Рубки ухода: старые и новые теории

Автор: Рогозин М.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 3, 2015 года.
Бесплатный доступ
Проведены анализ литературы по вопросам горизонтальной структуры, уходу и выращиванию лесов и определение путей поиска новых факторов, влияющих на продуктивность древо-стоев. Рассмотрена старая теоретическая основа рубок ухода в виде стремления к равномерному размещению и оптимизации площади питания деревьев. Описан безуспешный опыт по ее верификации. Показано, что существует закон развития простых одноярусных древостоев, в соответствии с которым развитие фитоценоза четко делится на два периода: прогресс и регресс. Для них нужен разный тип управления древостоем: активный - в период прогресса и пассивный - в период регресса, что не учитывает современная теория лесоводства. Отмечено, что в моделях развития древостоев ели с большой начальной густотой прогресс заканчивается в 20-25 лет, тогда как в изначально редких это происходит в 40 лет. По этим причинам ныне применяемые в 30-40 лет прочистки в густых древостоях безнадежно опаздывают и почти бесполезны, а прореживания и проходные рубки не имеют теоретической основы, фактически разрушая древостой, и поэтому должны быть либо запрещены, либо кардинально изменены. Наличие прогалов и биогрупп, в которых формируется 28-57% деревьев в самых разных насаждениях, является атрибутом древостоя, и его необходимо учитывать в лесоводстве, т.е. биогруппы сохранять, а не разреживать. Их размещение, а также образование прогалов не случайно и может быть обусловлено взаимодействиями в системе Земля-растение в геоактивных зонах и геобиологических сетях (ГБС). По данным автора, прогалы приурочены к дискомфортным зонам этих сетей с размерами от 0.55 до 2.0 м, занимающими около 2% территории, на которых ель достоверно снижала рост в высоту на 14%. Скопления деревьев приходятся на зоны, благоприятные для их роста; они имеют размеры от 1.0 до 8.0 м и занимают 25% территории. Биогруппы, а также самые крупные деревья в насаждениях указывают на места таких зон. Однако наличие ГБС подвергают сомнению на основании того, что их выделяют биолокацией. Тем не менее, патогенные сети уже определяют техническими приборами и объективными методами контроля, фиксируя излучения ГБС на фотоматериалы, и они полностью подтверждают данные биолокации. В поисках причин неравномерного размещения деревьев нужны новые гипотезы, которые следует проверять, какими бы невероятными они не казались.
Лесоводство, рубки ухода за лесом, древостой, горизонтальная структура, биогруппы, геоактивные зоны
Короткий адрес: https://sciup.org/147204726
IDR: 147204726 | УДК: 582.47:
Thinning: old and new theories
The analysis of the literature on horizontal patterns, the care and cultivation of forests and identify ways to find new factors affecting the productivity of forest stands. Considered old theoretical basis of thinning in the form of a desire for uniform placement and optimization of the feeding area of trees. Describes an unsuccessful experience for its verification. It is shown that there is a law of development of simple single-tiered stands, according to which the development of phytocenosis is clearly divided into two periods: progress and regress. Each requires a different type of forest management: active - a period of progress and passive - in the period of regression that ignores the modern theory of silviculture. It is noted that in the models of development of spruce forests with a large initial density of progress ends in 20-25 years, whereas in initially rare it happens in 40 years. For these reasons, the current in 30-40 years clearing in dense forest stands is hopelessly late and almost useless, and decimation and through cuttings do not have a theoretical basis, effectively destroying the forest and therefore should either be banned or radically changed. The presence of gaps and displays biological groups, which is formed in 28-57% of the trees in different plantations, is an attribute of the forest, and it must be considered in forestry, i.e. biological groups to maintain and not retrieving. Their placement, as well as the formation of gaps is not accidental and may be due to interactions in the system Earth-a plant in geoactive zones and geobiological networks (GBS). According to the author, the gaps are confined to the uncomfortable zones of these networks with sizes ranging from 0.55 to 2.0 m, which occupies about 2% of the area on which the spruce was significantly reduced growth in height by 14%. Clumps of trees occur in areas favorable for their growth; they have dimensions of from 1.0 to 8.0 m and is 25% of the territory. Biological groups, as well as the largest trees in the stands indicate the location of such zones. However, the presence of GBS questioned on the basis that they are biolocation. However, pathogenic network devices determine the technical and objective methods of control, locking the radiation of GBS on the photographs, and they fully confirmed by the evidence of dowsing. In search of reasons for the uneven placement of trees new hypotheses that should be checked, no matter how improbable they may seem.
Текст научной статьи Рубки ухода: старые и новые теории
В лесоводстве проводят рубки ухода, известные под названиями «прочистки», «прореживания» и «проходные рубки». Теория этих рубок основана на увеличении площади питания у оставляемых деревьев с целью повышения прироста древостоя. Их интенсивность до 1980 г. не превышала 10-20% запаса, убирали в основном подчиненную часть древостоя. Но в последующем их рекомендуемая интенсивность возросла вдвое, и площадь только технологических коридоров (по сути, широких просек) допускается ныне уже в 15% от площади лесосеки; при этом общая интенсивность принимается 25-30% [Правила..., 2007]. Такого рода рубки называют также коммерческими. Возникает проблема верификации достижения лесово-детвенных целей такими рубками.
И здесь возникает просто вакуум — такой верификации в лесоводственной практике просто нет. Есть много эмоциональных высказываний о нарушениях технологии, но нет сводки их отдаленных результатов. Между' тем уже достаточно давно были получены результаты опытных рубок [Сеннов. 1984], и результаты просто обескуражили ученых-лесоводов. Подводя их итоги после 60 лет наблюдений, было выяснено, что увеличение площади питания дерева в среднем возрасте путем удаления конкурирующих с ним соседей отнюдь не увеличивает их прирост, как это полагалось по теории; далее этот вывод был подтвержден и в одном из учебников [Сеннов. 2005]. Получается, что деревья «не слушаются» лесовода и прирост не увеличивают, несмотря на декларируемые в теории рубок ухода пожелания увеличить его на оставляемых деревьях. Более того, рубки эти на практике имеют сейчас совершенно другие цели: они попросту изымают будущие запасы спелой древесины задолго до полноценной спелости древостоев.
В работе проведен краткий анализ литературы по вопросам горизонтальной структуры, ухода и выращивания лесов, включая работы автора, а также определение путей поиска новых факторов, влияющих на продуктивность лесов.
Теория рубок ухода
Знакомство с некоторыми современными учебниками по лесоводству и лесной таксации [Набатов, 2002: Сеннов. 2005: Верху нов. Черных, 2007; Калинин, 2011] показало, что их авторы, рассматривая теоретические основы рубок ухода, всегда ссылаются на классификацию деревьев немецкого лесовода Крафта. который в 1884 г. предложил поделить взрослые деревья на 5 классов по степени развития кроны. С некоторыми дополнениями она используется и сейчас [Желдак, Атрохин, 2003]. Основным методом ухода за чистыми и одновозрастными древостоями при такой классификации считается «низовой метод», при котором удаляют усыхающие, отставшие в росте и искривленные деревья. При этом авторы учебников, соблюдая традиции еще XIX в., отодвигают время начала регулирования текущей густоты ценоза к возрасту деревьев в 40-50 лет, так как именно к этому моменту наступает «дифференциация» деревьев на указанные классы Крафта. В хвойных молодняках в 21-40 лет такие рубки называли «прочистками», а после 40 лет - «прореживаниями». Примерно до 1980 г. при таких рубках в массовом порядке заготавливали мелкую деловую древесину7 (в основном колья и жерди), Но затем спрос упал почти до нуля. и взоры лесников обратились к древесине средней крупности, которую МОЖНО было получить только после 40—50 лет. И в правилах рубок ухода, принятых в те годы, это нашло свое отражение усилением интенсивности прореживаний и проходных рубок в 2 раза - до 25-30%.
При этом, как ни странно, их теоретическая основа осталась прежней. Появились новые знания о развитии леса (прошло более 100 лег после публикации работ Крафта), и их нужно было интегрировать в лесоводство и в соответствующие правила, по которым предписывают ухаживать за лесом. Однако этого нс произошло. Так, в перечисленных учебниках мы не встретили упоминаний и ссылок на «основную закономерность естественного морфогенеза древостоев» Г. С. Разина [Разин, 1979]. а также на «ранговый закон роста деревьев» Е.Л. Маслакова [Маслаков, 1981. 19S4J, к моменту выхода этих учебников известных уже четверть века. Получается, что теория рубок ухода осталась на уровне XIX в. Мало того, и некоторые моменты плантационного выращивания леса (новая технология с изначально редким размещением деревьев и отбором лидеров в раннем возрасте) совершенно в них не рассматриваются и не находят в теории рубок ухода своего воплощения. В результате получаются два лесоводства - «классическое» и «плантационное». В итоге получаем кризис теории лесоводства вообще.
Почему7 же это произошло? Рассматривая теоретическую основу рубок ухода, мы обнаружили, что ее можно свести к одной простой и в чем-то механистической идее: прирост биомассы дерева определяется его площадью питания, и чем она больше, тем выше должен быть прирост дерева. Эго положение много лет казалось настолько очевидным и бесспорным, чю нс возникало даже мысли проверить его истинность в длительных опытах на разных объектах. Например, в одном варианте изреживать молодые древостои в 10-15 лет, в другом - 20-30-летние, а в третьем - 40-50-летние.
При анализе литературы по моделированию хода роста и развития древостоев [Рогозин, Разин, 2015]. мы были удивлены тем, что из множества работ по вопросам выращивания леса нам встретились работы лишь одного автора [Сеннов, 1984, 1999, 2005], где теория рубок ухода была верифицирована. В его работах было доказано, что увеличение площади питания у дерева в возрасте начиная с 40 лет удалением соседствующих деревьев достоверно не увеличивает прирост и продуктивность древостоя. Отметим, что опытов, начатых в более молодых ценозах (15-30-летних), у С.Н. Сеннова не было. Таким образом, казавшаяся логически безупречной идея повышения проективности древостоев рубками ухода в возрасте старше 40 лет не подтвердилась строгими опытами. Из этого следует. что какие-то общие закономерности в развитии леса классическое лесоводство упускало из виду*
Анализируя историю лесоводства, мы обнаружили в чем-то знаковую работу «Рубки ухода за лесом в новом освещении» Г.Р. Эйтингена [1934]. Его идеи в свое время были передовыми и вдохновили поколения лесоводов на применение более интенсивных рубок ухода; на нее ссылаются и сейчас.
Суть ее состояла в следующем. На 8 участках культур сосны и ели в возрасте от 15 до 38 лет автор срубил 4 тыс, деревьев и измерял у них все приросты по высоте (более 70 тыс ). Это были поистине феноменальные по объему7 исследования. На каждое дерево была заведена карточка, и по динамике прироста деревья разделили на 3 типа и 2^1 подтипа (всего 10). В те годы статистика уже была известна, и Г.Р. Эйгинген использовал ее, но анализ точечных диаграмм (полей автокорреляции). как это делается сейчас [Рогозин, Разин, 2015] тогда еще не применяли. Поэтому выяснение вероятностей формирования деревьев-лидеров из деревцев разного размера было заменено Г.Р. Эй-тингеном делением деревьев на некие заранее указанные типы. В итоге результаты анализа свелись к тому, что в молодости устойчиво-слабый и устойчиво-сильный прирост имели около 90%, а неустойчивый - около 10% деревьев. Далее автор выяснил, что численность последней группы деревьев с неустойчивым приростом ближе к 40-летнему возрасту почти удваивалась.
На этом основании и в результате описания разнообразной динамики прироста у деревьев за 12-35 лег, а также акцента на его постоянные изменения с возрастом от дерева к дереву, автор предложил отказаться « .от консервативных способов рубок у^ода ПОЛСТОЛСТИЯ господствовавших Б ЛССОБОДСТВСННОЙ литературе, ИСХОДИВШИХ из представлений о необходимости огульной вырубки всех отставших в росте деревьев.. ». Далее ГР. Эйгинген выдвинул новое положение, согласно которому «. всякое дерево, не-зависимо от степени своего развития, может быть назначено в рубку в порядке ухода, и это находится в соответствии с представлением о подвижности положения дерева в ряду остальных деревьев, рост которых может быть (курсив наш) улучшен путем вырубки части окружающих деревьев. Практические результаты этого способа рубок приводят к весьма благоприятным результатам, которые достигнуты в лесах, где он применяется под названием датского способа рубок ухода» [Эйгинген, 1934, с. 77].
Мы не случайно выделили курсивом слова может быть, так как Г.Р. Эйгинген не имел результатов предлагаемых им новых рубок и только предполагал, что они приведут к положительному эффекту7. Он предложил их широкое внедрение, в том числе и потому, что лично побывал в Дании и описал в упомянутой работе результаты рубок ухода в этой стране, которые проводились там с 15-летнего возраста. Далее он приводит описание опытных рубок, заложенных в 1930-е гг, в лесах СССР, причем отметим особо, что возраст начала рубок был не 15 лет. а 27-55 лет, те. намного позднее, чем в Дании. К сожалению, в период Великой Отечественной войны 1941—1945 гг. большая их часть была утрачена. Однако в Ленинградской области подобные опытные рубки, заложенные в те же годы, были сохранены и выше о них мы уже говорили, упоминая работы С.Н. Сеннова.
Законы развития древостоев
В лесоведении собственных признанных (цитируемых) законов пока нет. Во всяком случае, ни в одном из современных учебников о них нет упоминания. Почему-то наши исследователи-лесоводы стесняются называть (и признавать) открытые важные закономерности законами, в противоположность зарубежным исследователям, где законов в экологии десятки, и на них давно ссылаются по именам их авторов [Реймере, 1994]. У нас же можно упомянуть пока только редко упоминаемые «ранговый закон роста деревьев в молодняках» Е.Л. Маслакова [Маслаков, 1981, 1984] и, по видимому7, к рангу' закона можно отнести «основную закономерность морфогенеза древостоев» ГС. Разина [Разин. 1979]. далее названную законом [Рогозин. Разин. 2012].
Согласно этому закону, древостой один раз за свою жизнь достигает предельных состояний развития по коэффициенту' перекрытия кронами горизонтальной поверхности, а также по сомкнутости полога, сумме площадей сечений стволов, текущему приросту и запасам древесины, после чего снижает их тем сильнее, чем выше была начальная густота. Параметры густоты были определены от 0.7-1.0 до 172 тыс. шт./га. В соответствии с этим законом развитие древостоя четко делится на два периода: прогресс и регресс. Момент перехода от прогресса к регрессу' можно считать критическим. Для этих периодов нужен совершенно разный тип хозяйственного воздействия (управления древостоем) - активный на стадии прогресса и пассивный -на стадии регресса. Активное воздействие включа ет в себя удаление живых растений, пассивное -удаление только отмирающих деревьев [Рогозин. Ра зин. 2015].
Однако развитие отдельного дерева продолжается всю его жизнь - дерево неуклонно увеличивает свои размеры, несколько снижая прирост, и регресс для него означает неизбежное и близкое отмирание. В этом и состоит различие между' деревом и древостоем и причина непониманий среди ученых. Древостой как целое, как сообщество растений. обладает свойствами, которых нет у его частей. Поэтому' для понимания законов, по которым развивается древостой, нужно изучать его свойства именно как целого. А для этого «ходом роста» у деревьев-моделей нс обойтись: нужны интегральные характеристики ценоза - фитомасса крон и их объем, сомкнутость полога и. самое важное, отслеживание их динамики. Но как раз весь XX в. этого и не хватало разработчикам таблиц хода роста. По этой причине модели роста разрабатывали по состояниям древостоев в статике на основе однократных наблюдений по классам бонитета, где собственно развитие древостоев не изучалось.
Если в качестве показателя прогресса в развитии древостоя считать подъем текущего прироста по запасу, а регресс связывать с его падением, то в наших моделях еловых древостоев прогресс ограничен 40 годами. В общем-то. падение прироста после 40 лет известно уже давно, но совершенно не было известно, что пик прироста зависит от начальной густоты ценоза; в моделях с высокой густотой прогресс вообще заканчивается очень рано -в 20-25 лет (рис. 1).
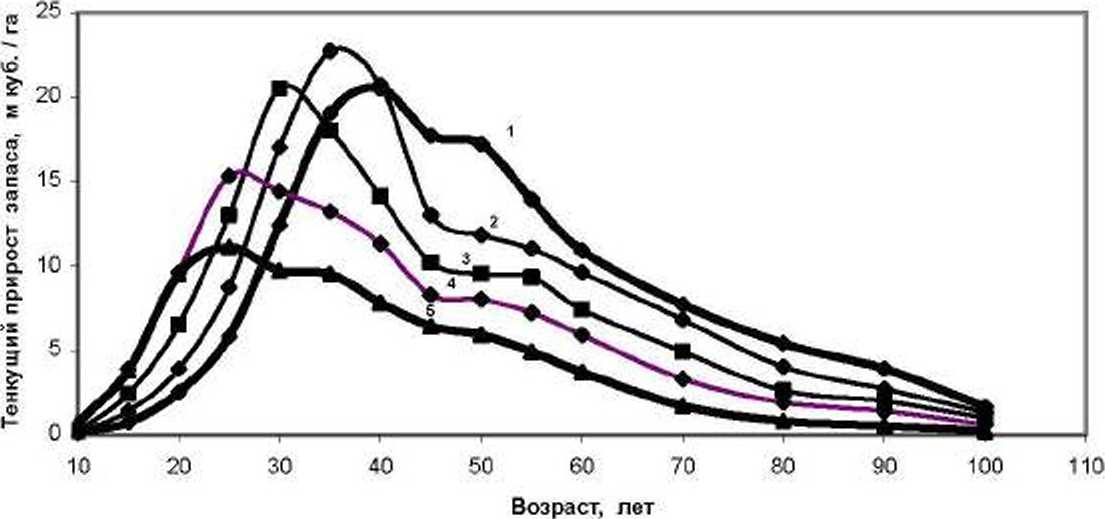
Рис. 1. Текущий прирост по запасу в моделях еловых древостоев в типах условий местопроизрастания С2-СзС начальной густотой:
7 - 1.0; 2 - 1.65; 5 - 2.9; 4 - 5.1; 5 - 14 тыс. шт./га [по Рогозин, Разин, 2012, с. 191]
Если древостой миновал кульминацию прироста. то «переместить» древостой на более производительную линию развития какими-либо разреживаниями почти невозможно, во всяком случае, у ели. Эта инерция появляется из-за уже сформировавшегося цснотического статуса крон, а также вполне определенного генотипического состава ценоза - преобладания правых, либо левых форм деревьев [Голиков. 2014]. Их численность быстро меняться не может, а сами формы обладают совершенно разной адаптацией, о которой многие лесоводы даже не подозревают: левые формы предпочитают прямой свет и слабую конку ренцию. а правые - рассеянный, и толерантны к конкуренции. Кроме того, левые формы лучше растут в сухих условиях, а правые - во влажных. В оптимальных условиях их частота становятся ных условиях их частота становятся одинаковой, но если ценоз оказывается петым, то начинают доминировать правые. а если он более редкий - то обязательно левые формы [Голиков, 2014].
Кроме того, каждая из моделей развития имеет свое распределение частот деревьев по классам Крафта, и оно отражает адаптацию ценоза именно для своей модели и адаптация эта, скорее всего, завершается на пике прироста. Деревья, безусловно, имеют инерцию развития, в том числе и по классам Крафта, т.е. по размерам своих крон. И если ранги по ним уже определились. то быстро изменить их. а тем более «переместить» развитие всего древостоя на более производительную линию во время, а тем более после кульминации прироста вмешательством разреживаниями уже невозможно, так как меняется лишь одно условие из многих - увеличивается только площадь питания дерева. Пик прироста является наиболее критичным периодом, когда деревья стремятся сохранить свою численность, и они адаптируются к нему7 путем формирования малообъемных крон с потерей части фотосинтезирующего аппарата. Теряют его деревья всех рангов, сокращая длину7 и диаметры крон. Полому «фаза чащи» уже не эффективна для изменения прироста [Рогозин. Разин. 2015].
По-ВИДИМОМу, именно Б лот период на большом количество опытных участков С.Н. Ссннова и были проведены рубки ухода, в результате наблюдения за которыми в течение 60 лет было получено кажущееся парадоксальным заключение о невозможности повышения производительности средневозрастных древостоев регулированием их густоты [Сеннов, 1984, 1999, 2005]. Заключение это совершенно не согласуется с современными результатами плантационного выращивания леса, где регулирование густоты начинается в 9-13 лет. и снижение густоты значимо увеличивает их прирост [Плантационное. .. 2007; Большакова, 2007].
Как только прирост древостоя падает, он начинает стареть. Но деревья продолжают увеличивать свои размеры, и это вводит нас в заблуждение. Таксаторы и лесоводы часто оперируют техническими понятиями, и прогресс у них ассоциируется с увеличением размеров деревьев. Поэтому вплоть до достижения нужных кондиций деревья «как бы прогрессируют». После пика прироста фитоценоз уязвим для повреждающих факторов; это учитывает плантационное лесоводство. где после 50 лет целевые древостои вообще рекомендуется вырубать [Плантационное ... 2007].
Из описанных выше опытов выращивания леса и моделирования его развития следует важнейшее практическое положение для лесоводства: рубки ухода должны быть завершены в период до куль-лн/нташ текущего прироста. Для ели это возраст 25 лет в густых (начальная густота 5,1 тыс. шт./га и более) и 30-35 лет — в редких моделях (начальная густота 2.9 тыс. шт./га и менее). В соответствии с этим положением современные прорежива ния и проходные рубки, выполняемые по определенным правилам [Правила... к 2007] не имеют теоретических обоснований, не прошли верификации И ДОЛЖНЫ быть либо прекращены, либо кардинально изменены.
О равномерности структуры фитоценоза
В лесоводстве долгое время безраздельно господствовало мнение о том, что после рубок ухода, причем в любом возрасте, равномерное размещение деревьев при условии оптимальной площади питания должно повышать продуктивность фитоценоза. И равномерно создаваемые лесные культуры много раз подтверждали это. Однако в динамике этот вопрос не отслеживали. В самом деле, молодняк и средневозрастное насаждение - это разные состояния лесного сообщества, и в первом состоянии оно прогрессирует, а во втором находится уже в фазе регресса.
В этой связи оказались примечательны рубки с уходом за «деревьями будущего». Рубки эти предложил немецкий лесовод Вагнер в 1930 гг., и на него ссылается С.Н. Сеннов [2005], когда в лесах Ленинградской области такой уход был проведен в 40-летних сосняках. В этих опытных рубках удалили более 70% (f) деревьев, соседствующих с «деревьями будущего», и оставили на пробной площади равномерно размещенные 112 деревьев (650 шт./га), которые имели диаметры от 12 до 20 см, и это были деревья 1-4 классов Крафта. Спустя 60 лет были подведены итоги, и они оказались просто поразительны. При сравнении деревьев с начальными диаметрами 16 и 12 см, частота которых в сумме была 82%, прирост по диаметру у них за 60 лет достоверно не отличался от контроля, где рубки не проводили. Такой же результат в отношении прироста у ели автор обнаружил и в смешанном древостое, где рост ели не зависел от расстояния до ближайших деревьев березы, и где сила влияния фактора изменения этого расстояния рубками оказалась всего 8% и была недостоверна [Сеннов, 2005; с. 193-194].
На основании этих данных автор делает вывод о том, что развитие дерева слабо зависит от размеров деревьев-соседей и от расстояния до них. Но самым поразительным результатом интенсивного освобождения от деревьев-соседей для «деревьев будущего» оказалась их собственная гибель по точно тем же причинам и с такой же интенсивностью, что и на контроле, где густота оставалась в 3-4 раза больше. Однако этот странный результат автор объясняет лишь предположениями о влиянии корневой конкуренции и некими микроусловиями, оставляя читателя в недоумении, почему7 же так произошло, причем такие же неудачи с «деревьями будущего» были и в Германии [Сеннов.
2005, с. 34-39].
Почему же деревья, получившие после рубок ухода в несколько раз большею площадь питания не ответили на это повышением прироста? Почем}7 они «не послушались» нас? На наш взгляд, причинами «безразличия» деревьев к уход}7 за ними являются некие особые свойства древостоя как целого: во-первых, наличие определенного типа (модели) развития ценоза с фазами прогресса и регресса и воздействие на древостой* скорее всего, было либо на пике его прироста, либо в фазе регресса; во-вторых, групповое размещение деревьев* которое является неотъемлемым свойством древостоя и которое было разрушено рубками при стремлении к более равномерному размещению «деревьев будущего».
О биогруппах С.Н. Сеннов говорит следующее: «.. . групповое размещение деревьев в ельниках в 40-45 лет. отмеченные на старых схемах пробных площадей, сохранились и спустя 60 лет» [Сеннов. 2005, с. 39]. При этом деревья* оставшиеся в одиночестве после разреживания биогрупп* не увеличили свой прирост по сравнению с контрольными деревьями с такими же начальными диаметрами [Сеннов, 1999].
Причины и гипотезы появления биогрупп
Впервые о групповой структуре древостоев упоминают И.В. Логвинов [1955] н Н.Д. Лесков [1956]. В опытной лесной даче Тимирязевской сельхозакадемии в спелых насаждениях группы растений и «пустые» места образовывались независимо от того, возникали ли древостои естественно или создавались культурами [Нестеров, 1961]. Однако в аридной зоне в распределении расстояний между растениями не найдено отклонений от случайного [Жирин. 1970].
Детальный анализ размещения деревьев по территории в синузиях подроста, молодняках, средневозрастных, приспевающих и спелых насаждениях сосны, ели и березы с анализом частот наименьших расстояний между деревьями показал [Ипатов, Тархова, 1975], что на всех участках обнаружено групповое размещение деревьев. Например, в приспевающих и спелых насаждениях встречаемость групп у сосны с расстоянием между деревьями 65-204 см имеет частот}7 28-50%; у ели в распределении наименьших расстояний наблюдаются пики со средним расстоянием между деревьями 50-122 см, и на таких расстояниях встречается 44-48% растений. Исследования в этом направлении показали, что групповая структура свойственна насаждениям сосны обыкновенной, где деревья территориально чаще всего располагаются неравномерно [Грейг-Смит, 1967; Зайченко, 1973; Чудный, 1976; Лебков, 1992; Грибанов,
1993], а также ели тянь-шаньской, где в пространственной структуре насаждений биогруппы выступают как ее управляющие центры [Проскуряков, 1981]. Эти результаты свидетельствуют о том, что в природе, хотя и наблюдается случайное размещение травянистых и древесных растений, но преобладает групповое, контагиозное. В хвойных насаждениях Северо-Запада деревья-лидеры с раннего возраста размещаются нерегулярно, и они сохраняют эту7 неравномерность до возраста спелости [Маслаков, 1999], В последних работах этого региона [Мартынов, 2010] было показано, что групповая структура в насаждениях имеет место, и с возрастом приближается к случайной* Заметим, что в Сибири некоторые авторы [Бузыкин и др*, 1983] также подметили стремление структуры ценоза с возрастом становиться более случайной* Но это совсем не значит что структура полностью становится слу чайной*
Перечисленные работы* рассматривающие биогруппы как природное явление, убедительно показывают, что на развитие деревьев в древостое действуют некие факторы* значимо влияющие на их размещение по территории. Биогруппой считают скопление деревьев (не менее двух) и в раннем возрасте к ним относят деревья-лидеры или, в другой трактовке, деревья будущего. Критерием биогруппы можно принять расстояние между ними до 2.5 м, так как в спелых древостоях 1-2 класса бонитета среднее расстояние между деревьями* рассчитанное из таблиц хода роста, составляет примерно 4 м и расстояние менее 2.5 м можно считать скоплением деревьев* а 6 м и более - окнами в пологе древостоя*
При исследовании искусственных биогрупп В культурах сосны, созданных площадками из 144 и 50 сеянцев в Опытном лесничестве Брянского технологического института, было выяснено, что площадки, которые вначале прижились, в 38-47 лет в 31-34% случаев оказались пустыми или без наличия деревьев-лидеров [Марченко, 1995, с. 109]. Брянские лесоводы давно изучают биогруппы в насаждениях. Так, в структуре 160-летнего сосняка лещинового состава 7СЗЕ обнаружено, что 37-39% деревьев размешались биогруппами [Марченко и др., 1983]. В культурах сосны 13, 29 и 48-летнего возраста в Черниговской области 48% лучших деревьев располагались на расстоянии 2*5 м и менее, из которых 18% росли ближе L5 м [Марченко. Брайко. 1985], В 35-летних культурах сосны с полнотой 1.0 57% лучших деревьев росли в биогруппах, при этом различия в диаметрах одиночных деревьев и деревьев в биогруппах оказались несущественны [Марченко и др., 1983].
Исследования в этом направлении позволили прийти указанным авторам к заключению, что случайное размещение деревьев на самом деле со- всем не случайно и обусловлено энергетическими взаимодействиями в системе Земля-растение. Образование прогалов и скоплений деревьев вызвано наличием дискомфортных и благоприятных зон. Эти закономерности привели брянских исследователей к выводу7, что необходимо изменить всю систему лесоводства, основанную на принципах равномерного размещения деревьев и на том. что древостой представляет собой совокупность растений, организованных борьбой за существование, в свете которых в основном и рассматривалась структура в сообществах древесных растений [Сукачев. 1962l Ткаченко. 1962; Мелехов. 1980; Маслаков. 19 81; Сеннов. 1984]. Предложено отказаться от этих принципов и заложить в основу лесоводства принцип гру ппового размещения деревьев по биоактивным зонам; в БГИТА разработан даже специальный учебный курс «Нетрадиционное лесоводство» [Марченко. Марченко. 1998].
Становится очевидным, что биогруппы для лесного насаждения являются его атрибутом. Однако по канонам лесоводства их необходимо разреживать. Эта рекомендация, похожая на требование. появилась из тезиса, который казался бесспорным. что деревья конкурируют между собой за элементы питания, и чем они ближе друг к другу, тем конкуренция сильнее; поэтому для ее снижения деревья надо «отдалить» друг от друга. И тезис этот убедительно доказали лесные культуры, где деревья растут намного быстрее благодаря равномерному размещению с ранних лет. Однако со временем и в них образуются неравномерности структуры, сохраняющиеся до спелого возраста [Нестеров. 1961].
Значит, в теориях лесоводства оказалось что-то не предусмотренным, и нужно было с пристрастием проверить их. Но верифицировать теории и названный тезис никак не получалось, так как не было ОПЫТНЫХ объектов, кроме единственно упомянутых в работах С.Н. Сеннова. И поэтому правила ухода за лесами как в 1980-е гг., так и ныне действующий их вариант [Правила.... 2007] как бы «не заметили», что постулат о том. что оптимизация густоты б любом возрасте должна приводить к увеличению прироста древостоя, все еще не доказан.
На наш взгляд, для объяснения причин образования неравномерностей в структуре насаждений можно привлечь так называемые геобиологические сети (ГБС), изучаемые геофизикой [Поносов. 2009]. а также гео динамические активные зоны, которые изучает геодинамика и которые являются источником геориска и учитываются при прокладке газо- и нефтепроводов, разработке полезных ископаемых и б строительстве [Копылов, 2014]. Наличие ГБС, тем не менее, подвергают сомнению на основании того, что некоторые сети и зоны в них выделяют только операторы биолокации, т.е. специально обученные люди, и их данные субъективны. Это не значит, что их нельзя использовать в исследованиях. Операторы биолокации используют «прибор», и он имеет название Человек. Если исследователь провел юстировку этого биоприбора по определенным правилам, то он доверяет ему и исследует окружающий мир. Патогенные сети ныне уже абсолютно точно определяются техническими приборами и объективными методами контроля, в частности, фиксацией излучения ГБС на фотоматериалы, и они полностью подтверждают данные биолокации [Агбалян. 2009]. Отмечена и активность исследований в этом направлении: библиография на русском языке за последние 30 лет превышает две согни публикаций.
Мы изучили влияние некоторых геобиологиче-ских сетей, в частности, элементов сетей Хартмана и Карри, известных как «патогенные зоны» [Рогозин. 2011]. В культурах ели в возрасте 21 год измерялась высота деревьев, и они наносились на план на участке размером 100^62 м. Сравнивали растения, выросшие в патогенных и нейтральных местах. Оказалось, что в узлах этих сетей, имеющих диаметры от 0.55 до 2.0 м. растения достоверно снижали высоту' на 14% при 1^= 3 34>Uhs=2 0 На краях узлов снижение было меньше (12%) и еще меньше - на соединяющих узлы полосах (5-8%). Зоны сильного влияния данных сетей занимали 1.9%. а среднего и слабого влияния - 10-11% площади. На сохранность растений они не ВЛИЯЛИ.
В начальной стадии изучения находятся и ГБС с благоприятным влиянием на рост деревьев, о которых в литературе мы не нашли вообще никаких упоминаний. Они имеют узлы (зоны) б диаметре от 1.0 до 8 м. а связи междуг ними представляют собой в плане рисунок в виде цепей, 6- и 4-угольных сеток. Эти сети мигрируют в течение вегетационного периода (май-сентябрь) на расстояние от 0.15 до 0.8 м. За 8 лет полевых маршрутных наблюдений было выяснено, что все обнаруженные нами в Пермском крае наиболее крупные деревья-феномены (так называемые плюсовые деревья), а также наиболее старые деревья в насаждениях располагаются в пределах действия таких зон. [Рогозин, 2014а, б]. В общей сложности мы изучили около 400 таких деревьев в самых разных по составу и происхождению древостоях. Благоприятные зоны по суммарной общей площади составляют в статике примерно 15-18%. а в динамике - до % территории. Было также замечено, что при нахождении дерева одновременно в «хорошей» и «патогенной» зонах у него часто возникали самые разные и часто незначительные, на первый взгляд, пороки ствола, приводящие в старшем воз- расте к появлению корневых гнилей и последующей гибели даже крупного по размерам дерева [Рогозин, 2013а, б].
Также мы начали изучение биогрупп в чистых и смешанных древостоях разного возраста. И уже самые первые наблюдения показали, что буквально все биогруппы с расстоянием между центрами деревьев от 0.4 до 2.5 м, формируются на узлах ГБС, имеющих диаметры зон 1.0, 3.0, 4.5 и 8 м. Поэтому скопление двух-трех и более деревьев в виде биогруппы, а также самые крупные деревья в насаждении безошибочно указывают на места таких благоприятных зон (рис. 2). ______ ____

Рис. 2. Плотная биогруппа из ели и сосны в 130-летнем сосняке-кисличниковом на зоне ГБС с положительным действием диаметром 8 м
Заключение
Все изложенное выше позволяет утверждать, что в структуре древостоя формируется устойчивое неравномерное размещение деревьев по площади в виде хорошо различимых биогрупп, и это является, во-первых, неотъемлемым свойством древостоя (атрибутом) и его необходимо учитывать во всех правилах, технологиях, рекомендациях и моделях по выращиванию леса.
Во-вторых, размещение биогрупп по территории, а также образование прогалов в пологе древостоя оказывается совсем не случайным и вполне может быть обусловлено биофизическими взаимодействиями в системе Земля-растение, связанными с наличием геоактивных зон и геобиологических сетей. Использование биолокации позволило установить, что прогалы приурочены к дискомфортным зонам, имеющим размеры от 0.55 до 2.0 м и занимающим 1.9% территории, а скопления деревьев - к зонам, благоприятным для их роста и имеющим размеры от 1.0 до 8.0 м. Иные объяснения. в частности, действие некоего фактора «микроусловий» среды, например, особенности почвы в конкретной точке, ее микрорельеф, отличия в напочвенном покрове и прочие объяснения не имеют конкретных результатов их изучения. На этот неизвестный фактор обычно ссылаются, когда не могут объяснить что-либо и оставляют его для последующего изучения. Неизвестное же явление никуда не исчезает, и нам нужно двигаться дальше в поисках причин неравномерного размещения деревьев. привлекая для этого новые гипотезы, которые следует проверять, какими бы невероятными они не казались.
Однако наиболее важным в теории лесоводства должно быть признание того, что одновозрастные древостои, как целостные сообщества, в своем развитии имеют два периода: прогресс и регресс, определяемые ходом текущего прироста. Отказ от постушата равномерности и оптимальной площади питания дерева как раз и может совершиться в результате признания этой идеи. В период возрастания прироста рубки ухода могуг быть интенсивными и должны формировать будущую структуру спелого древостоя. После пика прироста наступает регресс и старение древостоя и допустимы только пассивные рубки с удалением сильно отстающих и погибающих деревьев. По этим причинам ныне применяемые в 30 40-летнем возрасте деревьев «прочистки» безнадежно опаздывают по времени и почти бесполезны, а интенсивные «прореживания» и «проходные рубки» не имеют верифицированной теоретической основы, фактически разрушают древостои и поэтому их применение должно быть либо запрещено, либо они должны быть кардинально изменены. Но эти рубки в современном «коммерческом» виде, вероятно, не будут меняться до тех пор, пока существует сама идея получать от них прибыль.
Библиографический сиисок
Агбалян ЮТ. Глобальная энергетическая сеть Хартмана. Мифы и реальность // Сознание и физическая реальность. 2009. № 12. С. 14-20.
Большакова Н.В. Влияние густоты и размещения посадочных мест на рост ели при выращивании культур по интенсивным технологиям: автореф. дис. ... канд. с.-х. наук. СПб., 2007. 24 с.
Бузыкин А.И. и др. Анализ пространственной структуры одновозрастных древостоев И Структурно-функциональные взаимосвязи и продуктивность фитоценозов. Красноярск, 1983. С. 5-12.
Верху нов П.М., Черных В. Л. Таксация леса. Йошкар-Ола: МарГТУ, 2007. 395 с.
Голиков АЛ/ Эколого-диссимметрический подход в генетике и селекции видов хвойных. LAP LAMBERT Academic Publishing, 2014. 162 с.
Гре иг-Смит И. Количественная экология растений. М.: Мир. 1967. 359 с.
Грибанов В. Я. Пространственная структура древостоев U Структура и рост древостоев Сибири. Красноярск: ИЛиД 1993. С. 55-67.
Желдак B.1L Атрохин ВТ Лесоводство: учебник. М/ ВНИИЛМ, 2003. Ч 1. 306 с.
Жирин В.М. Распределение расстояний между деревьями в насаждениях аридной зоны // Науч, тр. ЛТА. Л, 1970. № 131. С. 35-39.
Зайченко Л.П. Исследование размещения деревьев в сосновых древостоях Н Лесная таксация и лесоустройство: межвуз. сб. науч. тр. Красноярск, 1973. Вып. 2. С 82-87.
Ипатов ВС. Тархова Т.Н. Количественный анализ деистических эффектов в размещении деревьев по территории Н Ботан. журн. 1975 . № 9. С. 1237-1250.
Калинин К. К Лесоводство: курс лекций. Изд. 2 стереотипное. Йошкар-Ола: МарГТУ, 2011. 248 с.
Копылов ИС. Научно-методические основы геоэкологических исследований нефтегазоносных регионов и оценки геологической безопасности городов и объектов с применением дистанционных методов: автореф. дис ... д-ра г.-м. наук. Пермь. 2014 48 с.
Лебков В. В. Дендрометрические основы структурно-динамической организации древесных ценозов сосны: автореф. дис. ... д-ра биол. наук. М., 1992. 43 с.
Лесков ИД. Особенности таксационной характеристики и структуры биогрупп в типе леса бор-брусничник Н Тр. по лесн. хоз-ву Уральского лесотехн, ин-та. Свердловск, 1956. С. 56-60.
Логвинов ИВ. О некоторых особенностях в строении и росте смешанных сосново-еловых насаждений типа леса сосняк-черничник Ленинградской области // Техническая информация по результатам научно-исследовательских работ ЛТА. Л., 1955. С. 65-71.
А/аряшшя А.Н. Динамика горизонтальной структуры древостоев ели // Труды С-ПбНИИЛХ. 2010. Вын. 21. С 109-113
Марченко И.С. Биополе лесных экосистем. Брянск: БРИТА, 1995. 188 с.
Марченко И.С., Брайко ВБ. Деревья главного пользования И Молодые ученые 40-лстию победы и 1000-летию г. Брянска. Брянск, 1985. С. 38-39.
Марченко II. С. и др. Рост лучших деревьев в насаждении И Молодые ученые — народному хо зяйству; Тез. докл. 2-й науч.-техн. конф. Брянск: БТИ. 1983. С.46-47.
Марченко И.С, Марченко СП. Нетрадиционное лесоводство: авторский курс / под ред. Е С. Мурахтанова. Брянск: БГИТА, 1998. 419 с.
Маслаков Е.А. Эколого-цснотичсскис факторы возобновления и формирования (организации) насаждений сосны: автореф. дис. ... д-ра биол. наук. Свердловск, 1981. 50 с.
Маслаков Е.Л. Формирование сосновых молодня-ков. М : Лесная Пром-сть, 1984. 168 с.
Маслаков Е.Л. Генезис и динамика социальных структур сосны в фазе индивидуального роста // Таёжные леса на пороге XXI века. СПб.: СПбНИИЛХ, 1999. С. 42-51.
Набатов ИМ. Лесоводство: учебное пособие. М.: МГУ Л. 2002. 190 с.
Нестеров ВТ. Вопросы современного лесоводства. М.: Госсельхозиздат, 1961. 384с.
Поносов В. А. и др. Экологический мониторинг в геофизике: учеб, пособие / под ред. В.А. Поносова. Пермь: ПГНИУ, 2009. 297 с.
Проскуряков М.А. Биогруппы деревьев как управляющие центры в лесу7 // Биоэко логические исследования в еловых лесах Тянь-Шаня. Алма-Ата. 1981. С. 42-54.
Разин ГС. Динамика сомкнутости одноярусных древостоев//Лесоведение. 1979. № 1. С. 23-25.
Рогозин М В. Геобиологические сети Хартмана и Карри в испытательных культурах ели сибирской И Вестник Пермского университета. Серия Биология, 2011. Вып. 2. С. 54-60.
Рогозин М.В. Биогруппы, деревья-лидеры и рубки ухода И Современные проблемы лесного хозяйства и лесоустройства: материалы междунар конф. СПб.: СПбГЛУ. 2012. Т. 1. С 28-29.
Рогозин М.В. Геоактивные зоны и долговечность плюсовых деревьев И Биоразнообразие и куль-туроценозы в экстремальных условиях: материалы докл. II Всерос. науч. конф. Апатиты, 2013а. С 162-167.
Рогозин М.В. Деревья-лидеры, биогруппы и геоактивные зоны // Лабораторный журнал. 20136. № 2 (2). С. 29-44. URL: (дата обращения: 14.04.2015).
Рогозин М.В. Высочайшие деревья Пермского края И Вестник Пермского университета. Серия Биология. 2014. Вып. 3. С. 21-26.
Рогозин М.В. Новые знания для биологии: биополя растений и геоактивные зоны Земли И Образование и наука: современное состояние и перспективы развития: сб. науч. тр. по материалам межлун. научно-практ. конф. Тамбов, 2014. С. 144-147.
Рогозин М.В., Голиков А.М, Разин ГС. О выращивании леса на сухих почвах: теоретические подходы И Вестник Поволжского гос. технолог, ун-та. Серия Лес. Экология. Природопользование 2014. № 3 (23). С. 5-17.
Рогозин M.B.Y Разин ГС. Лесные культуры Тспло-уховых в имении Строгановых на Урале: история, законы развития. селекция ели. 2-е изд Пермь: ПГНИУ. 2012. 210 с. URL: elibrary, m; http ://wvvw. campus, node/ 176612
Рогозин MB., Разин ГС. Развитие древостоев. Модели, законы, гипотезы [Электронный ресурс]. Пермь. 2015. URL: library/node/ 298268.
Сеннов CIL Уход за лесом: экологические основы. М.: Лесная пром-ты 1984. 127 с.
Сеннов СИ* Итоги 60-летних наблюдений за естественной динамикой леса. СПб.: СПбНИИЛХ, 1999. 98 с.
Ояяое С Н. Лесоведение и лесоводство: учебник для студентов вузов. М.: Академия. 2005. 256 с.
Сукачев В.Н* О внутривидовых и межвидовых взаимоотношениях среди растений Н Сообщения института леса. М.: Изд-во АН СССР, 1953. Вып. 1. С. 5-44.
Чудный А. В. О размещении деревьев в популяциях сосны обыкновенной // Лесоведение. 1976. № 5. С 63-68.
Эй тин ген ГР, Рубки ухода за лесом в новом освещении. М_ 1934. 224 с.
Список литературы Рубки ухода: старые и новые теории
- Агбалян Ю.Г. Глобальная энергетическая сеть Хартмана. Мифы и реальность//Сознание и физическая реальность. 2009. № 12. С. 14-20
- Большакова Н.В. Влияние густоты и размещения посадочных мест на рост ели при выращивании культур по интенсивным технологиям: автореф. дис.... канд. с.-х. наук. СПб., 2007. 24 с
- Бузыкин А.И. и др. Анализ пространственной структуры одновозрастных древостоев//Структурно-функциональные взаимосвязи и продуктивность фитоценозов. Красноярск, 1983. С. 512
- Верхунов П.М., Черных В.Л. Таксация леса. Йошкар-Ола: МарГТУ, 2007. 395 с
- Голиков А.М. Эколого-диссимметрический подход в генетике и селекции видов хвойных. LAP LAMBERT Academic Publishing, 2014. 162 с
- Грейг-Смит П. Количественная экология растений. М.: Мир, 1967. 359 с
- Грибанов В.Я. Пространственная структура древо-стоев//Структура и рост древостоев Сибири. Красноярск: ИЛиД, 1993. С. 55-67
- Желдак В.И., Атрохин В.Г. Лесоводство: учебник. М.: ВНИИЛМ, 2003. Ч. 1. 306 с
- Жирин В.М. Распределение расстояний между деревьями в насаждениях аридной зоны//Науч. тр. ЛТА. Л., 1970. № 131. С. 35-39
- Зайченко Л.П. Исследование размещения деревьев в сосновых древостоях//Лесная таксация и лесоустройство: межвуз. сб. науч. тр. Красноярск, 1973. Вып. 2. С. 82-87
- Ипатов В.С., Тархова Т.Н. Количественный анализ ценотических эффектов в размещении деревьев по территории//Ботан. журн. 1975. № 9. С. 1237-1250
- Калинин К.К. Лесоводство: курс лекций. Изд. 2, стереотипное. Йошкар-Ола: МарГТУ, 2011. 248 с
- Копылов И.С. Научно-методические основы геоэкологических исследований нефтегазоносных регионов и оценки геологической безопасности городов и объектов с применением дистанционных методов: автореф. дис.... д-ра г.-м. наук. Пермь, 2014. 48 с
- Лебков В.В. Дендрометрические основы структурно-динамической организации древесных ценозов сосны: автореф. дис.... д-ра биол. наук. М., 1992. 43 с
- Лесков Н.Д. Особенности таксационной характеристики и структуры биогрупп в типе леса бор-брусничник//Тр. по лесн. хоз-ву Уральского лесотехн. ин-та. Свердловск, 1956. С. 56-60
- Логвинов И.В. О некоторых особенностях в строении и росте смешанных сосново-еловых насаждений типа леса сосняк-черничник Ленинградской области//Техническая информация по результатам научно-исследовательских работ ЛТА. Л., 1955. С. 65-71
- Мартынов А.Н. Динамика горизонтальной структуры древостоев ели//Труды С-ПбНИИЛХ. 2010. Вып. 21. С. 109-113
- Марченко И.С. Биополе лесных экосистем. Брянск: БГИТА, 1995. 188 с
- Марченко И.С., Брайко В.Б. Деревья главного пользования//Молодые ученые 40-летию победы и 1000-летию г. Брянска. Брянск, 1985. С. 38-39
- Марченко И.С. и др. Рост лучших деревьев в насаждении//Молодые ученые -народному хо зяйству: Тез. докл. 2-й науч.-техн. конф. Брянск: БТИ, 1983. С.46-47
- Марченко И.С., Марченко С.И. Нетрадиционное лесоводство: авторский курс/под ред. Е.С. Мурахтанова. Брянск: БГИТА, 1998. 419 с
- Маслаков Е.А. Эколого-ценотические факторы возобновления и формирования (организации) насаждений сосны: автореф. дис.... д-ра биол. наук. Свердловск, 1981. 50 с
- Маслаков Е.Л. Формирование сосновых молодня-ков. М.: Лесная пром-сть, 1984. 168 с
- Маслаков Е.Л. Генезис и динамика социальных структур сосны в фазе индивидуального роста//Таёжные леса на пороге XXI века. СПб.: СПбНИИЛХ, 1999. С. 42-51
- Набатов Н.М. Лесоводство: учебное пособие. М.: МГУЛ, 2002. 190 с
- Нестеров В.Г. Вопросы современного лесоводства. М.: Госсельхозиздат, 1961. 384 с
- Поносов В.А. и др. Экологический мониторинг в геофизике: учеб. пособие/под ред. В.А. Поносова. Пермь: ПГНИУ, 2009. 297 с
- Проскуряков М.А. Биогруппы деревьев как управляющие центры в лесу//Биоэкологические исследования в еловых лесах Тянь-Шаня. Алма-Ата, 1981. С. 42-54
- Разин Г.С. Динамика сомкнутости одноярусных древостоев//Лесоведение. 1979. № 1. С. 23-25
- Рогозин М.В. Геобиологические сети Хартмана и Карри в испытательных культурах ели сибирской//Вестник Пермского университета. Серия Биология, 2011. Вып. 2. С. 54-60
- Рогозин М.В. Биогруппы, деревья-лидеры и рубки ухода//Современные проблемы лесного хозяйства и лесоустройства: материалы междунар. конф. СПб.: СПбГЛУ, 2012. Т. 1. С. 28-29
- Рогозин М.В. Геоактивные зоны и долговечность плюсовых деревьев//Биоразнообразие и культуроценозы в экстремальных условиях: материалы докл. II Всерос. науч. конф. Апатиты, 2013а. С. 162-167
- Рогозин М.В. Деревья-лидеры, биогруппы и геоактивные зоны//Лабораторный журнал. 2013б. № 2 (2). С. 29-44. URL: http://labfranep.com/article/7 (дата обращения: 14.04.2015)
- Рогозин М.В. Высочайшие деревья Пермского края//Вестник Пермского университета. Серия Биология. 2014. Вып. 3. С. 21-26
- Рогозин М.В. Новые знания для биологии: биополя растений и геоактивные зоны Земли//Образование и наука: современное состояние и перспективы развития: сб. науч. тр. по материалам междун. научно-практ. конф. Тамбов, 2014. С. 144-147
- Рогозин М.В., Голиков А.М., Разин Г.С. О выращивании леса на сухих почвах: теоретические подходы//Вестник Поволжского гос. технолог. ун-та. Серия Лес. Экология. Природопользование. 2014. № 3 (23). С. 5-17
- Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. 2-е изд. Пермь: ПГНИУ, 2012. 210 с. URL: http.www. elibrary.ru; http://www.campus.psu.ru/library/node/176612
- Рогозин М.В., Разин Г.С. Развитие древостоев. Модели, законы, гипотезы . Пермь, 2015. URL: http://k.psu.ru/library/node/298268
- Сеннов С.Н. Уход за лесом: экологические основы. М.: Лесная пром-ть, 1984. 127 с
- Сеннов С.Н. Итоги 60-летних наблюдений за естественной динамикой леса. СПб.: СПбНИИЛХ, 1999. 98 с
- Сеннов С.Н. Лесоведение и лесоводство: учебник для студентов вузов. М.: Академия, 2005. 256 с
- Сукачев В.Н. О внутривидовых и межвидовых взаимоотношениях среди растений//Сообщения института леса. М.: Изд-во АН СССР, 1953. Вып. 1. С. 5-44
- Чудный А.В. О размещении деревьев в популяциях сосны обыкновенной//Лесоведение, 1976. № 5. С. 63-68
- Эйтинген Г.Р. Рубки ухода за лесом в новом освещении. М., 1934. 224 с
