Рост и метаболизм гидробионтов в экспериментальных интегрированных рециркуляционных системах при постоянных и переменных терморежимах

Автор: Зданович В.В., Пушкарь В.Я.
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Зоология
Статья в выпуске: 2, 2008 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/14719000
IDR: 14719000
Текст статьи Рост и метаболизм гидробионтов в экспериментальных интегрированных рециркуляционных системах при постоянных и переменных терморежимах
Стремительное развитие аквакультуры в последние десятилетия обеспечило увеличение объема производимой продукции более чем в два раза и достигло трети от мирового объема добычи гидробионтов. В настоящее время в аквакультуре выращивают более 260 видов рыб, ракообразных, моллюсков, разнообразные виды гидрофитов, при этом в общем объеме культивируемых гидробионтов 51 % составляют рыбы, 27 % гидрофиты, 17 % моллюски и 4 % ракообразные. Если объем промысла гидробионтов в естественных водоемах в последнее время остается практически неизменным, ежегодный прирост получаемой продукции в аквакультуре составляет 7— 10 % [17; 28]. Для культивирования гидробионтов используют как экстенсивные методы, так и методы интенсивного выращивания в садках, бассейнах, замкнутых рециркуляционных системах. Создан ряд высокоэффектив них установок с замкнутым циклом водоснабжения для выращивания разных видов рыб, однако при увеличении плотности посадки продуктивность ограничивается высоким уровнем накопления в воде азотистых продук- тов метаболизма и расхода кислорода, что заставляет включать все более сложные системы очистки и оксигенации воды (20; 21; 25; 31].
При выращивании различных видов рыб только 15—65 % фосфора и 20—50 % азота, содержащихся в кормах, утилизируются рыбами, тогда как остальная часть поступает в воду в растворенном виде и в составе фекалий. В настоящее время при очистке воды, поступающей от рыбоводных ферм, в основном используются физические (механические) способы обработки (фильтрация, седиментация), позволяющие изымать только взвешенные в воде частицы, содержащие 7—32 % общего азота и 30—84 % общего фосфора, находящихся в воде. Растворенные в воде биогенные вещества возможно изъять только с использованием химических или биологических методов очистки [23].
Попыткой решить проблему утилизации вносимого в систему органического вещества было применение в рыбоводных установках поликультуры рыб, а также создание установок, в которых продукты обмена, выделяемые
Работа выполнена при финансовой поддержке РФФИ (грант 07-04-00581)
ВЕСТНИК Мордовского университета | 2008 | № 2
рыбами, используются для выращивания растений [32, 22, 36]. В последнее время интенсивно разрабатываются интегрированные замкнутые рециркуляционные системы, в которых степень утилизации вносимых кормов повышается за счет совместного выращивания организмов разных трофических уровней при условии их пространственного разграничения. В случае выращивании рыбы в таких системах неиспользованные корма и продукты метаболизма утилизируются организмами других трофических уровней (водоросли, ракообразные, моллюски, макрофиты), культивируемых в отдельных блоках [18, 29, 30, 33].
В условиях индустриальной аквакультуры, как правило, используются постоянные температурные условия, прогнозируемые в качестве оптимальных для роста выращиваемых организмов, тогда как в естественных условиях реальны переменные. Результаты работ ряда исследователей по влиянию переменных температур на гидробионтов указывают, что флуктуация температуры в пределах границ экологической валентности вида благоприятна для водных эктотермов. На примере водорослей и макрогидрофитов [3; 4; 15], простейших [7], коловраток [5], ракообразных [6; 19; 37] , моллюсков [15; 34], голотурий [26], рыб [8; 16; 13; 14; 12; 24], земноводных [2] выявлено положительное влияние некоторых колебаний температуры на рост, энергетику, продукционные показатели и физиологическое состояние гидробионтов. Показано, что периодические колебания температуры, не выходящие за верхнюю температурную границу экологической нормы, значительно ускоряют темп роста зоогидробионтов, снижают интенсивность их дыхания, расход кислорода на прирост единицы массы тела, повышают эффективность использования пиши на рост. В условиях осцилляции температуры оптимизируются рост, размножение, энергетика водорослей и высших растений-гидрофитов. Отмечено, что наибольший метаболический эффект у рыб наблюдается в переменных терморежимах при более частых колебаниях температуры с амплитудой 1—3 °C [10].
Цель настоящего исследования — выяснение особенностей роста, энергетики и продукционных показателей гидробионтов разных трофических уровней (продуценты, консументы) при их совместном выращивании в замкнутой рециркуляционной системе в условиях про странственного разграничения при постоянных и переменных терморежимах.
В экспериментальных рециркуляционных установках, каждая из которых состояла из двух или трех блоков, выращивали карпа Cyprinus carpio и брюхоногого моллюска ам-пулярию Ampullaria gigas (опыт 1), а также золотую рыбку Carassius auratus, брюхоногого моллюска физу пузырчатую Physa fontinaiis и смесь сине-зеленых водорослей Oscillatoria spp. и Lyngbya spp. (соотношение по численности — соответственно 90 и 10 %) (опыты 2 и 3) в течение 14—-23 сут. при постоянных и переменных терморежимах. Как модельные объекты карп, золотая рыбка, моллюски и сине-зеленые водоросли выбраны по ряду причин — выдерживают значительное загрязнение воды органическими веществами (мезосапробы), обладают высокой скоростью роста и развития. Кроме того, сине-зеленые водоросли до типу питания мик-сотрофы, т. е. наряду с фотосинтезом активно используют и готовые органические вещества, растворенные в воде.
В первом опыте вода из блока с рыбами (объем 20 л) с помощью аэрлифта подавалась в блок с моллюсками (объем 8 л), а в опытах 2 и 3 затем поступала в блок с водорослями (объем 1,5 л, площадь 460 кв. см) и вновь возвращалась в блок рыб. Скорость протока составляла 3,5 л / ч. В установках с постоянной температурой (20, 23 и 26 °C) необходимый ее уровень поддерживался при помощи терморегуляторов с точностью ± 0,5 °C. В переменном терморежиме 23 ± 3 °C синусоидальные колебания температуры осуществлялись в автоматическом режиме с периодом в 3 ч. Содержание кислорода в воде в блоках с рыбами и моллюсками составляло 80—90 % от полного насыщения (принудительная аэрация). Рыб кормили сухим кормом (Tetra Min XL Flakes; состав — 48 % протеина, 8 % липидов, 2 % углеводов, содержание энергии — 14,21 кДж/г). Рацион составлял 3 % от массы тела рыб в сутки. Рыб кормили два раза в сутки — утром и вечером.
Рыб и моллюсков взвешивали индивидуально в начале и конце каждого опыта. Оценивали среднесуточный прирост, удельную скорость роста по сырой массе по формуле: С^, = (la W2- InW, / t) 100%, где W7, и У72 — средняя масса (г) животных в начале и конце опыта, / — длительность опыта (сут.).
Интенсивность дыхания зоогидробионтов определяли методом прерванного потока {11] при помощи оксиметра с точностью 0,01 мг О2 / л. Определение интенсивности дыхания рыб и моллюсков проводили с 5-кратной повторностью через каждые 7 сут опыта при постоянных и переменных терморежимах. В дальнейших расчетах интенсивность дыхания животных принималась как среднее применительно к тому или иному терморежиму. Оценивали также величину расхода кислорода на прирост 1 г массы тела рыб и моллюсков.
Перед началом опытов в блоки, где выращивали сине-зеленые водоросли, вносили по 0,5 г их сырой массы. Рециркуляционные установки круглосуточно сверху освещали люминис-центными лампами (40 Вт, освещенность 3 000 лк). В конце опытов измеряли массу водорослей при каждом терморежиме, оценивали среднесуточный прирост и удельную скорость роста по сырой массе.
С целью оценки скорости продуцирования рыб, моллюсков и водорослей в рециркуляционных системах при постоянных и переменных температурах рассчитывали суточный Р/В-коэффициент как отношение среднесуточного прироста массы каждого гидробионта к его средней биомассе за время опыта. Статистическая обработка полученных данных осуществлялась с помощью программы «Статистика 5» с использованием критерия Стьюдента.
В табл. 1 представлены результаты выращивания карпа и моллюска ампулярии в рециркуляционных системах при постоянной и переменной температуре. Видно, что все регистрируемые показатели у гидробионтов в переменном терморежиме 23 ± 3 °C значительно отличаются от аналогичных при постоянной температуре 23 °C. У карпа в осциллирующем терморежиме среднесуточный прирост массы тела, удельная скорость роста оказались соответственно в 1,26 и 1,17 раза выше (р < 0,05), тогда как интенсивность дыхания и расход кислорода на прирост единицы массы тела соответственно на 3 и 16 % (р < 0,05) меньше, чем при эквивалентной по сумме тепла постоянной температуре 23 °C. Аналогичные изменения ростовых и метаболических показателей в переменном терморежиме наблюдаются и у ампулярии. Среднесуточный прирост массы тела и скорость роста достоверно возрастают соответственно в 1,45 и 1,43 раза (р < 0,05), а потребление кислорода и его расход на прирост единицы массы тела снижаются соответственно на 5 и 34 % (р < 0,05).
В табл 2 приведены результаты опытов, в которых в интегрированных рециркуляционных системах при постоянных и переменных терморежимах выращивали золотую рыбку, моллюска физу пузырчатую и сине-зеленые водоросли.
В опыте 2 суточный прирост массы тела и удельная скорость роста золотой рыбки при 23 ± 3 °C оказались соответственно в 1,20 и 1,13 раза выше (р < 0,05), чем при 23 °C, тогда
Таблица /
Рост и метаболизм карпа и ампулярии при постоянных и переменных температурах (опыт. /, длительность 15 сут.)
|
Показатель |
Терморежим, °C |
|||
|
Карп (л= 8) |
Ампулярия (л=18) |
|||
|
23 |
23 ± 3 |
23 |
23 ±3 |
|
|
Начальная средняя масса, г |
4,25 ± ± 0,018 |
4,25 ± ±0,019 |
0,672 ± ± 0,002 |
0,667 ± 0,0023 |
|
Конечная средняя масса, г |
9,0 ± ±0,14 |
10,2 ± ± 0,065* |
0,778 ± ±0,001 |
0,822 ± ±0,0005* |
|
Прирост массы тела, г/сут |
0,316 |
0.397 |
0,0071 |
0,0103 |
|
Суммарный прирост массы за время опыта, г |
38,00 |
47,60 |
1,908 |
2,790 |
|
Удельная скорость роста, г %/суг. |
5,00 |
5,84 |
0,97 |
1,39 |
|
Интенсивность дыхания, мг О2/г ч |
0,840 ± ± 0,001 |
0,816± ±0,001* |
0,149± ±0,0004 |
0,141± ± 0,0009* |
|
Потреблено кислорода за опыт, г |
16,03 |
16,98 |
0,70 |
0,680 |
|
Расход кислорода на прирост 1 г массы, г |
0,423 |
0,356 |
0,366 |
" 0,24 3 |
Примечание: * р < 0,05
как интенсивность дыхания и расход кислорода на прирост единицы массы тела — ниже на 4 и 13 % соответственно (р < 0,05). У моллюска физы пузырчатой в переменном терморежиме суточный прирост массы тела и скорость роста по массе превышали наблюдаемые при / 23 °C соответственно в 1,31 и 1,20 раза (р < 0,05), а у сине-зеленых водорослей -— соответственно в 1,12 и 1,03 раза.
В 3 опыте регистрируемые ростовые и метаболические показатели гидробионтов в переменном терморежиме 23 ± 3 °C сравнивались с наблюдаемыми при постоянных температурах, равных нижнему (20 °C), среднему (23 °C) и верхнему (26 °C) уровням температуры при ее осцилляции. У золотой рыбки среднесуточный прирост массы тела при / 23±3 °C был выше соответственно в 2,0,
1,33 и 1,23 раза (р < 0,05— 0,01), чем при 20, 23 и 26 °C. Скорость роста рыб в переменном терморежиме превышала в 1,85 и 1,04 раза, регистрируемую при / 20 и 23 °C и была практически равной в 26 °C. Интенсивность дыхания золотой рыбки и расход кислорода на построение единицы массы тела в переменном терморежиме снижались соответственно на 11, 14 и 20 % (р < 0,01) и 51, 14 и 14 % (р < 0,01) по сравнению с постоянными температурами 20, 23 и 26 °C. Среднесуточные прирост и скорость роста по массе у моллюска физы пузырчатой при / 23 ± 3 °C превышали соответственно в 2,08, 1,35, 1,42 и 1,78, 1,25, 1,22 раза по сравнению с 20, 23 и 26 °C (р < 0,05—0,01). Прирост массы тела и удельная скорость роста сине-зеленых водорослей в условиях осцилляции температуры
Таблица. 2
Рост и метаболизм золотой рыбки, моллюска физы и сине-зеленых водорослей при постоянных и переменных температурах
При совокупности данных, представленных в таблицах, среднесуточный прирост массы тела и скорость роста карпа и золотой рыбки при t 23 ± 3 °C в среднем превышали аналогичные при / 23 °C соответственно в 1,26 и 1,11 раза. Интенсивность дыхания рыб и расход кислорода на прирост единицы массы их тела в переменном терморежиме в среднем ниже соответственно на 3,7 и 14,3 %. У мол люсков суточный прирост массы тела и скорость роста при t 23 ± 3 °C в среднем соответственно в 1,37 и 1,29 раза больше, чем при 23 °C а у сине-зеленых водорослей соответственно в 1,43 и 1,12 раза.
Известно, что скорость продуцирования значительно возрастает с повышением экологической обеспеченности роста особей, с улучшением условий существования гидробионтов, в первую очередь температурных, трофических. На рис. представлены данные о величинах суточных Р/В-коэффициентов рыб, моллюсков и водорослей при разных температурных условиях. Видно, что скорость продуцирования гидробионтов повышается с увеличением температуры от 20 до 26 °C. Однако в условиях переменного терморежима 23 ± 3 °C она неизменно выше, чем при 23 °C, равной средней температуре за цикл, и выше или практически равна с наблюдаемой при 26 °C. Таким образом, при одинаковой сумме тепла и даже меньшей, получаемой гидробионтами при переменном терморежиме, скорости продуцирования выше, чем наблюдаемые при постоянных температурах. Полученные данные еще раз подтверждают, чтб астатичные температурные условия, к которым эволюционно адаптированы гидробионты, являются экологической нормой, тогда как статичность — уклонением от нее, вызывающим некоторое ухудшение энергетики организмов. Сопряженность изменения различных параметров метаболизма, физиологического состояния, темпа роста, продукционных показателей гидробионтов, вызываемых колебаниями температуры, служит тому подтверждением [9].
Интегрированные рециркуляционные аква-культуральные системы с пространственным разграничением биокомпонентов (рыбы — моллюски — водоросли/макрофиты, рыбы — ракообразные — водоросли/макрофиты, рыбы — водоросли / макрофиты) позволяют в большей степени использовать питательные вещества корма на продукционные процессы, уменьшать потребление воды и сброс биогенных элементов в окружающую среду, получать больше продукции. По расчетам, с площади в 1 га интегрированного бассейнового хозяйства с рециркуляцией воды возможно ежегодно получать 25 т рыбы, 50 т двустворчатых моллюсков и 30 т сырой массы водорослей [29]. В экспериментальной морской рециркуляционной установке площадью 3,3 кв.м, где в отдельных блоках выращивали золотистого спара Sparus aurata, брюхоного моллюска морское ушко Н aliotts discus hannai и в качестве биофильтра использовали макроводоросль Ulya lactuca, вода от рыб поступала к моллюскам, затем в блок с водорослями и вновь возвращалась к рыбам среднегодовая продукция составляла соответственно 28, 9,4 и 78 кг / кв.м. При прохождении воды от блоков с рыбами и моллюсками через блок с водорослями со- дрржанир аммонийного азота в нодр снижа лось с 45 до 10 % [33]. В интегрированной рециркуляционной системе, где в трех бассейнах объемом 70 куб. м каждый выращивали трепанга Apostichopus japonicus и в одном — водоросль UIva pertusa, при полном водообмене один раз в сутки из поступающей от трепангов воды водоросли изымали 68 % общего аммонийного азота и 26 % ортофосфата, при этом скорость их роста составляла 3,3 % / сут. [38].
В последние годы возрос интерес к культивированию в рециркуляционных системах сине-зеленых водорослей, в частности спиру-лины Spirulina platensis, которая в благоприятных условиях намного быстрее, чем эука-риотные водоросли, накапливают биомассу, при этом эффективно изымает растворенные органические вещества из воды, обогащает воду кислородом и в дальнейшем используется как корм для выращиваемых рыб [27; 35].
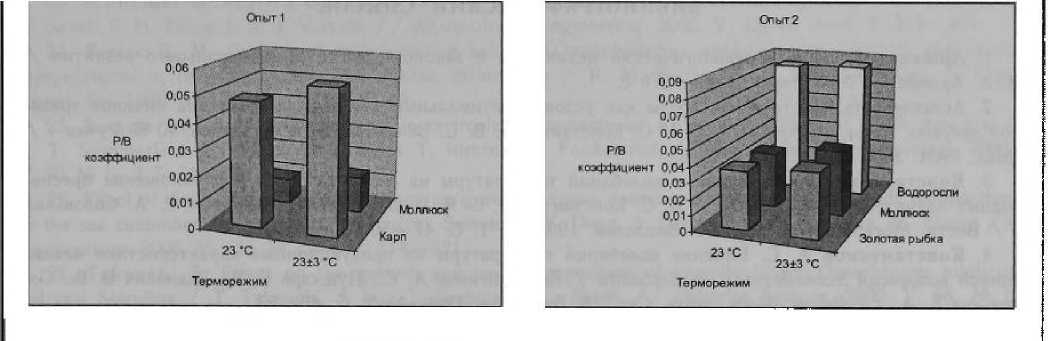
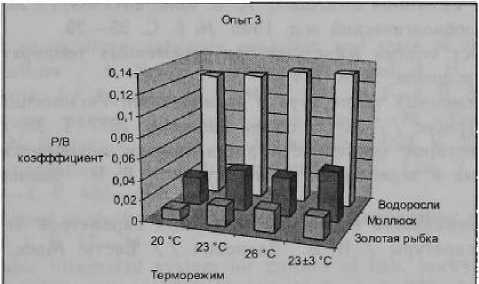
Рисунок.
Суточный Р/В коэффициент рыб, моллюсков и сине-зеленых водорослей при постоянных и переменных терморежимах
Таким образом, проведенные исследования показывают, что в условиях осцилляции температуры в пределах экологической нормы наблюдается усиление темпа роста гидробионтов, оптимизируется энергетика, улучшаются продукционные показатели по сравнению с постоянными температурными условиями. У рыб и моллюсков в переменных терморежимах достоверно снижаются интенсивность дыхания и расход кислорода на прирост единицы массы тела. Величина суточного Р/В-коэффициента рыб, моллюсков и сине-зеленых водорослей при колебаниях температуры заметно . превышает таковые в условиях постоянных температур. В колебательных терморежимах оптимизируется жизнедеятельность различных групп гидробионтов, что указывает на общебиологическую закономерность — необходимость непрерывного нарушения го меостаза с последующим восстановлением за счет работы соответствующих адаптивных механизмов. В ответ на непродолжительные стрессоры слабой и средней силы возникает физиологический стресс (эустресс), сопровождающийся увеличением интенсивности метаболических процессов, что и стимулирует все жизненные процессы [1].
Разработка и внедрение новых перспективных биотехнологий индустриального выращивания гидробионтов, таких как интегрированные рециркуляционные системы для совместного выращивания организмов разных трофических уровней с использованием переменных терморежимов, позволит в значительной мере повысить продуктивность, эффективность использования кормов, уменьшить загрязнение окружающей среды и снизить затраты на производство.
Список литературы Рост и метаболизм гидробионтов в экспериментальных интегрированных рециркуляционных системах при постоянных и переменных терморежимах
- Аршавский И. А. Физиологические механизмы и закономерности индивидуального развития/И. А. Аршавский М.: Наука, 1982. 276 с.
- Астатичность абиотической среды как условия оптимальные для роста и развития личинок травяной лягушки Rana temporaria L/А. С. Константинов, В. С. Вечканов, В. А. Кузнецов, А. Б. Ручин//Докл. РАН. 2000. Т. 371, № 4. С. 559.
- Константинов А. С. Влияние колебаний температуры на скорость роста и размножение пресноводных планктонных водорослей/А. С. Константинов, В. Я. Пушкарь, В. В. Зданович, Е. А. Соловьева//Вестн. Москов. ун-та. Сер. 16. Биология. 1998. № 1. С. 47-50.
- Константинов А. С. Влияние колебаний температуры на продукционные характеристики планктонной водоросли Scenedesmus quadricauda/Константинов А. С., Пушкарь В. Я., Зданович В. В., Соловьева Е. А.//Вестн. Моск. ун-та. Сер. 16. Биология. 1999. № 2. С. 49-53.
- Константинов А. С. Влияние колебаний некоторых абиотических факторов на рост, размножение и энергетику коловратки Euchlanis dilataia/А. С. Константинов, Н. А. Тагирова, В. М. Степаненко, Е. А. Соловьева//Гидробиолгический ж-л. 1995. № 6. С. 25-29.
- Галковская Г. А. Рост водных животных при переменных температурах/Г. А. Галковская, Л. М. Сущеня. Минск: Высш.школа, 1978. 144 с.
- Заар Л. П. Роль переменных температур в размножении Paramecium caudatum/Л. П. Заар, В. А. Тополовский, Ж. М. Трибис//Журн. общей биологии. 1977. Т. 38, № 4. С. 609-619.
- Зданович В. В. Некоторые особенности роста молоди мозамбикской тиляпии Oreochromis mossambicus при постоянных и переменных температурах/В. В. Зданович//Вопр. ихтиологии. 1999. Т. 39, № 1.С. 105-110.
- Зданович В. В. Сопряженность изменения различных параметров метаболизма карпа Cyprinus carpio при колебаниях температуры/В. В. Зданович//Вестн. Моск. ун-та. Сер. 16. Биология. 2004. 3. С. 45-49.
- Зданович В. В. Влияние частых периодических колебаний температуры на метаболизм рыб/B. В. Зданович, В. Я. Пушкарь//Вопр. ихтиологии. 2001. Т. 41, № 3. С.429-432.
- Кляшторин Л. Б. Установка для автоматического измерения дыхания рыб и других гидробион-тов при заданных температурных и кислородных условиях/Л. Б. Кляшторин, Р. Ф. Саликзянов//Вопр. ихтиологии. 1979. Т. 19, № 1. С. 558-561.
- Константинов А. С. Астатичность температурных условий как фактор оптимизации роста, энергетики и физиологического состояния рыб/А. С. Константинов, В. В. Зданович, А. М. Шолохов//Вестн. МГУ. Сер. 16. Биология. 1991. № 2. С. 38-44.
- Константинов А. С. Влияние переменной температуры на рост эвритермных и стенотермных рыб/А. С. Константинов, В. В. Зданович, А. А. Калашников//Вопр. ихтиологии, 1987. Т. 27, вып. 6. C. 971-977.
- Константинов А. С. Влияние осцилляции температуры на интенсивность обмена и энергетику молоди рыб/А. С. Константинов, В. В. Зданович, Д. Г. Тихомиров//Вопр. ихтиологии. 1989. Т. 29, Вып. 6. С. 1019-1027.
- Константинов А. С. Влияние колебаний различных абиотических факторов на метаболизм некоторых гидробионтов/А. С. Константинов, В. Я. Пушкарь, О. В. Аверьянова//Изв. РАН. Сер. биол. 2003. 6. С. 1-7.
- Константинов А. С. Некоторые особенности роста рыб при переменных температурных режимах/А. С. Константинов, В. В. Зданович//Вопр. ихтиологии. 1986. Т. 26, № 3. С. 448-456.
- Макоедов А. Н. Основные тенденции развития аквакультуры/А. Н. Макоедов//Вопр. рыболовства. 2006. Т. 7, № 3. С. 366-384.
- Пушкарь В. Я. Установка для выращивания рыб/В. Я. Пушкарь, В. Н. Дубровин, В. Н. Григорьев: авторское свидетельство СССР. 1979. № 789082, МКИ: А01К 61/00.
- Сарвиро В. С. Экологическая оценка влияния термических колебаний на параметры роста бо-коплава Gammarus lacuslris/В. С. Сарвиро//Гидробиол. журн. 1983. Т. 19, 4, С. 71-73.
- Спотт С. Содержание рыбы в замкнутых системах/С. Спотт. М.: Легк. и пищ. пром-сть. 1983. 192 с.
- Стеффене В. Индустриальные методы выращивания рыбы/В. Стеффене. М.: Агропромиздат. 1985. 384 с.
- Schuenhoff A. A semi-recirculating, integrated system for the culture of fish and seaweed/A. Schuenhoff, M. Shpigel, I. Lupatsch, A. Ashkenazi, F. Msuya, A. A Neori//Aquaculture. 2003. V. 221, 1-4. P. 167-181.
- Schneider О. Analysis of nutrient flows in integrated intensive aquaculture systems/O. Schneider, V. Seveti.E. H. Eding,J. A. J. Verreth//Aquacultural Engineering. 2005. V. 32, № 3-4. P. 379-401.
- Biette R. M. Growth of underyearling salmon (Oncorhynchus nerka) under constant and cyclic temperatures in relation to live zooplankton ration size/R. M. Biette, G. H. Geen//Can. J. Fish. Aquat. Sci. 1980. V. 37, №2. P. 203-210.;,v
- Summerfeld S. T. Developments in recirculating systems for Arctic char culture in North America/S. T. Summerfeld, G. Wilton, D. Roberts, T. Rimmer, K. Fonkalsrud//Aquacultural Engineering. 2004. V. 30, № 1-2. P. 31-71.
- Effect of diel temperature fluctuations on growth, oxygen consumption and proximate body composition in the sea cucumber Aposiichopus japonicus Selenka/Y. Dong, S. Dong, X. Tian, F. Wang, M. Zhang//Aquaculture. 2006. V. 255,№ 1-4. P. 514-521.
- Takeuchi T. Effect on the growth and body composition of juvenile tilapia Oreochromis niloticus fed raw Spirulina/T. Takeuchi, J. Lu, G. Yoshizaki, S. Satoh//Fisheries Science. 2002. V. 68, № 1. P. 34-40.
- Gardibaldi L. List of animal species used in aquaculture/L. Gardibaldi//FAO Fish. Circ. 1996. № 914. P. 1-38.
- Neori A. Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofitration in modern mariculture/A. Neori, T. Chopin, M. Troell, A. H. Buschmann, G. P. Kraemer, C. Hailing, M. Shpigel, C. Yarish//Aquaculture. 2004. V. 231, № 1-4. P. 361-391.
- Integrated mariculture: asking the right questions/M. Troell, C. Hailing, A. Neori, T. Chopin, A. H. Buschmann,N. Kautsky,C. Yarish//Aquaculture. 2003. V. 226, № 1-4. P. 69-90.
- Liao I. C. Technical innovations in eel culture systems/I. C. Liao, Y. K. Hsu, W. C. Lee//Rev. Fish. Sci. 2002. V. 10, № 3-4. P. 433-450.
- Naegel L. C. A. Combined production of fish and plants in recirculating water/L. C. A. Naegel//Aquaculture. 1977. V. 10, № 1. P. 17-24.
- Neori A. A sustainable integrated system for culture of fish, seaweed and abalone/A. Neori, M. Shpigel, D. A Ben-Ezra//Aquaculture. 2000. V. 186, № 3-4. P. 279-291.
- Pilditch C. A. Effect of temperature fluctuations and food supply on the growth and metabolism of juvenile sea scallop (Placopecten magellanicus)/C. A. Pilditch, J. Grant//Mar. Biol. 1999. V. 134, P. 235-248.
- Palmegiano G. B. Spirulina as a nutrient source in diets for growing sturgeon (Adpenser baerii)/G. B. Palmegiano, E. Agradi, G. Fornesis, F. Gal, S. Gasco, E. Rigamonti, B. Sicuro, I. Zoccareto//Aquaculture Research. 2005. V. 36,№ 2. P. 188-195.
- Yarish C. The aquaculture of Porphyra leucosticta (Rhodophyta) for an integrated finfish/seaweed recirculating aquaculture system in an urban application/C. Yarish, P. He, R. Carmona, S. Liu, G. Kraemer, C. Neefus, T. Chopin, G. Nardi, J. Curtis, S. Lonergan, P. Trupp//Journal of Phycology. 2002. V. 38, № 1. P. 39-68.
- Tian X. The effects of thermal amplitude on the growth of Chinese shrimp Fenneropenaeus chinensis (Osbeck, 1765)/X. Tian,S. Dong//Aquaculture. 2006. V. 251, № 2-4. P. 516-524.
- Wang H. Using a macroalgae Ulva pertusa biofilter in a recirculating system for production of juvenile sea cucumber Aposiichopus japonicus/H. Wang, C. F. Liu, С. X. Qin, S. Q. Cao, J. Ding//Aquacultural Engineering. 2007. V. 36, № 3. P. 217-224.
