Рост потомства ели сибирской в густых и редких испытательных культурах

Автор: Рогозин Михаил Владимирович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3-4, 2011 года.
Бесплатный доступ
Приведены результаты изучения испытательных культур в 21 и 23-летнем возрасте с анализом структуры древостоев материнских популяций из естественных и искусственных насаждений ели сибирской. Обосновывается отбор материнских особей и испытание их потомства в условиях оптимальной густоты выращивания древостоев.
Ель сибирская, происхождение, конкуренция, испытательные культуры
Короткий адрес: https://sciup.org/147204556
IDR: 147204556 | УДК: 630
The growth of the offspring of Siberian spruce in a luxuriant and rare test cultures
Presents the results of studying the test crops in 21 and 23 years of age with the analysis of the structure of forests in the parent populations from natural and artificial stands of spruce siberian. The author justifies the selection of the parent individuals and test their offspring in conditions of optimum density of cultivation of forests.
Текст научной статьи Рост потомства ели сибирской в густых и редких испытательных культурах
Методика
Исследованы потомства 525 деревьев в 21–23летнем возрасте. Семена получены в 1986 г. от 453 плюсовых и кандидатов в плюсовые деревья, 24 минусовых и 48 нормальных деревьев в 12 различных типах насаждений (ценопопуляциях), в основном в подзоне южной тайги. Контрольные образцы получены от 20–30 случайных деревьев в каждом из насаждений. Заготовку семян проводили путем отстрела шишек дробью из охотничьего ружья. От каждого дерева получено по 700–2000 шт. семян, которые были разделены на две части. Первая часть (до 500 шт.) высевалась в 1987 г. в 4 повторностях в теплицу и спустя 2 года пересаживалась в 4–7 повторностей в школьное отделение. В 4-летнем возрасте весной 1991 г. саженцы высадили в Ильинском лесничестве на площади 11.0 га на среднесуглинистую почву после сплошной раскорчевки вырубки. Растения в испытательные культуры (ИК) высаживали по схеме 2.5×1.0 м, в 4–6 повторностей на делянки в виде отрезков рядов из 5–9 растений.
Вторая часть семян, от 300 до 500 шт., высевалась в 1988 г. в 4 повторностях в орошаемый питомник. В 3-летнем возрасте растения были высажены в испытательные культуры на супесчаную почву, подстилаемую с 60–80 см плотным опесчаненным суглинком, в раскорчеванные на вырубке полосы в кв. 5 бывшего Юго-Камского лесничества Оханского лесхоза, на площади 8.0 га.
Из-за недостатка площади было принято решение о повышении густоты и посадке на одной раскорчеванной полосе 3 рядов культур со схемой по- садки 0.7×0.7 м. Такая схема не оптимальна для выявления продуктивных семей, но она позволила нам выяснить реакцию потомства на усиление конкуренции. На этом участке культуры в течение 20 лет конкурировали также с осиной и березой, которые местами превышали высоту ели на 1–5 м. Это обстоятельство мы использовали для выяснения самых начальных тенденций эволюции ели в условиях повышения конкурентного давления соседних деревьев.
Экспериментальная часть.
Результаты и их обсуждение
Особый интерес представляло поведение потомства от культур, заложенных в начале ХХ в. Ф.А. Теплоуховым, сыном А.Е. Теплоухова, основателя династии ученых-лесоводов на Урале. Это были культуры происхождений Сепыч, Очер и Верещаги-но, от которых нами испытывалось потомство 129 плюсовых деревьев.
Данные о росте потомства ели на первом участке в 8 лет и в 21 год обсуждались ранее (Рогозин, 2008; Рогозин, Жекин, 2010). В 2010 г. впервые были получены высоты потомств на втором (густом) участке ИК в 23-летнем возрасте. Сравнение высот потомств в редких и густых ИК показало, что различия между высотами достоверны у 9 происхождений из 12 (табл. 1). Это свидетельствует о разной реакции происхождений на изменение как почвенных, так и фи-тоценотических условий. При этом незначительно и недостоверно изменили свои высоты всего 3 цено-популяции (происхождения) из числа естест-
Таблица 1
Рост потомства ели сибирской (12 ценопопуляций) в испытательных культурах
|
Местонахождение (поселение, где размещалось лесничество) |
Число деревьев» шт |
Материнские деревья, средние значения |
Потомство в кв 41 Ильинского л-ва |
Потомство в кв 5 бывш Юго-Камского л-ва |
Различие в росте по-томств на участках |
||||||
|
возраст, лет |
высота |
диаметр, см |
число рас гений, шт |
средняя высота, % |
число растений, шт |
средняя высота, % |
±, % |
достоверность, t-критерий Стьюдента |
|||
|
м |
% |
||||||||||
|
Плюсовые деревья, естественные насаждения |
|||||||||||
|
Очер |
13 |
130 |
30 3 |
1162 |
45 3 |
459 |
108 0± 1 66 |
136 |
97 6 ±0 76 |
-104 |
5 69** |
|
Нытва |
29 |
95 |
29 1 |
120 3 |
40 8 |
834 |
105 9± 1 17 |
217 |
113 7± 1 04 |
7 8 |
4 98** |
|
Гайва* |
74 |
124 |
30 6 |
124 2 |
43 8 |
1898 |
95 4 ± 0 65 |
737 |
106 0i 0 50 |
106 |
12 93** |
|
Ильинск |
32 |
104 |
28 2 |
126 6 |
38 5 |
1010 |
101 8± 1 06 |
366 |
107 0± 0 78 |
5 2 |
3 95** |
|
Пермь |
108 |
118 |
30 1 |
125 6 |
43 2 |
2918 |
102 6± 0 57 |
1055 |
Ю24±040 |
-0 2 |
0 29 |
|
Кунгур |
27 |
102 |
31 7 |
122 3 |
47 3 |
785 |
102 7± 1 14 |
195 |
101 6±061 |
-1 1 |
0 85 |
|
Чусовой |
18 |
133 |
30 |
129 3 |
43 9 |
5)5 8419 |
100 2± 1 44 |
109 |
97 9 ± 1 12 |
-2 3 |
1 26 |
|
Всего; в среднем |
301 |
115 |
30 |
123.5 |
43.3 |
102.4 ±0.35 |
2815 |
104.4 ±0.25 |
1.9 |
4.42** |
|
|
Члюсовые деревья, лесные кузьтуры |
|||||||||||
|
Сепыч-1 |
85 |
77 |
28 |
126 1 |
352 |
2628 |
1078±062 |
1026 |
111 0± 0 38 |
32 |
4 40** |
|
Сепыч-П |
9 |
74 |
28 8 |
116 1 |
36 2 |
239 |
НО 4 г 1 84 |
79 |
99 8 ± 1 45 |
-106 |
4 52** |
|
Очер |
16 |
88 |
31 4 |
122 |
36 2 |
41 1 |
1107Т 1 53 |
173 |
106 2 ± 1 35 |
-4 5 |
2 21** |
|
Верещагине |
19 |
87 |
29 4 |
122 4 |
35 1 |
474 |
97 3 т 1 34 |
259 |
118 9± 0 69 |
21 6 |
14 33** |
|
Нижняя Курья* |
23 |
51 |
22 5 |
120 3 |
253 |
595 |
100 0i 1 38 |
195 |
108 1 ± I 07 |
8 1 |
4 63** |
|
Всего; в среднем |
152 |
75.4 |
28 |
121.4 |
33.6 |
4347 |
105.1 ±0.47 |
1732 |
110.9 ±0.31 |
5.7 |
10.12** |
|
Нормальные и минусовые деревья из лесных культур |
|||||||||||
|
Сепыч-1 |
19 |
77 |
24 3 |
109 6 |
22 4 |
495 |
101 0± 1 43 |
230 |
108 0± 0 99 |
7 0 |
4 02** |
|
Сепыч-П |
15 |
74 |
24 5 |
98 8 |
25 7 |
310 |
105 5± 1 65 |
152 |
92 6 ± 0 79 |
-129 |
7 05** |
|
Очер |
13 |
88 |
28 2 |
109 9 |
25 5 |
319 |
103 3± 1 73 |
136 |
102 0± 0 79 |
-1 3 |
0 68 |
|
Верещагине |
11 |
87 |
22 8 |
95 2 |
20 5 |
278 |
97 7±2 17 |
73 |
104 1 ± 1 72 |
64 |
2 31** |
|
Нижняя Курья* |
14 |
51 |
20 8 |
1114 |
22 |
409 |
95 6 ± 1 44 |
147 |
108 7 ± 1 И |
13 1 |
721** |
|
Всего; в среднем |
72 |
75.4 |
24.2 |
104.98 |
23.2 |
1811 |
100.8 ±0.74 |
738 |
103.5 ±0.47 |
2.60 |
2.97** |
|
Контроль, 12 ценопопуляций |
776 |
100.0 ± 1.06 |
246 |
100.0 ±0.43 |
0 |
0 |
|||||
|
Примечания *-микрорайон Перми **- различие достоверно при t00s |
|||||||||||
М В Рогозин
Таблица 2
Густота, высота, сбег ствола и средний прирост в высоту в культурах (плюсовые деревья и рост их потомства в 21-23 года)
|
Материнские ценопопуляции (культуры) |
Рост потомства (высота) в испытательных культурах |
|||||||||||||
|
Происхождение культур |
Возраст куль-тур, лет |
Схема посадки,м |
Текущая густота |
Средние |
Сбег ствола |
Средний прирост в высоту |
в 21 год на суглинистой почве с расстоянием между растениями в ряду 1.0 м |
в 23 года на супесчаной почве с расстоянием между растениями в ряду 0.7 м |
||||||
|
шт./га |
ранг * |
вы сота, м |
диаметр, см |
см/м |
ранг |
см /в год |
ранг |
% от контроля |
ранг |
% от контроля |
ранг |
|||
|
Сепыч-2 |
74 |
2 134.07 |
770 |
1 |
24.8 25.7 |
25.3 |
1.020 |
1 |
33.5 |
1 |
110.4 |
2 |
99.8 |
5 |
|
Очер |
88 |
2.134.07 |
920 |
2 |
25.1 |
0 977 |
2 |
29.2 |
2 |
110.7 |
1 |
106.2 |
4 |
|
|
Сепыч-1 |
77 |
2.134.07 |
1200 |
3 |
22.2 |
21.1 |
0.950 |
3 |
28.8 |
3 |
107.8 |
3 |
111.0 |
2 |
|
Верещагине |
87 |
2.13x1.07 |
1256 |
4 |
24.0 |
21.2 |
0 883 |
4 |
27 6 |
4 |
97.3 |
5 |
118.9 |
1 |
|
Нижняя Курья |
51 |
2.0x0.60 |
1548 |
5 |
18.7 |
16.2 |
0.866 |
5 |
36.7 |
** |
100.0 |
4 |
108.1 |
3 |
Примечания. * - ранги идут по возрастанию, в других случаях - по убыванию: ** - прирост не ранжировали, так как культуры были намного моложе, чем на первых четырех участках.
Рост потомства ела сибирской в густых и редких испытательных культурах
венных насаждений (Пермь, Кунгур, Чусовой), у которых высоты близки к средним значениям. Эти происхождения устойчивы в росте, но не имеют перспективы в селекции.
В целом потомство естественных насаждений при увеличении конкуренции повысило свой рост в среднем на 2.0%, а потомство культур – на 5.8%. Это может свидетельствовать о том, что материнские деревья в культурах сформировались в целом в более конкурентной среде. Поэтому они лучше переносят конкурентное давление соседей, и некоторые из них даже повышают рост в высоту при более густой посадке в рядах культур (табл. 1).
К устойчиво сохраняющим на обоих участках быстрый рост можно отнести культуры происхождение из Нытвы, имеющее высоты 105.9 и 113.7%, а также происхождения из культур Сепыч-1 и Очер с высотами, соответственно, 107.8 и 111 и 110.7 и 106.2%.
Происхождения из культур во всех случаях достоверно изменили свой рост и это требует подробного анализа причин таких изменений. Для анализа используем таксационные характеристики культур, поместив их в табл. 2.
По мере увеличения текущей густоты, при которой сформировались плюсовые деревья, высота потомства в редких испытательных культурах падает со 110.4 до 97.3–100%, и ранги высоты снижаются со 1–2 до 4–5. В густых испытательных культурах все наоборот: высота потомства возрастает в соответствии с увеличением густоты в материнских древостоях. Данное явление хорошо иллюстрирует
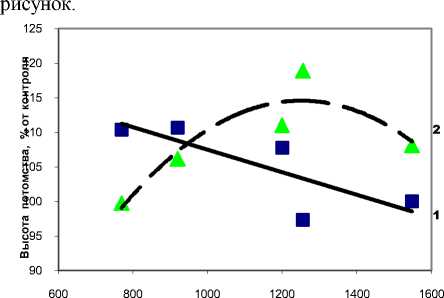
Густота в материнских популяциях, шт./га
Густота древостоев ели в материнских ценопопуляциях и ее влияние на высоту потомства в испытательных культурах:
-
1 – с редким размещение в рядах (через 1.0 м);
-
2 – с более густым размещением (через 0.7 м)
Таким образом, можно полагать, что если требуется выращивать потомство в условиях повышенной густоты, то для этого случая нужно отбирать родителей (плюсовые деревья) также в густых популяциях. И наоборот, при оптимальной конкуренции формируются материнские деревья, которые передают потомкам способность реализовать быстрый рост при снижении густоты их выращивания.
Последние родители нужны для плантационного выращивания их потомства. Для отбора таких родителей подходят естественные и искусственные древостои, редкие (или специально разреженные) в молодом возрасте и максимально сомкнутые в 50–60 лет. При этом придется смириться со слабой очи-щаемостью стволов от сучьев, которые у ели еще не опадают к этому возрасту и при отборе материнских особей по этому критерию вполне допустима средняя толщина сучьев.
Заключение
Таким образом, несколько упрощая ситуацию, можно сделать заключение о том, что у ели сибирской «наследуется» не столько быстрота роста плюсовых деревьев и популяций, сколько их реакция на условия конкурентной среды, в которой сформировалась популяция и конкретный древостой. Эту реакцию можно либо формировать в родительских древостоях ранними разреживаниями и затем ждать, когда плюсовые деревья в ней вырастут, либо искать популяции с оптимальной густотой. Историю конкурентного давления на дерево отражает сбег ствола и отбор плюсовых деревьев с его учетом (Рогозин, Жекин, 2010) в 2 раза повышает качество потомства, предназначенного для выращивания в условиях оптимальной густоты.
Список литературы Рост потомства ели сибирской в густых и редких испытательных культурах
- Рогозин М.В. Итоги 8-летних испытаний 525 семей ели сибирской в Пермском крае//Лесное хозяйство. 2008. № 1. С. 37-38.
- Рогозин М.В., Жекин А.В. Лесные генетические резерваты Пермского края и селекция ели сибирской//Хвойные бореальной зоны. 2010. Т. 27, № 1-2. С. 185-190.
