Рост проростков гречихи обыкновенной (Fagopyrum esculentum Moench) и накопление первичных и вторичных метаболитов при различных условиях минерального питания
 и накопление первичных и вторичных метаболитов при различных условиях минерального питания")
Автор: Цыпурская Е.В., Казанцева В.В., Фесенко А.Н., Загоскина Н.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Физиология адаптации
Статья в выпуске: 5 т.54, 2019 года.
Бесплатный доступ
Гречиха обыкновенная ( Fagopyrum esculentum Moench) - важная сельскохозяйственная культура. Помимо уникальных пищевых характеристик, для нее характерно образование различных фенольных соединений, в том числе рутина, широко применяемого в медицине. Поскольку фенольные соединения обладают высокой биологической и антиоксидантной активностью, в том числе рассматриваются как потенциальные компоненты функционального питания, изучение их накопления на начальных этапах онтогенеза растений представляет практический интерес. В представленной работе мы показали регуляторное действие макро- и микроэлементов на рост растений и накопление первичных и вторичных метаболитов у гречихи. Впервые охарактеризовано образование первичных и вторичных метаболитов в надземных органах нового и перспективного российского сорта гречихи обыкновенной Даша, внесенного в Государственный реестр селекционных достижений, допущенных к использованию в Российской Федерации, в 2018 году. Целью работы было изучение начальных этапов онтогенеза проростков гречихи обыкновенной, включая оценку их морфофизиологических характеристик в условиях различной обеспеченности элементами минерального питания, а также накопления в семядольных листьях фотосинтетических пигментов, сахаров и фенольных соединений. Объектом исследования служили проростки гречихи обыкновенной сортов Девятка и Даша селекции ВНИИ зернобобовых культур (г. Орел), внесенных в Государственный реестр селекционных достижений, допущенных к использованию в Российской Федерации, соответственно в 2004 и 2018 году. Растения выращивали в лабораторных условиях рулонным способом на воде (контроль) и питательной среде Хогланда-Арнона (опыт) при 24 °C и 16-часовом освещении. Определяли высоту гипокотилей, длину корней и массу семядольных листьев проростков. Оводненность растительного материала анализировали после его высушивания до постоянной массы при температуре 70 °C. Спектрофотометрическим методом определяли количество хлорофиллов a и b (при λ = 665 нм и λ = 649 нм), каротиноидов (λ = 440 нм), сахаров (λ = 490 нм), суммы растворимых фенольных соединений (λ = 725 нм), флавоноидов (λ = 415 нм) и фенилпропаноидов (λ = 330 нм) в этанольных экстрактах, полученных из семядольных листьев проростков разного возраста. Выращивание гречихи на питательной среде Хогланда-Арнона способствовало более быстрому росту надземных органов по сравнению с выращиванием на воде. Рост подземных органов в обоих случаях был одинаковым. Выявлены различия в накоплении фотосинтетических пигментов (хлорофилла a и b, каротиноидов) и сахаров в семядольных листьях у проростков: в опытных вариантах в большинстве случаев эти показатели были выше, чем в контрольных. Накопления фенольных соединений не столь явно зависело от условий минерального питания. Исключением было содержание фенилпропаноидов, которое в семядольных листьях проростков, выращенных на питательной среде, изменялось в большей степени по сравнению с контролем и достигало высоких значений в конце периода исследования. Следует также отметить, что при выращивании проростков гречихи сорта Даша на питательной среде в их семядольных листьях на поздних этапах онтогенеза отмечалось более значительное накопление пигментов (на 20 %, р ≤ 0,05), сахаров (на 33 %, р ≤ 0,05) и фенольных соединений (на 35 %, р ≤ 0,05) по сравнению с сортом Девятка. Таким образом, количество минеральных элементов имеет важное значение для начальных этапов онтогенеза растений гречихи обыкновенной. Более быстрый рост проростков и накопление первичных и вторичных метаболитов в их листьях характерны для опытных вариантов по сравнению с контролем. Использование различных условий минерального питания дает возможность регулировать рост и развитие растений, а также накопление в них различных метаболитов.
Гречиха, проростки, онтогенез, минеральное питание, пигменты, сахара, фенольные соединения, фенилпропаноиды, флавоноиды
Короткий адрес: https://sciup.org/142226263
IDR: 142226263 | УДК: 633.12:581.1:631.811 | DOI: 10.15389/agrobiology.2019.5.946rus
Growth of buckwheat (Fagopyrum esculentum Moench) seedlings and the accumulation of primary and secondary metabolites under various mineral nutrition conditions
Buckwheat ( Fagopyrum esculentum Moench) is an important agricultural crop; Russia, China and Ukraine are the world leaders of its production. In addition to the unique nutritional characteristics, it is characterized by the formation of various phenolic compounds including rutin widely used in medicine. The study of the various metabolites formation at the initial growth stages as well as those under the different conditions of mineral nutrition is important for estimation of plant potential productivity and adaptation to environmental conditions. In this paper, we showed the regulatory effect of macro- and microelements on the growth and accumulation of primary and secondary metabolites in buckwheat plants. For the first time, the formation of primary and secondary metabolites in the aerial parts of a new and promising Russian buckwheat variety Dasha (approved by the State Register of the Russian Federation in 2018) has been characterized. The aim of this work was to study the initial stages of F. esculentum ontogenesis, including the assessment of the morphophysiological characteristics of seedlings under various mineral nutrition conditions, as well as the accumulation of photosynthetic pigments, sugars, and phenolic compounds in cotyledon leaves. Studies were conducted using two varieties of this culture included in the State Register of the Russian Federation in 2004 and 2018 (Devyatka and Dasha, respectively). Plant cultivation was carried out by a roll method in water (control) and Hoagland-Arnon nutrient medium (sample) at 24 °C and 16-hour illumination in laboratory conditions. In the seedlings, the height of the hypocotyls, the length of the roots, and the mass of cotyledon leaves was determined. The water content of the plant material was analyzed after it was dried to constant weight at 70 °C. The spectrophotometric method was used to determine the amount of chlorophylls a and b (λ = 665 nm and λ = 649 nm), carotenoids (λ = 440 nm), sugars (λ = 490 nm), the total amount of soluble phenolic compounds (λ = 725 nm), flavono-ids (λ = 415 nm) and phenylpropanoids (λ = 330 nm) in ethanol extracts from cotyledon leaves of seedlings of different ages. The cultivation of buckwheat on Hoagland-Arnon nutrient medium contributed to faster growth of aboveground organs compared to control; in contrast, the growth of underground organs was the same in both cases. In most cases, in the experimental samples, the differences in the accumulation of photosynthetic pigments (chlorophyll a and b, carotenoids) and soluble sugars in the cotyledons of two buckwheat varieties were revealed to be higher than in control. As for the accumulation of phenolic compounds, it was not obviously dependent on the level of mineral nutrition. As an exception, in cotyledons of seedlings cultivated on a nutrient medium, the content of phenylpropanoids changed to a greater extent compared to control and reached high values at the end of the investigation period. It should also be noted that on a nutrient medium at the late ontogenesis stages cotyledons of Dasha seedlings significantly accumulate pigments, sugars and phenolic compounds in comparison with Devyatka. Thus, the obtained data indicate that the amount of mineral elements is important for the initial stages of F. esculentum ontogenesis. Faster growth of seedlings and the accumulation of primary and secondary metabolites in their leaves is characteristic of the experimental samples, compared to the control. Therefore, in plant cultivation, the different levels of mineral nutrition make it possible to regulate the plant growth and development, as well as the accumulation of various metabolites.
Текст научной статьи Рост проростков гречихи обыкновенной (Fagopyrum esculentum Moench) и накопление первичных и вторичных метаболитов при различных условиях минерального питания
Формирование и развитие проростков представляет собой важный этап онтогенетического развития растений, зависящий от эндогенного запаса метаболитов семян и их превращений, а также воздействия экзогенных факторов окружающей среды, включая влажность почвы, температуру, свет, 946
минеральное питание (1, 2). Для этого периода характерен переход от гетеротрофного типа питания к автотрофному, изменение структурной организации клеток и тканей, энергетических процессов (дыхание, фотосинтез), состояния гормональной, антиоксидантной и других систем (3-5).
К важным регуляторам жизнеспособности растений относятся фенольные соединения — одни из наиболее распространенных вторичных метаболитов, которые присутствуют во всех клетках и тканях (6, 7). Их содержание зависит от видовой принадлежности растений, стадии онтогенеза, условий произрастания и минерального питания (8-10). Функциональная роль фенольных соединений чрезвычайно разнообразна и связана с процессами фотосинтеза, дыхания, роста и развития растений, а также c устойчивостью к стрессовым воздействиям (6, 11, 12).
Гречиху обыкновенную ( Fagopyrum esculentum Moench) причисляют к главным продовольственным культурам, она возделывается во многих странах мира и успешно используется в различных отраслях промышленности. Для гречихи характерно значительное накопление фенольных соединений, в том числе рутина — вещества с высокой капилляроукрепляющей активностью (13, 14). Наибольшее содержание этих вторичных метаболитов отмечено в надземных органах растений, особенно в листьях и цветках (15, 16). Сообщалось об образовании фенольных соединений в проростках, где их количество было меньше, а состав менее разнообразен по сравнению со взрослыми растениями (17, 18). Поскольку фенольные соединения обладают высокой биологической и антиоксидантной активностью, в том числе как потенциальные компоненты функционального питания (13), изучение их накопления на начальных этапах онтогенеза растений представляет практический интерес.
В настоящей работе, сравнив ряд морфологических, физиологических и биохимических показателей у двух сортов гречихи (Девятка и Даша) в условиях различного минерального питания, мы выявили регуляторное действие макро- и микроэлементов на процессы раннего онтогенеза и накопления первичных и вторичных метаболитов в надземных органах при определенной сортовой специфичности реакции растений. У нового перспективного сорта Даша, который в 2018 году внесен в Государственный реестр селекционных достижений, допущенных к применению в Российской Федерации, эти процессы охарактеризованы впервые.
Нашей целью была оценка особенностей начальных этапов онтогенеза, морфофизиологических характеристик и накопления в семядольных листьях фотосинтетических пигментов, сахаров и фенольных соединений у проростков гречихи обыкновенной в зависимости от обеспеченности элементами минерального питания.
Методика . Изученные сорта гречихи обыкновенной Девятка и Даша (19, 20) получены во Всероссийском НИИ зернобобовых культур и внесены соответственно в 2004 и 2018 году в Государственный реестр селекционных достижений, допущенных к использованию в Российской Федерации.
Проростки выращивали методом рулонной культуры (10). Для этого семена помещали в чашки Петри на фильтровальную бумагу, смоченную водой (контроль) или питательной средой Хогланда-Арнона (опыт) (21). После выдерживания в темноте в течение 24 ч их помещали в рулоны из фильтровальной бумаги (15 шт. в один рулон), которые ставили в пластиковые стаканы (по 7 рулонов в один стакан) с водой или питательной средой и выращивали в камере фитотрона ИФР РАН при 24 °С и 16часовом фотопериоде (5000 лк). Для исследования брали проростки, нахо- дящиеся на одной фазе онтогенетического развития: при выращивании на воде это были 11-е, 14-е и 18-е сут роста, при выращивании на питательной среде — 6-е, 11-е и 14-е сут (соответственно фазы 1-я, 2-я и 3-я). Критерием служили форма и размеры семядольных листьев, которые использовали для биохимических исследований.
Оценивали морфофизиологические параметры проростков — высоту надземной части и длину корней, а также массу семядольных листьев. Оводненность тканей определяли после высушивания растительного материала при 70 °С в термостате до постоянной массы (10).
Для извлечения пигментов листья проростков гомогенизировали в 96 % этаноле в темноте. Гомогенат центрифугировали (СМ-50, «ELMI Ltd.», Латвия) в течение 5 мин при 13000 об/мин. В надосадочной жидкости спектрофотометрическим методом (СФ-46, «ЛОМО», Россия) определяли количество хлорофиллов a и b (соответственно X = 665 нм и X = 649 нм), а также каротиноидов (X = 440 нм). Их содержание рассчитывали стандартным методом (22).
Сахара извлекали из растительного материала посредством экстракции 96 % этанолом (23). В надосадочной жидкости, полученной после центрифугирования гомогената (2 мин, 16000 об/мин), определяли суммарное содержание сахаров спектрофотометрическим методом по реакции с фенолом и серной кислотой (поглощение при X = 490 нм) (24). Калибровочную кривую строили по сахарозе.
Для экстракции фенольных соединений растительный материал гомогенизировали в 96 % этаноле и выдерживали при 45 °С в течение 45 мин (10, 24). Гомогенат центрифугировали (2 мин, 16000 об/мин). Надосадочную жидкость использовали для определения различных классов фенольных соединений спектрофотометрическим методом. Содержание суммы растворимых фенольных соединений оценивали с реактивом Фолина-Дениса (X = 725 нм), содержание флавоноидов — с 1 % водным раствором хлористого алюминия (X = 430 нм). Количество фенилпропаноидов определяли методом прямой спектрофотометрии растворов при X = 330 нм. Калибровочные кривые для расчета суммы фенольных соединений и флавоноидов строили по рутину, для расчета содержания фенилпропаноидов — по кофейной кислоте. В экспериментах использовали 3-кратные биологические и 2-кратные аналитические повторности измерений.
Дисперсионный анализ (ANOVA) проводили в программах SigmaPlot 12.3 и Microsoft Excel. В таблице и на графиках представлены средние арифметические значения полученных величин (M) и их стандартные ошибки (±SEM). Надстрочные символы обозначают достоверность различий средних значений по тесту Тьюки при р < 0,05.
Результаты . Важным показателем роста и развития растений служат их морфофизиологические характеристики, которые зависят от стадии онтогенеза, видовых и сортовых особенностей, а также действия внешних факторов, в том числе минерального питания (2, 4, 8).
Сорта гречихи Девятка и Даша — среднеспелые, высокоурожайные, устойчивые к полеганию (25, 26). При выведении сорта Даша отбор проводился на высокую озерненность и фотосинтетическую активность (20). Для этого сорта характерна более выраженная устойчивость к засухе, поражению аскохитозом и ложной мучнистой росой по сравнению с сортом Девятка (26). Следовательно, можно предположить некоторые различия в морфофизиологических характеристиках названных сортов, в том числе на ранних этапах онтогенеза, начиная с формирования и развития семядоль- ных листьев.
При выращивании в водной культуре (контроль) длина корней у проростков обоих сортов была практически одинаковой и увеличивалась в течение всего периода исследований (табл. 1). К 3-й фазе она возрастала на 57-60 % по сравнению с 1-й фазой. Это свидетельствует о значительном сходстве начальных этапов роста подземных органов у проростков двух сортов гречихи (1).
Формирование и развитие надземных органов обеспечивается как эндогенным запасом метаболитов в семенах, так и образованием новых метаболитов за счет процессов фотосинтеза (2, 27). В наших опытах высота гипокотилей у проростков сорта Девятка во все фазы онтогенеза достоверно превышала таковую у сорта Даша (в среднем на 16 %, р ≤ 0,05). У обоих сортов ее увеличение отмечалось при переходе ко 2-й фазе (на 40 % по сравнению с 1-й фазой), в дальнейшем высота гипокотилей не изменялась.
Полученные данные согласуются с показателями массы гипокотилей, которая у проростков сорта Девятка была достоверно (р ≤ 0,05) выше, чем у сорта Даша. При этом у сорта Девятка в первые две фазы онтогенеза она не изменялась, а к 3-й фазе увеличивалась на 19 % (р ≤ 0,05). У проростков сорта Даша масса гипокотилей не изменялась в течение всего периода роста. Эти результаты свидетельствуют о более быстром росте и накоплении биомассы в надземных органах у проростков традиционного сорта (Девятка) по сравнению с сортом следующего поколения (Даша).
Формирование и развитие листьев обеспечивает переход растений к автотрофному типу питания (1, 28). На всех этапах онтогенеза масса семядольных листьев у проростков обоих сортов гречихи была небольшой и практически равной. Единственным исключением оказались проростки сорта Девятка, у которых на 3-й фазе масса семядольных листьев была на 33 % выше (р ≤ 0,05) (см. табл. 1).
1. Морфофизиологическая характеристика проростков у двух сортов гречихи обыкновенной ( Fagopyrum esculentum Moench), выращенных в разных условиях, в зависимости от возраста ( M ±SEM, лабораторный опыт)
|
Фаза онтогенеза |
Длина корня, см |
Гипокотиль |
Масса семядольных листьев, г |
Оводненность семядольных листьев, % |
|
|
высота, см |
масса, г |
||||
|
Контроль |
(вода) |
||||
|
Сорт Девятка |
|||||
|
1-я |
7,08±0,30с |
9,85±0,41d |
0,13±0,02d |
0,04±0,004d |
89,53±1,62а |
|
2-я |
11,67±0,44b |
14,07±0,31b |
0,14±0,02d |
0,04±0,009d |
91,07±0,37а |
|
3-я |
12,04±0,80а |
14,32±1,12b |
0,16±0,02c |
0,06±0,006c |
91,05±0,16а |
|
Сорт Даша |
|||||
|
1-я |
7,42±0,32с |
8,27±0,46e |
0,11±0,01e |
0,04±0,003d |
88,77±0,69а |
|
2-я |
11,85±0,44b |
11,74±0,31c |
0,11±0,01e |
0,04±0,005d |
91,24±0,17а |
|
3-я |
13,00±1,00а |
11,97±1,07c |
0,12±0,01e |
0,04±0,004d |
91,43±1,45а |
|
Пита |
т е л ь н ая с р ед а Х о г л а нд а - Ар н о н а (опыт) |
||||
|
Сорт Девятка |
|||||
|
1-я |
7,62±0,44с |
9,11±0,87d |
0,15±0,01c |
0,06±0,006c |
88,28±0,11а |
|
2-я |
8,20±0,50с |
14,69±0,91b |
0,21±0,02b |
0,06±0,010c |
91,96±0,29а |
|
3-я |
11,02±0,26b |
17,97±0,21a |
0,27±0,02a |
0,08±0,013a |
93,50±0,60а |
|
Сорт Даша |
|||||
|
1-я |
7,72±0,38с |
4,75±0,50f |
0,14±0,02d |
0,05±0,005c |
88,79±0,50а |
|
2-я |
8,26±0,44с |
13,69±0,29b |
0,21±0,01b |
0,07±0,007b |
91,93±0,27а |
|
3-я |
11,24±0,32b |
17,21±0,59a |
0,27±0,02a |
0,07±0,013b |
95,48±1,58а |
|
П р и м е ч а н и е. Достоверные различия |
средних значений при р ≤ 0,05 отмечены неодинаковыми ла- |
||||
|
тинскими буквами. |
|||||
На питательной среде Хогланда-Арнона (опыт), то есть в условиях обеспеченности макро- и микроэлементами, развитие проростков ускорялось по сравнению с контролем. Так, 6-суточные проростки в опытных вариантах по морфофизиологическим характеристикам соответствовали 11-суточным проросткам в контроле, 11-суточные — 14-суточным,
14-суточные — 18-суточным.
Длина корня у проростков двух сортов не различалась на всех этапах исследования. При этом во 2-ю и 3-ю фазы она была меньше, чем в контроле (см. табл. 1). Прирост корня за весь период составил 30 %, то есть наличие макро- и микроэлементов в среде замедляло развитие подземных органов.
Высота гипокотилей у проростков сорта Девятка в 1-ю и 2-ю фазы онтогенеза была равной в опытном и контрольном вариантах, а в 3-ю фазу оказалась больше в опыте. У сорта Даша опытный вариант всегда достоверно (р ≤ 0,05) отличался от контроля, причем в 1-ю фазу значения были ниже, во 2-ю и 3-ю фазы — выше. Следует также отметить, что высота гипокотилей у проростков сорта Девятка в 1-ю фазу была на 50 % ниже, чем у проростков сорта Даша, а в дальнейшем — практически равной. При этом ее общее увеличение за весь период исследования у сорта Девятка составило 50 %, у сорта Даша — 72 %. Что касается массы гипокотилей, то она была почти одинаковой у обоих сортов и в течение периода роста проростков увеличивалась на 44 %. Ее значения во все фазы онтогенеза в опытном варианте превышали контроль.
Определение массы семядольных листьев у проростков двух сортов гречихи не выявило значительных различий между ними. В процессе онтогенеза она увеличилась у сорта Девятка на 25 %, у сорта Даша — на 28 %. В целом почти все показатели надземных органов у проростков в опытных вариантах, особенно на завершающем этапе исследования (3-я фаза), оказались достоверно (р ≤ 0,05) выше контрольных, что свидетельствует о стимулирующем действии питательного раствора.
Содержание воды в тканях — важный показатель при оценке физиологического состояния растений (1). Оводненность семядольных листьев у проростков двух сортов гречихи была одинаковой и повышалась в процессе онтогенетического развития (см. табл. 1). Наибольшие значения были отмечены в завершающую фазу роста. Обеспеченность проростков гречихи минеральным питанием не влияла на этот показатель.
2. Содержание пигментов в семядольных листьях у проростков двух сортов гречихи обыкновенной ( Fagopyrum esculentum Moench), выращенных в разных условиях, в зависимости от возраста ( M ±SD)
|
Фаза |
Хлорофиллы, мг/г сухой массы |
Хлорофиллы, а/b |
Каротиноиды, |
|
онтогенеза |
а 1 b 1 а + b |
мг/г сухой массы |
|
Ко нтр оль (вода) Сорт Девятка |
|||||
|
1-я |
5,29±0,28d |
1,24±0,12e |
6,53±0,40d |
4,26 |
0,81±0,04d |
|
2-я |
6,19±0,05c |
1,51±0,04d |
7,70±0,09с |
4,09 |
0,98±0,03c |
|
3-я |
5,67±0,87cd |
1,48±0,26d |
7,15±1,13с Сорт Даша |
3,83 |
0,49±0,05f |
|
1-я |
5,14±0,18d |
1,30±0,11e |
6,44±0,29d |
3,95 |
0,71±0,10e |
|
2-я |
5,25±0,42d |
1,38±0,12de |
6,63±0,54d |
3,80 |
0,96±0,09c |
|
3-я |
5,14±0,68d |
1,39±0,04de Питат |
6,53±0,72d ельная среда Сорт Девятка |
3,70 (опыт) |
0,52±0,19f |
|
1-я |
2,28±0,02f |
3,51±0,05a |
5,79±0,07e |
0,01 |
|
|
2-я |
5,44±0,18d |
1,49±0,16d |
6,93±0,34d |
3,65 |
0,50±0,04f |
|
3-я |
7,25±0,03b |
1,90±0,15c |
9,15±0,18в Сорт Даша |
3,81 |
1,23±0,01b |
|
1-я |
3,19±0,09e |
3,78±0,26a |
6,97±0,35d |
0,84 |
|
|
2-я |
6,22±0,53c |
1,65±0,12d |
7,87±0,65с |
3,76 |
0,65±0,08e |
|
3-я |
9,09±0,61a |
2,39±0,17b |
11,48±0,70а |
3,80 |
1,45±0,04a |
П р и м еч а ни е. Достоверные различия средних значений при р ≤ 0,05 отмечены неодинаковыми латинскими буквами. Прочерки означают, то каротиноиды выявляли, начина со 2-й фазы онтогенеза.
Фотосинтез растений — главный биологический процесс, обеспечивающий жизнь всех организмов на планете (27). Для оценки его эффек- тивности используют такие показатели, как содержание хлорофиллов a и b в листьях и их соотношение (28). В контрольных условиях нашего опыта были выявлены различия в накоплении хлорофиллов a и b в семядольных листьях по мере роста проростков обоих сортов гречихи (табл. 2). У сорта Девятка содержание хлорофилла a в 1-ю фазу оказалось наименьшим, во 2-ю фазу достоверно повышалось на 17 % (р ≤ 0,05), а к 3-й фазе снижалось на 10 %, но превышало показатель в 1-ю фазу. Аналогичная, но менее выраженная тенденция проявлялась и в отношении хлорофилла b. В семядольных листьях у проростков сорта Даша содержание хлорофиллов a и b в течение всего периода исследования не изменялось и было практически равным таковому у проростков сорта Девятка в 1-ю фазу.
При оценке фотосинтетической продуктивности растительных тканей важно учитывать отношение хлорофиллов a/b, которое при оптимальных условиях роста приближается к 3 (27, 28). Для семядольных листьев проростков обоих сортов гречихи в контроле отмечали более высокие показатели отношения хлорофиллов a/b, причем в большей степени это было характерно для 1-й и 2-й фаз (см. табл. 2).
Пигментная система растений, помимо хлорофиллов, содержит каротиноиды, которые участвуют в функционировании реакционных центров и светособирающих комплексов фотосистем хлоропластов, поглощают свет в синей области спектра, защищают фотосинтетический аппарат от фотодеструкции, а также выполняют другие защитные функции (29, 30). У обоих сортов гречихи наблюдались одинаковые тенденции в накоплении каротиноидов: высокое содержание в 1-ю фазу, последующее увеличение во 2-ю фазу (примерно на 20 %) и значительное снижение в 3-ю фазу (почти в 2 раза). Следовательно, начальные этапы формирования и развития семядольных листьев у проростков гречихи при низком уровне минерального питания характеризуются значительным накоплением каротиноидов, что может свидетельствовать об их важной роли в этот период онтогенетического развития (31).
Изучение накопления пигментов в семядольных листьях у проростков гречихи, выращенных на питательной среде, выявило несколько иные тенденции. Содержание хлорофиллов a и b в них достоверно (р < 0,05) повышалось в течение всего периода наблюдений, что не было характерно для контрольных вариантов (см. табл. 2). В 1-ю фазу количество хлорофилла a в семядольных листьях у проростков сортов Девятка и Даша было минимальным (соответственно в 2,3 и 1,6 раза ниже, чем в контроле, р ≤ 0,05). Содержание хлорофилла b в эту фазу оказалось наибольшим и превышало контрольный показатель почти в 3 раза (р ≤ 0,05). Последующее развитие семядольных листьев (2-я и 3-я фазы) сопровождалось значительным (р ≤ 0,05) повышением в них содержания хлорофиллов a и b, что в большей степени проявлялось у сорта Даша. При его селекции проводился отбор на фотосинтетическую продуктивность растений (27), и эта особенность проявлялась уже на ранних этапах их развития. Следует также отметить, что суммарное содержание хлорофиллов a и b в семядольных листьях в опытных вариантах за весь период исследования достоверно (р ≤ 0,05) повысилось: у сорта Девятка на 58 %, у сорта Даша — на 64 %.
Что касается отношения хлорофиллов a/b в семядольных листьях, то в 1-ю фазу оно было низким (0,06 и 0,80 соответственно у сорта Девятка и сорта Даша), а во 2-ю и 3-ю фазы значительно повышалось и становилось практически равным у обоих сортов (в среднем 3,76). Эти показатели соответствовали контрольным, то есть наблюдалось значительное сходство в формировании фотосинтетического аппарата в семядольных ния хлоропластов — важных источников энергии
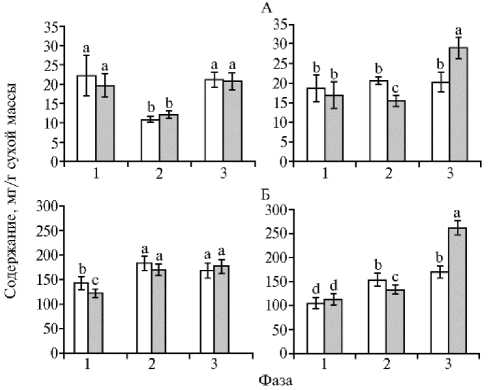
Рис. 1. Содержание сахаров (А) и фенольных соединений (Б) в семядольных листьях у проростков гречихи обыкновенной ( Fagopyrum esculentum Moench) сортов Девятка (белые столбики) и Даша (серые столбики) , выращенных на воде (слева) и питательной среде Хогланда-Арнона (справа) , в разные фазы онтогенеза. Достоверные различия средних значений при р ≤ 0,05 отмечены неодинаковыми латинскими буквами над барами.
листьях гречихи в более поздние фазы онтогенеза, которое не зависело от минерального питания проростков (см. табл. 2).
В опытных вариантах накопление каротиноидов в семядольных листьях было практически равным у проростков обоих сортов гречихи (см. табл. 2). Его отмечали, начиная со 2-й фазы, но оно было ниже, чем в контроле, а к 3-й фазе увеличивалось почти в 2,5 раза (р ≤ 0,05) и значительно превышало контроль.
В целом, наличие питательных элементов способствовало более эффективному образованию хлорофиллов и каротиноидов, что было следствием быстрого роста растений, развития семядольных листьев и формирова-метаболитов (27, 28).
Известно, что на начальных этапах онтогенеза растения испытывают существенную потребность в энергии и метаболитах для роста и накопления биомассы (1, 2). Растворимые сахара становятся для них основной транспортной формой ассимилятов и могут служить исходными субстратами для многих процессов метаболизма, а также образования структурных элементов клеток и тканей, что необходимо для развития проростков (32).
Накопления растворимых сахаров в семядольных листьях в контрольных вариантах было практически рав ным у проростков двух сортов гречихи в течение трех фаз онтогенеза (рис. 1, А). В 1-ю фазу количество сахаров было высоким, во 2-ю фазу — достоверно (р ≤ 0,05) снижалось (на 51 %), в 3-ю фазу — повышалось (р ≤ 0,05), достигая исходных значений. Эти различия были обусловлены обеспеченностью исходными субстратами для образования растворимых сахаров, а именно запасными веществами в семенах гречихи (1-я фаза), их последующим истощением и активным ростом проростков (2-я фаза) и, наконец, фотосинтезом (3-я фаза), приводящим к повышению количества метаболитов и накоплению растворимых сахаров (23, 32).
Для накопления растворимых сахаров в семядольных листьях проростков, выращенных на питательной среде, наблюдалась иная тенденция (см. рис. 1, А). У сорта Девятка на всех этапах исследования содержание растворимых сахаров было одинаковым и достаточно приближенным к значениям в контроле в 1-ю и 3-ю фазы. У сорта Даша в 1-ю фазу оно оказалось равным таковому у сорта Девятка, во 2-ю фазу — снижалось на 25 % (р ≤ 0,05), в 3-ю — фазу возрастало на 47 % (р ≤ 0,05). Такая закономерность характерна только для сорта Даша, который был создан селекционерами посредством отбора по интенсивности фотосинтетической активности (20, 26).
Фенольные соединения — одни из важнейших метаболитов растений, роль которых, как и химическая структура, чрезвычайно разнообразна (6, 7). При выращивании в контрольных условиях в 1-ю фазу у сорта Девятка количество фенольных соединений в семядольных листьях было на 13 % выше, чем у сорта Даша (см. рис. 1, Б). Во 2-ю и 3-ю фазы накопление фенольных соединений достоверно (р ≤ 0,05) увеличивалось у сортов Девятка и Даша соответственно на 22 и 28 % и становилось одинаковым. В определенной степени это могло быть следствием равной фотосинтетической активности семядольных листьев в указанный период онтогенеза, о чем свидетельствует содержание в них фотосинтетических пигментов (см. табл. 2). Известно, что хлоропласты — одно из основных мест биосинтеза фенольных соединений в клетках зеленых растений (33).
При выращивании на питательной среде у проростков обоих сортов гречихи суммарное содержание фенольных соединений в семядольных листьях в большинстве случаев было достоверно (р ≤ 0,05) ниже, чем в контроле (см. рис. 1, Б). При этом у сорта Девятка наименьшее значение показателя отмечали в 1-ю фазу, ко 2-й фазе оно повышалось на 32 % (р ≤ 0,05) и оставалось таким до 3-й фазы, как и в контрольном варианте. У сорта Даша в 1-ю фазу количество фенольных соединений также было самым низким и не отличалось от показателя у сорта Девятка. Ко 2-й фазе оно достоверно повышалось на 15 % (р ≤ 0,05), а к 3-й фазе уве- значения.
личивалось в 2 раза и достигало максимального
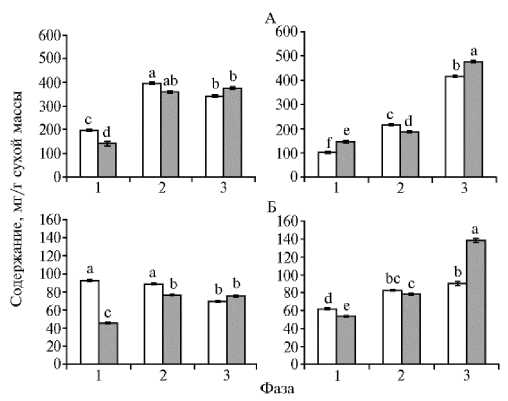
Рис. 2. Содержание фенилпропаноидов (А) и флавоноидов (Б) в семядольных листьях у проростков гречихи обыкновенной ( Fagopyrum esculentum Moench) сортов Девятка (белые столбики) и Даша (серые столбики) , выращенных на воде (слева) и питательной среде Хогланда-Арнона (справа) , в разные фазы онтогенеза. Достоверные различия средних значений при р ≤ 0,05 отмечены неодинаковыми латинскими буквами над барами.
Как уже отмечалось, фенольные соединения в растениях чрезвычайно разнообразны по структуре и представлены различными классами (7). К наиболее простым из них относятся фенилпропаноиды (6). В семядольных листьях у проростков гречихи, выращенных в контрольных условиях, в 1-ю фазу содержание фенил-пропаноидов было наименьшим, что в большей степени проявлялось у сорта Даша (рис. 2, А). Ко 2-й фазе оно достоверно (р ≤ 0,05) возрастало (у сортов Девятка и Даша соответственно на 50 и 60 %), а в дальнейшем (3-я фаза) уменьша- лось у сорта Девятка на 14 % (р ≤ 0,05) и не изменялось у сорта Даша. Результатом этого стало одинаковое содержание фенилпропаноидов в семядольных листьях у проростков обоих сортов гречихи в конце периода ис- следования.
При достаточном минеральном питании накопление фенилпро-паноидов в семядольных листьях в 1-ю фазу и особенно во 2-ю фазу оказалось ниже по сравнению с контролем, а в 3-ю фазу превышало его (см.
рис. 2, А). В 1-ю фазу этот показатель был самым низким, что в большей степени проявлялось у сорта Девятка. Ко 2-й фазе он достоверно (р ≤ 0,05) повышался (у сорта Девятка и Даша соответственно на 53 и 22 %). Наибольшие изменения в содержании фенилпропаноидов отмечали в 3-ю фазу, когда их значения резко возрастали (в среднем на 50 %, р ≤ 0,05) и становились достаточно близкими к таковым в контрольных вариантах.
Известно, что для гречихи характерно образование флавоноидов — наиболее распространенных представителей фенольных соединений в надземных органах растений (6, 16). При выращивании проростков на воде в 1-ю фазу онтогенеза содержание флавоноидов в семядольных листьях у сорта Девятка почти в 2 раза превышало значение у сорта Даша (см. рис. 2, Б). Во 2-ю фазу оно не изменялось, а в 3-ю фазу снижалось на 22 % (р ≤ 0,05). У сорта Даша наблюдалась иная тенденция: содержание флавоноидов повышалось ко 2-й фазе на 41 % (р ≤ 0,05) и в дальнейшем не изменялось.
При выращивании проростков гречихи на питательной среде содержание флавоноидов в семядольных листьях у обоих сортов отличалось от контрольных значений (см. рис. 2, Б). При этом у сорта Девятка оно повышалось на 25 % (р ≤ 0,05) в 1-ю и 2-ю фазы, а затем не изменялось, тогда как у сорта Даша увеличение наблюдалось в течение всего периода исследования (1-ю и 2-ю фазы — на 32 %, 3-я фаза — на 43 %; р ≤ 0,05).
Все вышеизложенное свидетельствует о том, что условия минерального питания оказывают значительное влияние на начальные этапы онтогенеза растений гречихи обыкновенной. При выращивании на питательной среде Хогланда-Арнона скорость роста надземных органов и накопление пигментов (хлорофиллов а и b, а также каротиноидов) в семядольных листьях было выше по сравнению с контролем, что в большей степени проявлялось в конце периода исследования (3-я фаза онтогенеза). В литературе имеются данные о положительном влиянии минерального питания на прирост биомассы растений и содержание различных форм хлорофилла в листьях растений (22, 34, 35). Что касается отношения хлорофиллы a/b, то этот показатель не зависел от условий минерального питания.
Сахара — промежуточный продукт фотосинтеза (28). Между их накоплением и содержанием хлорофиллов достаточно часто отмечалась положительная корреляция (28, 32). Однако в случае проростков двух сортов гречихи четко выраженной тенденции не наблюдалось. Суммарное содержание фенольных соединений в семядольных листьях в контроле было выше по сравнению с опытом. Следовательно, в условиях лучшей обеспеченности проростков гречихи минеральными элементами количество вторичных метаболитов фенольной природы в надземных органах снижалось, что может быть следствием их интенсивного роста. Об уменьшении накопления полифенолов при активация роста растений сообщалось в литературе (6, 9). В то же время содержание отдельных их классов (фенилпропаноидов и флавоноидов) в семядольных листьях у проростков сорта Даша, выращенных на питательной среде, как правило, повышалось, а у проростков сорта Девятка — снижалось, за исключением 3-й фазы роста, где прослеживалась противоположная тенденция. Изменения в биосинтезе определенных классов фенольных соединений в проростках гречихи обыкновенной при различных условиях минерального питания требуют дальнейших исследований.
Таким образом, начальные этапы онтогенетического развития проростков двух сортов гречихи обыкновенной определяются запасом пита-954
тельных веществ в семенах и наличием микро- и макроэлементов. Внесение последних ускоряет рост надземных органов (в среднем на 20-30 %), развитие семядольных листьев (на 25 % для сорта Девятка и на 42 % для сорта Даша) и повышает фотосинтетическую активность по сравнению с аналогичными показателями у проростков, при выращивании которых питательную среду заменяли на воду. Все это отражается на содержании пигментов, накоплении первичных (сахара) и вторичных (различные классы фенольных соединений) метаболитов. Следовательно, изменяя обеспеченность элементами минерального питания при выращивании растений, можно регулировать их рост и развитие, а также накопление метаболитов.
Список литературы Рост проростков гречихи обыкновенной (Fagopyrum esculentum Moench) и накопление первичных и вторичных метаболитов при различных условиях минерального питания
- Hopkins W.G., Hüner N.P.A. Introduction to plant physiology. 4rd Edition. John Wiley & Sons, Inc., NY, 2008.
- Bouman F., Boesewinkel F.D. The seed: structure and function. In: Seed development and germination /K. Jaime, G. Gad (eds.). CRC Press, 2017.
- Chauhan B.S., Johnson D.E. Influence of environmental factors on seed germination and seedling emergence of eclipta (Eclipta prostrata) in a tropical environment. Weed Science, 2008, 56(3): 383-388 ( ). DOI: 10.1614/WS-07-154.1
- Алиева З.М., Самедова Н.Х., Юсуфов А.Г. Реакция растений на стрессы на начальных этапах онтогенеза. Аридные экосистемы, 2013, 19(1): 54.
- Baxter A., Mittler R., Suzuki N. ROS as key players in plant stress signalling. Journal of Experimental Botany, 2013, 65(5): 1229-1240 ( ). DOI: 10.1093/jxb/ert375
- Запрометов М.Н. Фенольные соединения. Распространение, метаболизм и функции в растениях. М., 1993.
- Cheynier V., Comte G., Davies K.M., Lattanzio V., Martens S. Plant phenolics: recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiology and Biochemistry, 2013, 72: 1-20 ( ).
- DOI: 10.1016/j.plaphy.2013.05.009
- Nannipieri P., Badalucco L. Biological processes. In: Handbook of processes and modelling in the soil-plant system /D.K. Benbi, R. Nieder (eds.). Haworth, Binghamton, 2003.
- Волынец А.П. Фенольные соединения в жизнедеятельности растений. Минск, 2013.
- Казанцева В.В., Гончарук Е.А., Фесенко А.Н., Широкова А.В., Загоскина Н.В. Особенности образования фенольных соединений в проростках гречихи (Fagopyrum esculentum Moench) различных сортов. Сельскохозяйственная биология, 2015, 50(5): 611-619 ( ).
- DOI: 10.15389/agrobiology.2015.5.611rus
- Mierziak J., Kostyn K., Kulma A. Flavonoids as important molecules of plant interactions with the environment. molecules, 2014, 19(10): 16240-16265 ( ).
- DOI: 10.3390/molecules191016240
- Kumar V., Suman U., Rubal, Yadav S.K. Flavonoid secondary metabolite: biosynthesis and role in growth and development in plants. In: Recent trends and techniques in plant metabolic engineering /S. Yadav, V. Kumar, S. Singh (eds.). Springer, Singapore, 2018: 19-45 ( ).
- DOI: 10.1007/978-981-13-2251-8_2
- Kreft M. Buckwheat phenolic metabolites in health and disease. Nutrition Research Reviews, 2016, 29(1): 30-39 ( ).
- DOI: 10.1017/s0954422415000190
- Suzuki T., Morishita T., Kim S.J., Park S.U., Woo S.H., Noda T., Takigawa S. Physiological roles of rutin in the buckwheat plant. Japan Agricultural Research Quarterly: JARQ, 2015, 49(1): 37-43 ( ).
- DOI: 10.6090/jarq.49.37
- Высочина Г.И. Фенольные соединения в систематике и филогении семейства гречишных. Новосибирск, 2004.
- Campbell C.G. Buckwheat: Fagopyrum esculentum Moench. Research Ltd., Morden, Mantitoba, 1997.
- Koyama M., Nakamura C., Nakamura K. Changes in phenols contents from buckwheat sprouts during growth stage. Journal of Food Science and Technology, 2013, 50(1): 86-93 ( ).
- DOI: 10.1007/s13197-011-0316-1
- Nam T.G., Lee S.M., Park J.H., Kim D.O., Baek N.I., Eom S.H. Flavonoid analysis of buckwheat sprouts. Food Chemistry, 2015, 170: 97-101 ( ).
- DOI: 10.1016/j.foodchem.2014.08.067
- Сорт Девятка®. Режим доступа: https://vniizbk.ru/ru/2017-01-23-11-23-52.html. Дата об-ращения: 21.10.2019.
- Сорт Даша®. Режим доступа: https://vniizbk.ru/newvarieties.html#dasha. Дата обращения 21.10.2019.
- Hoagland D.R., Arnon D.I. The water-culture method for growing plants without soil. In: Circular. California agricultural experiment station. University of California, Davis Libraries, 1950.
- Шлык А.А. Определение хлорофиллов и каротиноидов в экстрактах зеленых листьев. В сб.: Биохимические методы в физиологии растений. М., 1971: 154-170.
- Olenichenko N.A., Zagoskina N.V., Astakhova N.V., Trunova T.I., Kuznetsov Y.V. Primary and secondary metabolism of winter wheat under cold hardening and treatment with antioxidants. Applied Biochemistry and Microbiology, 2008, 44(5): 589-594 ( ).
- DOI: 10.1134/S0003683808050141
- DuBois M., Gilles K.A., Hamilton J.K., Rebers P.T., Smith F. Colorimetric method for determination of sugars and related substances. Analytical Chemistry, 1956, 28(3): 350-356 ( ).
- DOI: 10.1021/ac60111a017
- Мартыненко Г.Е., Фесенко Н.В., Фесенко А.Н., Гуринович И.А. Создание холодостойкого детерминантного сорта гречихи Девятка. Вестник аграрной науки, 2010, 25(4): 85-87.
- Фесенко А.Н., Амелин А.В., Фесенко И.Н., Бирюкова О.В., Заикин В.В. Новый сорт гречихи Даша. Земледелие, 2018, 4: 36-38 ( ).
- DOI: 10.24411/0044-3913-2018-10411
- Мокроносов А.Т., Гавриленко В.Ф., Жигалова Т.В. Фотосинтез. Физиолого-экологические и биохимические аспекты. М., 2006.
- Андрианова Ю.Е., Тарчевский И.А. Хлорофилл и продуктивность растений. М., 2000.
- Nisar N., Li L., Lu S., Khin N.C., Pogson B.J. Carotenoid metabolism in plants. Molecular Plant, 2015, 8(1): 68-82 ( ).
- DOI: 10.1016/j.molp.2014.12.007
- Sun T., Yuan H., Cao H., Yazdani M., Tadmor Y., Li L. Carotenoid metabolism in plants: the role of plastids. Molecular Plant, 2018, 11(1): 58-74 ( ).
- DOI: 10.1016/j.molp.2017.09.010
- Stange C. Carotenoids in nature: biosynthesis, regulation and function. Springer International Publishing, Switzerland, 2016 ( ).
- DOI: 10.1007/978-3-319-39126-7
- Eveland A.L., Jackson D.P. Sugars, signaling and plant development. Journal of Experimental Botany, 2011, 63(9): 3367-3377 ( ).
- DOI: 10.1093/jxb/err379
- Zaprometov M.N., Nikolaeva T.N. Chloroplasts isolated from kidney bean leaves are capable of phenolic compound biosynthesis. Russian Journal of Plant Physiology, 2003, 50(5): 623-626 (doi: 10.1023/A:1025683922953).
- Нагорный В.Д., Арималала Р.Н. Влияние серы на содержание пигментов в листьях и накопление сухого вещества растениями картофеля в условиях вегетационного опыта. Вестник Российского университета дружбы народов. Серия: Агрономия и животноводство, 2016, 3: 7-14.
- Sharafzadeh S., Khosh-Khui M., Javidnia K. Effect of nutrients on essential oil components, pigments and total phenolic content of lemon balm (Melissa officinalis L.). Advances in Environmental Biology, 2011, 5(4): 639-647.
