increase of mutations, special rigidity of population waves in a zone the modern and ancient, overcome immersion of the region of Asia, highlands and on coast of the existing and disappeared seas. High radiation, chemism of air, rainfall, the top layer of soils pro-vide adaptive evolution for millions and tens of mil-lions of years. Evolution of global climate is consi-derably defined by dynamics of subregional struc-tures of a relief, in particular the huge morfostruktur central type (MCT). The stage of uplift of MCT pro-vided isolation of new growths in mountains of chalk oligocene where both characteristic to high latitudes types are created, and narrow endemy-stlanets, ancient, but younger (Taxusnana, Micro-biotadecussata, Sabinadavurica, S. sargentiiи etc.). At the last stages of development of MCT their cen-ters occupy the seas which coast become the envi-ronment including quarternary (quarter) speciation, including also an endemizm of poliploidny genesis. However, in system stadial and interstadial in quar-ter rotation of climate with a turn of 90-110 thou-sand years is inaccessible to the adaptive evolution which is replaced with hybrid formation. In zones of contact of global level and transition of forms of survival and the organization of a biota the main factor of evolution is stressirovany physiology of generative processes at the level of cenopopulation of types in special, namely extreme, peripheral conditions of the environment. New endem is found, it will be dated for ancient highlands and their coastal perimeter, types, new to Russia, for shelters from the fires in the southern zone of the coniferous and broad-leaved and dark-fur broad-leaved woods.
Текст научной статьи Российский Дальний Восток: эволюция биоты, морфотектоника, новинки флоры
Введение. Рассматривая узловые моменты эволюции флор и ландшафтов Дальнего Востока [23], мы не могли не остановиться на роли влияния зон контакта глобального уровня на макро- и микроэволюционные процессы, зависящие не только от радиационного баланса, мозаики экотопов и общих характеристик макроклимата, химизма почвы и воздуха, напряжённости (стрессируемости) репродукционного процесса, механизмов популяционных волн, репродуктивной и физической изоляции [21] и географии эндемизма, обусловленной прежде все- го положением древних высокогорий, древних и современных берегов морей и особо крупных водоёмов и водотоков, но и глобального эндемизма.
Цель исследования : выделить главные зоны и причины видообразования и эндемизма высокогорий и побережья российского Дальнего Востока и оценить возможности и темп стрессируемой физиологии биоты этих зон перехода глобального уровня.
Материал и методика исследования. Мы по-прежнему уверены в роли, даже ведущей роли разнопорядковых орографических комплексов (морфоструктур центрального типа, или МЦТ) Г.И. Худякова, А.П. Кулакова, С.М. Тащи [25], А.П. Кулакова [10, 11] в результате адаптивной эволюции, хотя бы потому, что они, во-первых, формируют надежнейшую изоляцию новообразованных форм от родительских популяций в процессе воздымания МЦТ в мезозое-кайнозое, во-вторых, обеспечивают надежные пути расселения видов по воздымающимся горным системам или по краю аркто-монтанных пустынь, если идет глобальное похолодание, по мостам суши, пульсирующим как из-за тектонических движений, так и при осыхании шельфа из-за колебания уровня Мирового океана, например, в ритме оледенений и межледниковий, в-третьих, формируют единства эволюционных процессов и их векторов в зонах полярных Арктической и Антарктической МЦТ 1-го порядка, открытых Г.И. Худяковым [24], основоположником геоморфологической школы Тихоокеанского института географии ДВО РАН, и МЦТ 1-2-го порядков Б.В. Ежова и В.Л. Андреева [6], что определяет главные черты и ареал Голарктического царства акад. А.Л. Тахтаджяна [15] и структуру его флористических областей, усложненную макропроцессами эволюции биоты в пределах МЦТ 2-го и 3-го порядков [23]. Мор-фоструктуры центрального типа (МЦТ) 3-го порядка все еще определяют географию по крайней мере восточноазиатских сосен секции Eupit-ys Spach. Их современное разнообразие приурочено к Корейской и, в меньшей степени, Наньлинской, или Восточно-Китайсткой МЦТ. Судя по палеоботаническим метериалам, до похолоданий антропогена эндемичные двухвойные сосны были характерны и МЦТ северной половины Дальнего Востока. Как свидетель- ствуют ареалы микротермных двухвойных сосен родства Pinus sylvestris, обнимающие зону тайги Северного полушария, их происхождение связано с Арктической МЦТ Г.И. Худякова (рис. 1).
На уровне научного предвиденья разработанная нами идея связки развития МЦТ и макромасштаба адаптивной эволюции предвосхищена М.Г. Поповым [12], придававшим первостепенное значение возникновению зональных флор последовательно в высоких широтах со
«спуском» их в низкоширотную зону. Однако М.Г. Попов ошибся в том, что счел арктическую флору наиболее молодой, а флорообразующий эффект МЦТ 3-го порядка, например, Ангарской, Охотской, Алданской, Японской и Корейской, разумеется, не мог и предвидеть.
Морфоструктурный фактор особенно нагляден в формировании ареалов двухвойных сосен, в том числе Дальнего Востока России и в целом Восточной Азии.
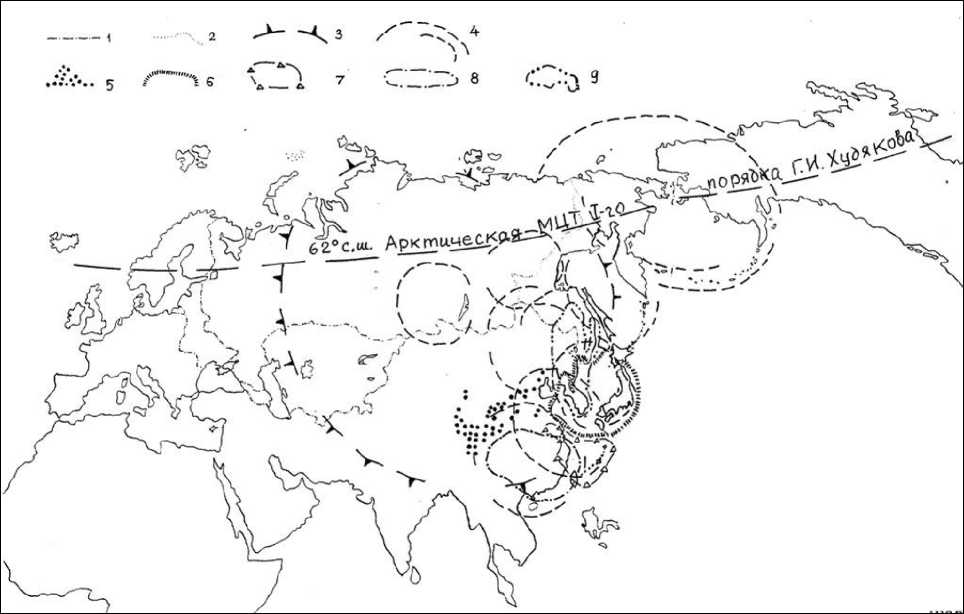
Рис. 1. Ареалы двухвойных сосен, в том числе Дального Востока России и в целом Восточной Азии. Границы: 1 – России и бывшего СССР; 2 – Дальнего Востока; 3 – Азиатской МЦТ 2-го порядка; 4 – МЦТ 3-го порядка (Берингийская, Охотская, Алданская, Ангарская, Амурская, Японская, Корейская, Наньлинская МЦТ); ареалы сосен: 5 – Pinus tabulaeformis; 6 – P. densiflora;
7 – P. luchuensis; 8 – P. massoniana; 9 – P. koraiensis
Более чем тысячекилометровые в диаметре, МЦТ 3-го порядка крайне важны для понимания особенностей распределения высокогорных и береговых эндемов Дальнего Востока (ДВ). Причём первые являются наиболее древними, например меловая (Sabina sargentii), маркирующая контур Японской МЦТ (рис. 2), мелолигоценовая Microbiota decussata, обрамляющая восток Амурской МЦТ, характерная в результате тектонического прогиба для всего периметра Охотского моря Sorbus sambucifolia, происходящая из позднего олигоцена - раннего миоцена. Перед нами ряд высокогорных и подгольцовых эндемов и близких к ним по компактности генерализованных ареалов видов, приуроченных как к верхнему пределу растительности, так и субальпийскому лесному поясу.
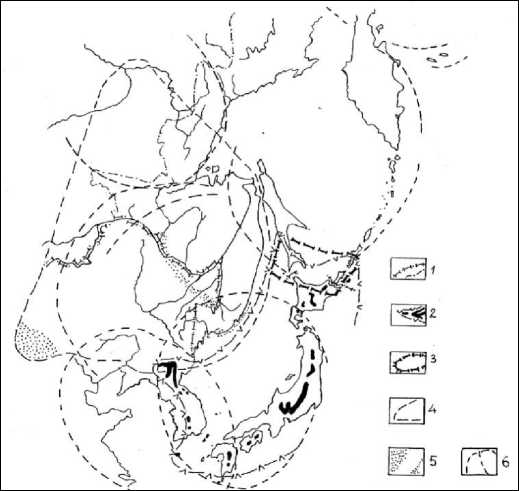
Рис. 2. Видовые ареалы сабиновых можжевельников Дальнего Востока, включая зарубежные страны: 1 – границы России и Дальнего Востока без Якутии; 2 – сабина Саржента (Sabina sargentii) и основные субальпы с её произрастанием; 3 – сниженные – прибрежных пляжей и морских террас – популяции сабины Саржента; 4 – сабина даурская (Sabina davurica); 5 – распространение морского подвида сабины даурской (S. davurica ssp. Maritime) и близких этому подвиду форм в общем ареале S. davurica; 6 – МЦТ 3-го порядка
Амурская МЦТ в силу грандиозной площади образовала, с одной стороны, несколько групп высокогорных эндемов, из которых д.б.н. А.Е. Кожевников [9] особо выделяет Буреинский генцентр и его Амгунь-Буреинскую подгруппу, в границах которых найдены или к которым тяготеют такие эндемичные роды, как Astrocodon, Popoviocodonia, Acelidanthus. Эндемичный сахалинский род Miyakea А.Е. Кожевников счел маркером особой древности области горного сахалинского эндемизма, в которую сместились и молодые эндемичные роды Acelidanthus и Po-poviocodonia .
К Сихотэ-Алинскому генцентру эндемизма мы относим Microbiota decussata, Duschekia mandshurica, Sorbus schneideriana, Lonicera max-imowizcii, Bergenia pacifica и еще целый ряд видов. Однако наиболее знаменитой насельницей данной МЦТ является низко-среднегорная сосна кедровая корейская (Pinus koraiensis), обнимающая линейку достаточно теплых и влажных макроклиматов с суммой активных температур 2000-3000 °C и не подходящая для культуры за пределами области маньчжурского муссонно-континентального климата [20] в отличие от «кореянки» Abies holophylla (Корейская МЦТ) и камчатско-сахалинской A. gracilis, пихте саха линской A. x sachalinensis (= A. mayriana x A. nephrolepis), не тождественной по генетике, морфологии, в т. ч. не типичным для Abies ромбическим листовым следам, микротермности [19] (Охотская МЦТ А.П. Кулакова).
Следующей за приамурским, точнее приамурско-приморским высокогорьем средоточием эндемов являются супралитораль и берега залива Петра Великого (зПв) в ареале эндемичных Heteropappus, Rosa maximowicziana, япономорских эндемов и береговых эндемов Приохотья + береговые эндемы оз. Ханка, ареалы которых отчасти размыты из-за смещений береговой полосы [22]. У Японского моря (береговой периметр) - эндемов и менее компактно расселённых полуэндемов - их ареалы выходят и на берега главных проливов региона. В основном это произошло из-за увеличения плоид-ности, которая весьма подробно изучена Н.С. Пробатовой, В.П. Селедцом, А.П. Соколовской [13], а также этими авторами в содружестве с В.Ю. Баркаловым и Э.Г. Рудыкой [14]). Здесь произрастают молодые подвиды и виды хвойных [19], полиплоидные облигатные виды супралиторали и береговых скал, вошедшие у А.Е. Кожевникова [9] в Япономорскую географи- ческую группу эндемов с подгруппами от Хасан-ской до Сихотэ-Алинско-Сахалинской.
Зоны высокого эндемизма обусловлены положением верхнего предела растительности в миоцене-позднем плиоцене (1) и мелу-олигоцене (2) (рис. 3). Вот что важно: 1) между высокогорьем и берегами эндемы удивительно малочисленны (рис. 4) и сменяются третичными характерными видами с некоторым количеством экстразональных древних сообществ и их реликтовых форм на известняковых скалах [1, 5, 17, 20] и гибридами эпохи активного погружения суши на рубеже плейстоцена вследствие нало- жения поднимающегося хвойно-широколиственного леса на тайгу (таким образом возникли Betula х paraermanii = B. lanata x B. costata [18] и некоторые другие гибриды); 2) временной люфт между эндемами высокогорий и побережья составляет до 30 и более млн лет, что не может не отражать некий штиль в адаптивной эволюции, сменившийся ее ускорением на переходе к квартеру и адаптивным гибридогенезом в плейстоцене-голоцене. Поэтому «пустыня» эндемизма между берегом и высокогорьем вполне понятна (рис. 4).
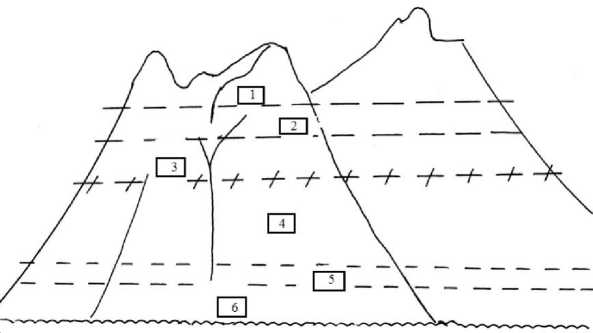
Рис. 3. Зоны высокого эндемизма (1, 2, 3, 6 и отчасти 5) и «пустыня» эндемов – зона 4
Л2& «О ЧАО <50 <60 170 180 КОО0
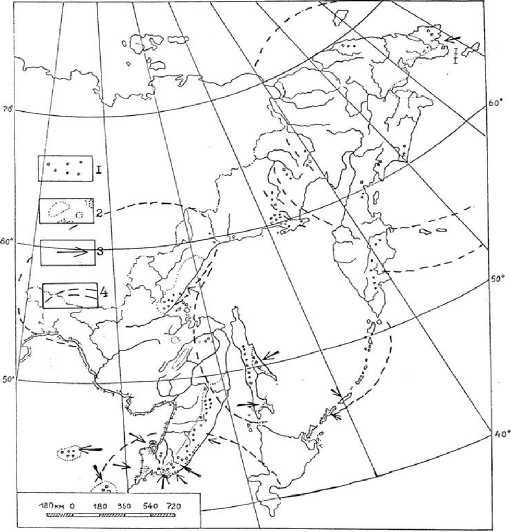
Рис. 4. Основные районы распространения на Дальнем Востоке эндемичных родов (1) и видов (2) сосудистых растений; 3 – зоны, перспективные на открытие новых эндемичных видов;
4 – контуры МЦТ 3-го порядка
Поясним график на рисунке 4, с изображенными на зонами контакта глобального уровня на современном и древнем пределах растительности. Высотно-зональные ландшафты высокого и почти отсутствующего эндемизма в связи с динамикой гигантских МЦТ мы сводим к следующему: 1 - древние эндемы-субальпийцы (в т.ч. вновь открываемые узкие эндемы олигоцено-вого возраста ряда Rhododendron bobrovii Vrishzc [2, 3]; их в ценотипе до 14-16 %); 2 - эн-демы современной границы леса, унаследованные от олигоцен-плиоценовой альпийской тайги (до 10-12 %); 3 - эндемы и полуэндемы поглощенного из-за проседания края Азии микро-термного леса, более полно, чем в Приморье, представленные в корейском секторе ВосточноМаньчжурских гор, включая вулкан Пэктусан (менее 4 % данного ценотипа; эндемизм связан с мощными блоками просевшего купола МЦТ 3-го порядка); 4 - «пустыня» эндемов, эндемизм которой около 1 % за счет таких узкоареальных реликтов, как Actinidia giraldii (это среднегорнонизкогорные полидоминантные леса); 5 - экстраординарные эндемы известняковых скал + местные гибриды (5-6 % и более - от скального ценотипа, здесь продолжают находить новые виды); 6 - эндемы берегов, включая супралито-ральный и каньонный комплексы (до 20 % от общего объема данного флороценотипа).
Итак, в фазе активного воздымания МЦТ 3– 5-го порядков [10] сформированы наиболее древние эндемы как обширных, так и локальных высокогорий. Узкоэндемичные шиповники и другие супралиторальные виды Дальнего Востока, например на берегах зал. Петра Великого (зПВ) Японского моря (в частности Rosa maximowizcii, Oxytropis ruthenicem , Dendranthema erubescens , может быть D. naktongense , D. coreanum , Heteropappus saxomarinus и др.) скорей всего происходят из погрузившейся в море МЦТ 5-го порядка, занимавшей часть суши и акватории зПВ к югу от о-вов Русский и Аскольд (рис. 5).
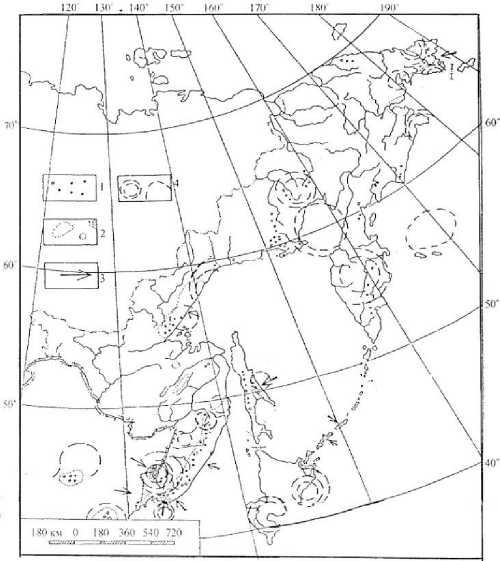
Рис. 5. Основные районы распространения на Дальнем Востоке эндемичных родов (1) и видов (2) сосудистых растений; 3 – зоны, перспективные на открытие новых эндемичных видов;
4 – контуры МЦТ 4–5-го порядков, известных к 1980-м годам
Погружение МЦТ обеспечивает изоляцию экосистем и их популяций на верхнем пределе жизни. Родительские популяции смещаются на более теплые этажи высокогорья или консервируются. Наоборот, на контакте с морями и круп- ными водоёмами, а также на переходе леса в степь или прерию происходит следующее: 1) ускоряется мутагенез, в особенности при подсолении воздуха и почв морскими туманами [7, 8], ускоряется из-за особой физиологии стресси- руемого репродуктивного цикла; 2) становится массовой полиплодия, выраженная почти у 40 % супралиторальных видов [13], что и обеспечило ускоренную макроэволюцию; 3) происходит становление модификаций, которым для перехода в наследственно стойкие подвиды достаточно 300-400 тыс. лет, в виды – 1–2 млн лет; 4) ускоренная, иногда массовая гибридизация, например, у шиповников, можжевельников, мятликов, овсяниц.
Выводы
-
1. В зонах контакта глобального уровня и перехода форм уцелевания и организации биоты главным фактором эволюции является стресси-рование физиологии генеративных процессов на уровне ценопопуляций видов в особых, а именно крайних, периферийных условиях среды.
-
2. Именно поэтому наиболее крупными вкладчиками в эндемизм флоры ДВ являются даже не высокогорья на верхнем пределе растительности, а берега окраинных морей, а это супралиторально-луговой и отчасти лесной и лугово-пойменный комплексы эндемов А.Е. Кожевникова, которые мы бы назвали супралито-рально-степными эндемами, а также аркто-монтанный комплекс эндемов этого же автора и комплекс эндемов крупнотравья. Общий уровень эндемизма – около 6 % (у А.Е. Кожевникова – 10,8 %), эндемов супралиторального комплекса и береговых скал – около 20% от общего объёма данного флороценотипа, аркто-монтанных – примерно столько же (для гольцев Сихотэ-Алиня И.Б. Вышиным выявлен 14 %-й эндемизм), крупнотравного флороценотипа – до 25 % при 10 %-м эндемизме флоры крупнотравных лугов. Следовательно, в занимающих не более, чем первые проценты суши, зонах перехода мутагенез результативней в разы, а мутации случаются на порядки чаще. Добавим к этому наличие свободных для заселения субстратов.
-
3. Если возраст эндемов высокогорий и известняковых скал преимущественно мел-олиго-ценовый, то береговые эндемы чаще четвертичные, не обязательно связаны напрямую с
-
4. Находки новых эндемов будут приурочены к древним высокогорьям и их береговому периметру, новых для России видов – к убежищам от пожаров в южной зоне хвойно-широколиственных и темнохвойно-широколиственных лесов.
МЦТ 5-го порядка и современным расчленением суши.
Список литературы Российский Дальний Восток: эволюция биоты, морфотектоника, новинки флоры
- Верхолат В.П. Флора известняковых обна-жений юга Приморья//Ботанические ис-следования на Дальнем Востоке. -Влади-восток: Изд-во ДВНЦ АН СССР, 1980. -С. 40-54.
- Врищ Д.Л. Предварительная информация о новых таксонах рода Rhododendron L. Си¬хотэ-Алиня (российский Дальний Восток)//Актуальные проблемы ботаники Армении: мат-лы междунар. конф., посвящ. 70-летию Ин-та ботаники. -Ереван, 2008. -С. 76-80.
- Врищ Д.Л., Майоров И.С., Урусов В.М. и др. Экология видов и форм рододендронов Си-хотэ-Алиня//Вестн. ТГЭУ. -2010. -№ 4 (56). -С. 110-124.
- Вышин И.Б. Сосудистые растения высоко-го¬рий Сихотэ-Алиня. -Владивосток: Изд-во ДВО АН СССР, 1990. -116 с.
- Гурзенков Н.Н. Кариологические характери-стики некоторых эндемов флоры Приморья и Приамурья//Комаровские чтения БПИ ДВФ СО АН СССР. -Владивосток, 1969. -Вып. XV-XVII. -С. 73-85.
- Ежов Б.В., Андреев В.Л. Оруденение в мор-фоструктурах центрального типа мантий-ного заложения. -М.: Наука, 1989. -126 с.
- Качур А.Н. Некоторые данные о химиче-ском составе атмосферных осадков При-морской части Среднего Сихотэ-Алиня//Природа и человек. -Владивосток: Изд-во ДВНЦ АН СССР, 1973. -С. 149-154.
- Качур А.Н. Некоторые особенности химиче-ского состава осадков в связи с техногене-зом//Геохимия зоны гипергенеза и техни-ческая деятельность человека. -Владиво-сток: Изд-во ДВНЦ АН СССР, 1976. -С. 28-48.
- Кожевников А.Е. Эндемичный элемент во флоре российского Дальнего Востока//Ко-маровские чтения БПИ ДВО РАН. -Влади-восток, 2007. -Вып. LIV(54). -С. 8-81.
- Кулаков А.П. Морфоструктура востока Азии. -М.: Наука, 1986. -175 с.
- Кулаков А.П. Мегаморфоструктураная эво-люция окраин континента -следствие эво-люции Земли//Структурная организация и взаимодействие упорядоченных социопри-родных систем. -Владивосток: Дальнаука, 1998. -С. 192-203.
- Попов М.Г. Растительный мир Сахалина. -М.: Наука, 1969. -136 с.
- Пробатова Н.С., Селедец В.П., Соколов-ская А.П. Галофильные растения морских побережий советского Дальнего Востока: числа хромосом и экология//Комаровские чтения БПИ ДВНЦ. -Владивосток, 1984. -Вып. XXXI. -С. 89-116.
- Пробатова Н.С., Селедец В.П., Баркалов В.Ю. и др. Основные итоги и перспективы изучения биоразнообразия сосудистых рас¬тений в контактной зоне «материк-океан» (рос-сийский Дальний Восток)//Ритмы и ка-тастрофы в растительном покрове Даль-него Востока. -Владивосток: Изд-во БСИ ДВО РАН, 2005. -С. 112-135.
- Тахтаджян А.Л. Флористические области земли. -Л.: Наука, 1978. -247 с.
- Урусов В.М. Структура разнообразия и про-исхождение флоры и растительности юга Дальнего Востока. -Владивосток: Даль-наука, 1993. -129 с.
- Урусов В.М. География биологического раз-нообразия Дальнего Востока (сосудистые растения). -Владивосток: Дальнаука, 1996. -245 с.
- Урусов В.М. Гибридизация в природной флоре Сибири и Дальнего Востока (при-чины и перспективы использования). -Владивосток: Дальнаука, 2002. -230 с.
- Урусов В.М., Лобанова И.И., Варченко Л.И. Хвойные российского Дальнего Востока. -Владивосток: Дальнаука, 2007. -440 с.
- Урусов В.М., Варченко Л.И., Врищ Д.Л. и др. Владивосток -юг Приморья: вековая и современная динамика растительности. -Владивосток: Дальнаука, 2011. -420 с.
- Урусов В.М., Варченко Л.И. Плоскохвойные ели Северной Пацифики: география, мор-фология, эволюция//Вестн. КрасГАУ. -2011. -№ 8. -С. 88-93.
- Урусов В.М., Варченко Л.И. Об эволюции биоты в береговой зоне дальневосточных морей//Вестн. КрасГАУ. -2013. -№ 11. -С. 187-194.
- Урусов В.М., Врищ Д.Л., Варченко Л.И. Уз-ло¬вые моменты эволюции флор и ланд-шафтов Дальнего Востока в мезозое-кай-нозое//Геогр. вестн. Пермск. гос. ун-та. -2014. -№ 3 (30). -С. 26-37.
- Худяков Г.И. Антиподальные структуры земли и их эволюция//Тихоокеанский еже-годник -1988. -Владивосток: Изд-во ДВО АН СССР, 1988. -С. 85-91.
- Худяков Г.И., Кулаков А.П., Тащи С.М. Но-вые аспекты морфотектоники северо-за-падной части Тихоокеанского подвижного пояса//Геолого-геоморфологические кон-формные комплексы Дальнего Востока. -Владивосток: Изд-во ДВНЦ АН СССР, 1980. -С. 7-24.
