Рост и накопление массы Hylocomium splendens в лесном поясе Хибин

Автор: Ермолаева Ольга Владимировна, Шмакова Наталья Юрьевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (161), 2016 года.
Бесплатный доступ
Изучена динамика годичного прироста (линейная и по массе) в течение вегетации у Hylocomium splendens в лесном поясе Хибин. Основная доля линейного прироста (80 %) формируется в летний период (с максимумом в середине июля). Осенью, при замедлении линейного роста, активно происходит нарастание массы текущего годичного прироста. Проведено сравнение показателей роста в разных условиях произрастания. Наибольшие значения линейных и весовых показателей приростов характерны для местообитания в просветах крон деревьев. В среднем за 2013-2015 годы линейный прирост составил 18-28 мм, масса - 4-7 мг сухой массы. Рост H. splendens продолжается, по крайней мере, в течение 2 лет: формирование длины годичного прироста происходит в основном в первый год жизни, а нарастание массы продолжается и в следующем году. Построена статистическая модель зависимости скорости линейного роста H. splendens в просветах крон деревьев от температуры воздуха и количества осадков.
Линейный прирост, масса годичного прироста, со2-газообмен, хибины
Короткий адрес: https://sciup.org/14751124
IDR: 14751124 | УДК: 581.1
The growth and mass accumulation of Hylocomium splendens in Khibiny mountain forest belt
The annual increment dynamics (linear and mass) of Hylocomium splendens during vegetation periods in the forest belt of Khibiny mountains are examined. The main part of the linear growth (80 %) was formed in summer (with its maximum in mid-July). In autumn, the linear growth slowed down, but the mass of annual increment was actively growing. The growth indicators of moss in different conditions were compared. The linear and mass increments are the largest in the openings found between trees. The average growth of the linear increment in 2013-2015 was 18-28 mm/year, and the average growth of the dry mass amounted to 4-8 mg/year. It was revealed that the growth of H. splendens continued during two years: the length of the annual increment was formed mainly in the first year of its life, and the increase in the masses continued through the next year. A statistical model reflecting dependence of the growth rate of H. splendens in the openings between trees with main environmental factors is presented.
Текст научной статьи Рост и накопление массы Hylocomium splendens в лесном поясе Хибин
Изучение роста мхов важно для оценки состояния экосистем, в которых бриофиты составляют значительную долю среди компонентов общей биомассы. В Хибинах Hylocomium splen-dens встречается в разных растительных поясах. В лесных сообществах является доминантом и содоминантом, а в ряде местообитаний представляет собой микрогруппировки с проективным покрытием 80–100 %. Это гигромезофит, довольно чувствительный к условиям обитания, и его часто используют в качестве индикатора на влияние факторов среды. Имеет четкие, визуально отличающиеся границы годичных приростов.
Формирование годичной продукции H. splen-dens и факторы, влияющие на этот процесс, оценены в разных географических районах: в nonarctic North American [16], в Субарктике [11], в лесотундре и северной тайге Центральной Сибири [5], [6], лесоболотных комплексах Томской области [1], в темнохвойных насаждениях южной тайги юго-восточной части Западной Сибири [8], в различных типах леса Приангарья в Красноярском крае [3], в кедровых лесах Ямало-Ненецкого АО [2] и др.
Несмотря на значительное число исследований, данных для понимания особенностей ростовых процессов у H. splendens недостаточно. Результаты данного исследования являются продолжением работы, начатой в 2009 году [12].
Цель исследования – изучение особенностей роста и накопления массы мха в зависимости от местообитаний и метеоусловий.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводили в 2013–2015 годах в редкостойном березово-еловом лесу (около 300 м над ур. м.) на территории Полярно-альпийского ботанического сада, Хибины (67о38’ с. ш.). Природные условия этой территории подробно описаны [7].
Объект исследования – мох гилякомиум блестящий Hylocomium splendens (Hedw.) Bruch et al. (Hylocomiaceae), представитель бокоплодных мхов. Встречается на почве и разложившейся древесине. Образует рыхлые зеленые или желтовато-зеленые дерновинки. Стебель состоит из этажно-расположенных облиственных веточек, каждый этаж соответствует годичному побегу за год. Наблюдения и замеры проводили на годичных приростах, которые развивались посередине обветвленной части стебля предыдущего года.
Пробные площади были заложены в местообитаниях H. splendens с разным уровнем освещенности (освещенность в солнечный день/пасмур-ный день): ПП1 – в просветах крон деревьев (30/5 клк) и ПП2 – открытое местообитание (70/10 клк). В 2013 году измерения длины и массы годичного побега были проведены в течение вегетации, в 2014–2015 годах – в конце вегетационного пери- ода. За начало и конец вегетационного периода принимали время, когда наблюдали устойчивый переход температуры воздуха через +5 оС. Линейный прирост измеряли на маркированных особях (по 50–100 штук в куртинах каждого местообитания). Для определения массы прироста отбирали особи из аналогичных куртин, находящихся рядом. Периодичность измерений составляла от 5 до 15 суток. На дату измерения определяли длину (мм) и сухую массу прироста (мг), рассчитывали скорость роста побега (мм или мг в сутки) за период между измерениями. Интенсивность СО2-газообмена годичных побегов определяли с помощью инфракрасного газоанализатора Гамма-100 (Россия). Контроль за температурой воздуха проводили измерительными комплексами (модель iBDLR-3-U-Х). Количество осадков определяли с помощью осадкомеров.
Температурный режим и количество осадков за вегетационные периоды представлены в табл. 1. В целом 2013 год характеризовался ранним началом вегетационного периода (с 16 мая), относительно высокими и выровненными температурами весеннего и летнего периодов с минимальным количеством осадков. Отмечено высокое значение суммы эффективных (1516 оС) и активных температур (1271 оС) в сезоне. Вегетационный период 2014 года отличался холодной и длительной весной, жарким летом и холодной осенью. 2015 год был самым холодным, с аномально низкой температурой в июле и почти ежедневными осадками. Сумма активных температур (+10 оС и выше) за сезон оказалась очень низкой - всего 712 оС, что на 34 оС меньше многолетних данных для территории ПАБСИ [9].
Статистическая обработка полученных данных выполнена с использованием универсальной программы STATISTICA 10.0.
Таблица 1
Среднемесячные показатели температуры воздуха и количества осадков за вегетационный период
|
Год |
2013 |
2014 |
2015 |
Среднемесячная температура, оС
|
VI |
13,8 |
7,8 |
9,5 |
|
VII |
14,7 |
16,6 |
10,5 |
|
VIII |
13,4 |
13,0 |
11,6 |
|
IX |
7,7 |
6,6 |
8,4 |
|
В среднем |
12,4 |
11,0 |
10,0 |
|
за вегетацию |
Количество осадков, мм
|
VI |
23 |
20 |
97 |
|
VII |
29 |
36 |
65 |
|
VIII |
23 |
62 |
34 |
|
IX |
11 |
29 |
61 |
|
Всего |
86 |
147 |
257 |
|
за вегетацию |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сезонная динамика длины и массы годичного прироста
Накопление фитомассы и изменение длины прироста являются связанными показателями, но фитомасса – более информативный показатель продуктивности [15]. В зависимости от поставленной цели некоторые исследователи изучают лишь линейный прирост мхов [4], [10].
В 2013 году для H. splendens на ПП1 изучение сезонной динамики линейного годичного прироста проведено одновременно с изменением его массы. В весенний и раннелетний период нарастание годичного побега в длину идет более интенсивно, чем накопление массы (рис. 1). Несмотря на продолжительную весну в этом году, к началу летнего периода длина прироста составила 5,7 мм (скорость роста не превышала 0,2 мм/день) и была на 28 % меньше, чем в благоприятный по условиям сезон 2009 года, когда скорость роста составляла 0,3–0,35 мм/ день. Для H. splendens высокая скорость роста была отмечена нами при следующем сочетании факторов: температура – 10–13 оС, осадки 2–4 мм в день [12]. В 2013 году повышенная температура воздуха (12–18 оС) и осадки меньше 2 мм в день (рис. 1С), вероятно, не позволили реализовать ростовые возможности. Наибольшая скорость роста отмечена во второй половине июля – 0,5 мм/день, что на 30–40 % ниже, чем в благоприятные по климатическим условиям годы. Увеличение массы прироста связано с началом его ветвления и наблюдается также с середины июля. В летний период скорость нарастания массы составила 0,06–0,08 мг/день. К концу летнего периода длина линейного прироста составляет уже 80 %, а масса всего 56 % от годичной величины (27,7 мм и 6,7 мг, табл. 2). Осенью интенсивность удлинения снижается, но увеличивается масса прироста. При этом скорость нарастания массы была в 1,6 раза выше (0,13 мг/день), чем летом. Между длиной и массой приростов на ПП1 установлена высокая корреляционная связь (r = 0,9).
На основе регрессионного анализа материалов трехлетних наблюдений (2009, 2012–2013 годы, n = 33) была построена следующая статистическая модель зависимости скорости линейного роста (Y, мм/день) H. splendens на ПП1 от температуры воздуха (T, оС) и количества осадков (P, мм) за весь период вегетации:
Y = –0,501 + 0,098Т – 0,003T2 + 0,083P – 0,007Р2 + 0,001ТР
(R = 0,636; R2 = 40,4 %; F (5, 27) = 3,66 > F0,05 = 2,56; p < 0,01), где R – коэффициент множественной корреляции, R2 – коэффициент множественной детерминации; F – расчетное значение критерия Фишера (в скобках после буквы F – число степеней свободы).
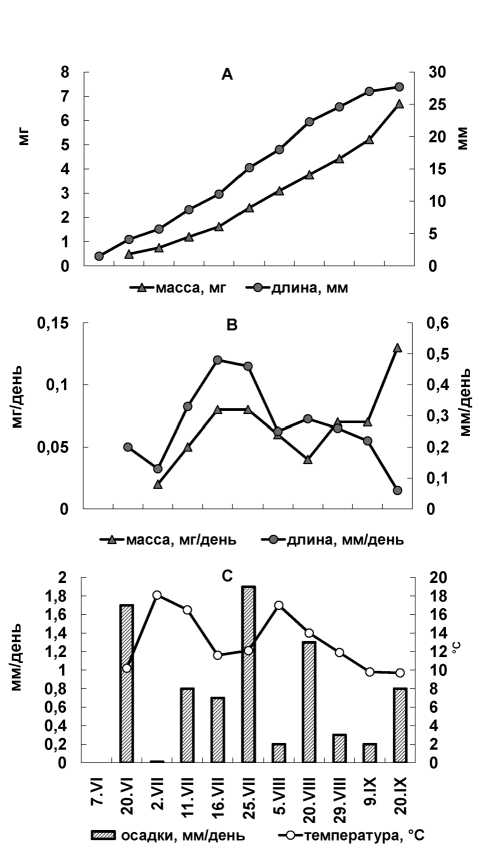
Рис. 1. Динамика роста (линейная и по массе) годичного прироста мха Hylocomium splendens в просветах крон деревьев в течение вегетации 2013 года (А); суточный прирост в периоды между измерениями (В); количество осадков и температура воздуха в периоды между измерениями (С)
Данное уравнение можно использовать для прогнозирования скорости роста H. splendens в длину в реальном диапазоне вариаций мете-
орологических факторов за период вегетации. Необходимо отметить, что характер связи роста H. splendens с метеоусловиями отличался в разные периоды вегетации. Летом корреляция между скоростью роста и температурой была теснее (r = 0,50; р < 0,05), чем с осадками (r = 0,41; р < 0,05). Осенью (на фоне пониженных температур воздуха) зависимость ростовых процессов H. splendens от температурного фактора усиливалась (r = 0,76; р < 0,01), а с осадками приобретала выраженную отрицательную направленность (r = –0,44; р < 0,05).
Годичный прирост в разных местообитаниях
Как видно из данных табл. 2, более высокие значения длины и массы приростов H. splendens характерны для местообитаний на ПП1, причем величины линейного прироста оказались одинаковыми при разной характеристике вегетационных сезонов, а различия отмечены только по массе. Колебания величин линейного прироста по годам наблюдений сильнее выражены на ПП2. Диапазон варьирования массы годичного прироста в годы наблюдений одинаков в обоих местообитаниях. Удельный вес прироста (нарастание массы относительно длины годичного прироста, М/Л) мало изменчив в разные годы наблюдений. Самый низкий годичный прирост отмечен в 2013–2014 годах на ПП2. В дождливый и холодный сезон вегетации (2015) различия в величине длины прироста в разных условиях обитания выражены менее, чем в жаркие и сухие годы.
В среднем за годы исследований (2013–2015) длина линейного прироста у H. splendens на ПП1 составила 27,2 ± 1,7 мм, масса – 6,9 мг, на ПП2 – меньше на 28 % и 38 % соответственно. Следует отметить, что длина годичного прироста на 20 % меньше, чем в благоприятные по климатическим факторам годы (2009, 2012), когда величина годичного прироста достигала 35 мм [12]. Недостаток осадков и высокие температуры летних периодов 2013–2014 годов привели к замедлению роста в длину, не позволяя реализовать ростовой потенциал. В этих условиях наблюдали высыхание верхнего органогенного слоя почвы и развитие водного стресса у мхов, что особенно
Таблица 2
Длина и масса годичного прироста Hylocomium splendens в конце вегетационного периода в зависимости от местообитания
|
Год |
ПП1 |
ПП2 |
||||||
|
n |
L |
M |
M/L |
n |
L |
M |
M/L |
|
|
2013 |
47 |
27,7 ± 0,7 |
6,7 |
0,24 |
44 |
18,2 ± 0,5 |
4,0 |
0,22 |
|
2014 |
65 |
26,9 ± 0,6 |
7,4 |
0,28 |
174 |
18,7 ± 0,4 |
4,4 |
0,24 |
|
2015 |
97 |
27,0 ± 0,7 |
6,6 |
0,24 |
46 |
22,2 ± 0,8 |
4,5 |
0,20 |
|
среднее |
27,2 ± 1,7 |
6,9 ± 0,3 |
0,25 |
19,7 ± 1,3 |
4,3 ± 0,3 |
0,22 |
||
Примечание. L – длина годичного прироста (мм); M – масса годичного прироста (мг сухого веса); M/L – нарастание массы относительно длины (удельный вес прироста, мг/мм).
заметно у особей на открытых местообитаниях. В 2015 году при низких температурах и достаточном количестве осадков линейный годичный прирост также небольшой.
Представленные данные сравнимы с имеющимися сведениями по H. splendens в других районах и экосистемах России. Максимальный линейный годичный прирост имеет тенденцию к увеличению от лесотундры к северной тайге. Так, в кедровых лесах Ямало-Ненецкого района линейный прирост составил 14–21 мм, масса 7 мг [2]; в лесоболотных комплексах юга Западной Сибири годичный прирост – 19–26 мм, масса – 19–27 мг [1]; в различных типах леса Приангарья линейный прирост варьирует от 10 до 23 мм, масса – от 5 до 14 мг [3]; в разных лесорастительных зонах Сибири годичный прирост составляет 9–15 мм, масса – 4–11 мг [5].
Измерение длины и массы годичного прироста 2014 года в течение 2 лет жизни (рис. 2) показало, что в конце второго года жизни длина годичного прироста в обоих местообитаниях почти одинакова, различия видны только по массе. В варианте ПП1 масса годичного прироста в конце 2-го года жизни увеличилась на 37 %, ПП2 – на 46 %. Удельный вес прироста изменился на ПП1 от 0,28 до 0,40, на ПП2 от 0,24 до 0,38. Таким образом, удлинение годичного прироста происходит в первый год жизни, а нарастание массы продолжается в следующем году и увеличивается на 40–50 %, что указывает на рост H. splendens , по крайней мере, в течение 2 лет.
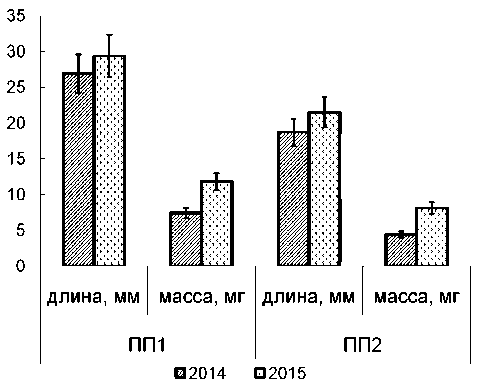
Рис. 2. Изменение длины и массы годичного прироста Hylocomium splendens (2014 – годичный прирост текущего года, 2015 – прирост 2-го года жизни)
Масса годичного прироста, нарастающая за год, определяется фотосинтетической продуктивностью. Интенсивность углекислотного газооб- мена H. splendens измеряли в приростах текущего и второго года жизни в разных местообитаниях (табл. 3). Уровень газообмена в приростах текущего года несколько выше на ПП1 по сравнению с ПП2. В приростах второго года жизни уровень газообмена был ниже, но на ПП1 на 40 %, а на ПП2 на 20 %. Вероятно, этим можно объяснить большее изменение массы в течение второго года жизни на ПП2.
Таблица 3
Интенсивность фотосинтеза Hylocomium splendens (мг СО2/гсух. в. ч) в разных местообитаниях
|
Годичный прирост |
ПП1 |
ПП2 |
|
Текущего года |
5,00 ± 0,58 |
4,68 ± 0,49 |
|
Второго года жизни |
3,05 ± 0,33 |
3,71 ± 0,41 |
Примечание. n = 16, содержание воды в побегах не менее 60 %.
Таким образом, формирование годичного прироста H. splendens продолжается, по крайней мере, в течение 2 лет. Рост годичного прироста в длину завершается в основном в первый год жизни, а формирование массы происходит в течение двух лет. Большая доля линейного прироста формируется в летний период, а его массы -в осенний. Полученные нами данные о разделении во времени линейного роста и накопления биомассы согласуются с результатами, полученными для таких видов, как Brachythecium rutabu-lum, Thuidium tamariscinium и Lophocolea biden-tata, и не согласуются для Plagiomnium undulatum и Pseudoscleropodium purum [14].
Более высокие значения показателей линейных размеров и массы приростов характерны для местообитаний на ПП1 – в более затененных условиях (просветах крон), что также согласуется с регуляторной ролью освещенности в линейном росте мха в работе [13], где в условиях низкой освещенности отмечалось значительное вытягивание растений.
Величина годичной продукции мхов зависит от количества осадков и температуры воздуха вегетационного периода. Условиями, ингибирующими рост мхов в летний период, являются относительно высокие температуры и небольшое количество осадков, особенно в условиях открытых местообитаний.
Сопоставимость полученных данных с аналогичными для других регионов с разными условиями вегетационных периодов свидетельствует о генетической детерминированности потенциальных значений функциональной ростовой активности H. splendens и возможности его реализации на большом градиенте условий.
THE GROWTH AND MASS ACCUMULATION OF HYLOCOMIUM SPLENDENS IN KHIBINY MOUNTAIN FOREST BELT
The annual increment dynamics (linear and mass) of Hylocomium splendens during vegetation periods in the forest belt of Khibiny mountains are examined. The main part of the linear growth (80 %) was formed in summer (with its maximum in mid-July). In autumn, the linear growth slowed down, but the mass of annual increment was actively growing. The growth indicators of moss in different conditions were compared. The linear and mass increments are the largest in the openings found between trees. The average growth of the linear increment in 2013–2015 was 18–28 mm/year, and the average growth of the dry mass amounted to 4–8 mg/year. It was revealed that the growth of H. splendens continued during two years: the length of the annual increment was formed mainly in the first year of its life, and the increase in the masses continued through the next year. A statistical model reflecting dependence of the growth rate of H. splendens in the openings between trees with main environmental factors is presented.
Список литературы Рост и накопление массы Hylocomium splendens в лесном поясе Хибин
- Гончарова И. А., Беньков А. В. Динамика приростов зеленых мхов в лесоболотных комплексах юга Западной Сибири//Лесоведение. 2005. № 1. С. 43-51
- Гончарова И. А. Продуктивность Hylocomium splendens (Hylocomiaceae) в кедровых лесах Ямало-Ненецкого автономного округа//Растительные ресурсы. 2013. Т. 49. Вып. 2. С. 175-180.
- Гончарова И. А., Собачкин Р. С. Годичные приросты и годичная продукция мхов Helodium blandowii (Thuidiaceae) и Hylocomium splendens (Hylocomiaceae) в лесах Приангарья//Растительные ресурсы. 2013. Т. 49. Вып. 4. С. 490-497.
- Грабовик С. И. Динамика годичного прироста у некоторых видов Sphagnum L. в различных комплексах болот Южной Карелии//Растительные ресурсы. 2002. Т. 38. Вып. 4. С. 62-68.
- Кнорре А. А., Ваганов Е. А. Особенности роста и годичная продукция Hylocomium splendens (Hylocomiaceae) в северных экосистемах//Растительные ресурсы. 2005. Т. 41. Вып. 4. С. 12-21.
- Кнорре А. А., Кирдянов А. В., Ваганов Е. А. Изменчивость годичной продукции надземной фитомассы основных доминантов высокоширотных сообществ Центральной Сибири//Растительные ресурсы. 2007. Т. 43. Вып. 1. С. 3-17.
- Костина В. А., Белкина О. А, Константинова Н. А. Краткий очерк природных условий//Мохообразные и сосудистые растения территории Полярно-альпийского ботанического сада. Апатиты: Изд-во КНЦ РАН, 2001. С. 8-15.
- Кошурникова Н. Н. Годичная продукция мохового яруса в темнохвойных лесах Кеть-Чулымского лесорастительного округа (на примере гилякомиума блестящего Hylocomium splendens)//Известия РАН. Серия биологическая. 2007. Вып. 5. С. 636-640.
- Семко А. П. Режим тепла и влаги для роста и развития дикорастущих и интродуцированных растений в центральной части Кольского полуострова. Апатиты: Изд-во КНЦ АН СССР, 1989. 30 с.
- Тимлянова А. А., Косых Н. П., Миронычева-Токарева Н. П. Прирост болотных растений//Сибирский журнал экологии. 2000. № 5. С. 653-658.
- Callaghan T. V., Carlson B. A., Sonesson M., Temesvary A. Between-year variation in climate-related growth of circumarctic populations of the moss Hylocomium splendens//Funct. Ecol. 1997. Vol. 11. P. 157-165.
- Ermolaeva O. V., Shmakova N. Yu., Lukyanova L. M. Of the crowth of Polytrichum, Pleurozium and Hylocomium in the forest belt of the Khibiny Mountains//Arctoa. 2013. Vol. 22. P. 7-14.
- Glime J. M. Bryophyte Ecology. 2007. Vol. 1. Chapters 4, 5, 12. Available at: http://www.bryoecol.mtu.edu (assecced 12.05.2016).
- Rincon E., Grime J. P. Plasticity and light interception by six bryophytes of contrasted ecology//J. Ecol. 1989. Vol. 77. P. 439-446.
- Stark L. R. Invited essay, new frontiers in bryology and lichenology: phenology and its repercussions on the reproductive ecology of mosses//Bryologist. 2002. Vol. 105. P. 204-218.
- Vill D. H. Growth and production dynamics of boreal mosses over climatic, chemical and topographic gradients//Bot. J. Linn. Soc. 1990. Vol. 104. P. 35-59.
