Ручьевые сообщества макрозообентоса и оценка экологического состояния малых водотоков Самарской Луки

Автор: Чужекова Т.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 1 т.18, 2009 года.
Бесплатный доступ
Охарактеризованы некоторые ручьевые сообщества макрозообентоса малых водотоков Самарской Луки.
Макрозообентос, самарская лука
Короткий адрес: https://sciup.org/148314761
IDR: 148314761
Текст краткого сообщения Ручьевые сообщества макрозообентоса и оценка экологического состояния малых водотоков Самарской Луки
На территории Среднего Поволжья протекает 11.6 тыс. водотоков, из которых 8.8 тыс. (75.9%) являются ручьями - 1.5 тыс. (13.0%) – речками, 1.25 тыс. (10.8%) – малыми реками (Папченков, 2008). И такое соотношение водотоков является достаточно обычным. Однако, несмотря на столь широкое распространение, сообщества ручьев (реокренов) (Чертопруд, 2005) являются практически не изученными, не только в пределах региона, но и по все России работы по этой тематике единичны. Тем не менее, ручьи часто представляют собой верхние звенья рек и могут влиять не только на их гидрохимический состав, но и в совокупности служить источниками биоразнообразия, хотя число видов в каждом может быть не высоко (Дгебуадзе, 2004; Vannote, Minshall, 1980). Поэтому представляется интерес в изучении реокренных сообществах макрозообентоса на территории Поволжья, и в частности на Самарской Луке – уникального места по многим показателям и как места недостаточно исследованного с точки зрения гидробиологии.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для настоящего исследования послужили разовые сборы макрозообентоса, проведенные в июле-августе 2008 г. Было отобрано 58 качественных и количественных проб в 11 водотоках (1-2 станции в зависимости от биотопического разнообразия водного объекта), находящихся на территории Самарской Луки. В городском округе Жигулевск – 4 (ручьи Железнодорожный (RZHD), по ул. Морквашинской (RPM), по ул. Жигулевской (РПЖ) и родник на ул. Репина (RR)) и в Национальном Парке – 7 (подзона постоянной заповедности – ручей Анурьевский (RAN), подзона заповедного восстановления – ручьи Ширяевский (RSHI) и Яблоневый, рекреационная зона – родник Холодный (RH), Большерязанские родники (RBRya) и речка Брусянка (RBRU), зона хозяйственного назначения – ручей Александровский (RAL)). Все ручьи кроме Александровского, вытекающего из малого озера с родниковым питанием, имеют непосредственно родниковое происхождение и
Татьяна Александровна Чужекова научный сотрудник.
характеризуются невысокими температурами даже в летний период 8-16оС. Водообмен водотоков не превышает 0,01м3/с (что соответствует первой размерной группе по Чертопруду (2005) - микрореокрены), однако в реке Бру-сянка этот показатель равен 0,15 м3/с (макрореокрен), что в два раза меньше порогового значения (0,3 м3/с), являющегося границей между ручьями и самыми малыми реками (Чертопруд, 2005; Папченков, 2008). Таким образом, все исследованные водные объекты можно считать ручьями.
Зообентос отбирали количественно - зубчатым водолазным дночерпателем 1/40 м2 (по 2-3 пробы на станции) и качественно – сачком. Образцы промывали через сито с диаметром ячеи 0,6 мм. Пробы фиксировали 4% раствором формалина. Обработку материала проводили с помощью бинокуляра МБС-10, пробы просматривали целиком. Определение проводили под микроскопом. Биомассу организмов зообентоса определяли непосредственно взвешиванием на электронных весах Adventuter OHAUS (0,0001 г). Определение организмов производили с использованием определителей (Определитель пресноводных…, 1977; Панкратова, 1970, 1984; Определитель пресноводных…, 1994-2001).
На станциях измерялись такие абиотические параметры как температура (водным спиртовым термометром с точностью 10С), глубина и ширина водотока (с помощью рулетки), скорость течения (поплавковым методом). Интенсивность водообмена вычисляли по форму Q= υω . Где Q- расход вводы, υ – скорость течения, ω – площадь сечения водотока (нами было использовано допущение, что сечение представляет собой прямоугольник) (Давыдов и др., 1973).
Для оценки экологического состояния водотоков были использованы широко применяемые методики, учитывающие как общее разнообразие организмов макрозообентоса, так и значимость отдельных таксономических групп - индекс сапробности Пантле-Букка в модификации Сладечка, олигохетный индекс Пареле D1, индекс Вудивисса, хирономидный индекс Балушкиной (Балушкина, 1987; Чертопруд, 2002; Шитиков и др., 2003; Основы геоэкологии…, 2004; Sladecek, 1973).
Статистическую обработку проводили в программах Microsoft Excel, BioDiversity Pro и Statistica6.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В составе макрозообентоса было обнаружено 112 таксономических единиц рангом вида и выше. Более половины всего разнообразия (53%) приходилось на долю двукрылых (Diptera) – всего 60 таксономических единиц, из них 38 – это представители семейства Chironomidae (33%). Другие отряды насекомых были представлены более бедно – 1-5 видов. Кроме насекомых значительную роль структуре сообществ играли Oligochaeta (7 таксономических единиц), Hirudinae (6) и Mollusca (10) (рис.1). Такое распределение таксонов совпадает результатами полученными автором ранее при исследовании ручьев г.о. Жигулевска и его окрестностей (Чужекова и др., 2008; Чужекова, Полякова, 2008).
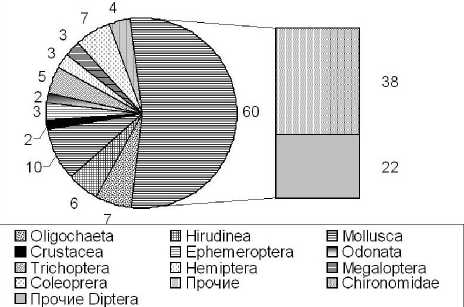
Рис.1. Таксонмический состав макрозообентоса ручьев Самарской Луки. Прим.: Прочие - Nematoda, Hydracarinae, Ple-coptera, Lepidoptera
Число видов и соотношение основных крупных таксонов совпадает с таковым, указанным рядом авторов (Зинченко, 2002; Зинченко, 2002; Чужекова, Полякова, 2008; Зинченко, 2002) для ручьев, где 60% всего разнообразия может приходиться на долю двукрылых, причем 30% составляют личинки сем. Chironomidae. В то время как другие группы беспозвоночных представлены в меньшей степени Число видов обнаруженных в пределах одного водотока (по разовым качественным и количественным сборам) было не велико и варьировало от 7 до 36. Для отдельных станций этот показатель колебался от 7 до 23. Такое число обнаруженных видов в пределах одного реокре-на совпадает с литературными данными (Vannote, Minshall, 1980; Bronmark et all., 1984; Brunke, 2004), указанными для ручьев различных географических зон незначительной схожей глубины и протяженности. Встречаемостью более 10% обладало 34 видов, причем наиболее часто в пробах находились малощетинковые черви Tubifex tubifex и Limnodrillus spp. (Oligochaeta: Tubifecidae) – 67%, далее следовали двукрылые Prodiamesa olivacea (Diptera: Chironomidae) – 48% и Dicranota bimaculata (Diptera: Limonidae) –38%, Pseudodiamesa gr. ni-vosa (Diptera: Chironomidae) а также пиявки Helobdella stagnalis (Hirudinae, Glossiphonidae) по 33%. Также достаточно регулярно встречались личинки комаров долгоножек Tipula (Yamatotipula) sp. (29%) и пиявки Erpobdella lineata (24%). Эти организмы преимущественно и определяли величины обилия сообществ. Видовой состав можно охарактеризовать как обычный для малых рек Поволжья. Однако есть некоторые расхождения – так значительную роль в сообществах приобретают личинки Chironomidae рода Pseudodia-mesa (Pseudodiamesa gr. nivosa и Pseudodiamesa brankii) видами, являющиеся холодноводными и тяготеющими к олигосапробной зоне, но способными жить при незначительных глубинах и растительных обрастаниях смачиваемых водой. Эти виды при уменьшении глубины и при уменьшении антропогенной нагрузке (Паньков и др., 2008; Lencioni et all., 2006; Lencioni et all., 2006) могут заменять в сообществах Prodiamesa olivacea (сходный по экологическим характеристикам вид, но обладающий большей экологической валентностью по отношению к содержанию РОВ). Тем не менее, большую часть разнообразия Chironomidae формировали представители семейства, приуроченные к мягким илистым грунтам и слабым течениям – Chironominae – 55,3%, но они не делали столь существенного вклада в величины обилия (за исключением личинок рода Chironomus). Другие подсемейства были представлены слабее как по числу видов(Tanypodinae – 21,1%, Ortocladinae – 13,2%, Prodiamesinae и Diamesinae по 5,3%) так и по величинам обилия, хотя литературные данные указывают на обратную ситуацию (Зинченко, 2002; По-здеев, 2006; Lencioni et all., 2006) и значительную роль в структуре сообществ именно личинок Ortocladinae. Однако такое не соответствие, скорее всего, может быть связано с индивидуальными различиями ручьев по морфологическим характеристикам и различной антропогенной нагрузкой (Галимзянова и др., 2008).
Численность и биомасса сообществ варьировали в широких пределах: от 1,4±0,2 тыс. экз./м2 и 3,2±0,8 г/м2 (RPM2) до 19,2±2,7 тыс. экз./м2 и 122,4±4,5 г/м2 (RBRU1) (Прил. табл.1). Но для большинства станций величины обилия сообществ укладывались в диапазон 3-20 тыс.экз./м2 и от 8-70 г/м2. Полученные величины обилия могут быть охарактеризованы как высокие и обычные для ручьев, как можно судить по не многочисленным данным по водотокам именно такого типа (Галимзянова и др., 2008; Паньков и др., 2008; Чужекова, Полякова, 2008). Высокие показателей обилия некоторые авторы склонны связывать с отсутствием бентоядных рыб обитания и недостаточной представленностью беспозвоночных хищников (Паньков и др., 2008). Так как из преимущественного поступления органического вещества в форме растительного опада и не высоких скоростей течения в структуре макрозообентоса ручьев в значительной мере развиваются группы грунтоедов и детритофагов-собирателей (Ивановский, 2008а,б; Паньков и др., 2008; Паньков и др., 2008), биомасса которых определяется содержанием органического вещества в донных отложениях в независимости от систематической принадлежности (Чу-жекова, Полякова, 2008). В целом сообщества могут быть охарактеризованы как олигомиксные, поскольку основу структуры составляют 1-2 вида, обладающие сходными экологическими характеристиками и определяющие 7090% величин обилия. Наибольший вклад в численность на различных станциях вносили мелкие личинки Chironomidae ( Tanitarsus spp., Glyptotendipes gripencoveni, Procladius (P.)sp. ). И на численность и на биомассу оказывали влияние малощитинковые черви семейства Tubificidae, крупные личинки Chi-ronomidae ( Pseudodiamesa spp., Prodiamesa olivacea, Chironomus spp .). Только на биомассу оказывали влияние комары долгоножки из семейств Tipulidae ( Tipula spp .) и Limoniidae (Dicranota bimaculata) (Прил. табл.1). На станции RYAB – сообщество было сформировано из оксифильных псамофильных организмов отряда двукрылых- Dicranota bimaculata, Tipula spp. и Telmatoscopus britteni ( Psychodidae ), при отсутствии Chironomidae и Tubifeci-dae. Виды формирующие основу сообществ - это виды обладающие большим ареалом, но к группе эврибионтов могут быть отнесены только Tubifecidae и личинки рода Chironomus. Прочие из перечисленных организмов предпочитают хорошо аэрируемые местообитания (Определитель пресноводных…, 1994-2001). Следует отметить, что личинки других отрядов насекомых, кроме двукрылых, имели единичную встречаемость и не оказывали влияния на показатели обилия, тогда как в малых реках Самарской области их роль значительно более существенна (Филиппов, Барбашова, 2006; Голубая книга…, 2007; Чужекова, Филиппов, 2008).
На основе доминирующих групп на уровне семейства было выделено 2 крупных типа сообществ:
-
1) «Хирономидные» - величины обилия определяют личинки Chironomi-dae, а в биомассу могут также вносить существенный вклад крупные личинки
комаров долгоножек (Tipulidae и Limoniidae) или пиявки ( Erpobdella spp ., He-lobdella stanalis ).
-
2) «Тубифицидные» - численность и биомассу сообществ определяли малощитинковые черви сем. Tubificidae ( Tubifex tubifex и Limnodrillus spp. в различных соотношениях), а как со-доминирующие формы выступали личинки Chironomus spp . (доля которых по биомассе могла достигать 70%) или другие формы из подсемейств Chironominae и Tanypodinae (рис.2).Сообщества «хирономидного» типа могут быть разделены на 2 группы. Группа - 1 сообщества сформированные преимущественно двукрылыми ( Pseudiamesa spp. + D. maculata и/или T. (Yamatotipula) spp.), однако рядом авторов эти виды указываются как субдомининты при наличие таких доминирующих форм, как ручейники Potamophylax nigricornis и поденки Baetis rhodani для реокренов с подобными морфометрическими характеристиками (Ивановский, 2008а,б; Паньков и др., 2008; Галимзянова и др., 2008). Эти сообщества были сформированы преимущественно в ручьях на территории НП Самарская Лука, на территории же г. Жигулевска при сходных параметрах течения и глубины в сообществах макрозообентоса заменялись на сообщества второй группы – Prodiamesa olivacea + Erpobdella spp . и Helobdella stanalis. Доминирование личинок Chironomidae было показано на станциях с глубиной около 7,1 см и скоростях течения приблизительно равным 0,2 м/с. Сообщества «тубифицид-ного» типа формировались как в ручьях на территории как города, так и национального парка. Эти сообщества были приурочены к более низким скоростям течения и большим глубинам. Было показано наличие отрицательной корреляции между долей малощитинковых червей (%) и скоростью течения (r= -0,66, p<0,05). Расхождения сообществ по доминирующим формам было достоверно связано с условиями биотопов. Так, с помощью t-критерия Стьюдента было подтверждено, что сообщества с преобладанием Chironomidae формировались на более быстром течении – 0,16≤V≤0,23 м/с (t=3,989149, df=17, p=0,000949) и мелких глубинах - 3,2≤H≤10,9 см - (t=-3,23, df=17, p=0,004741), чем сообщества с доминирование Tubifecidae 0,04≤V≤0,12м/с и 14,3≤H≤35,1см соответственно.
Рис. 2. Дендрограмма сходства станций на по долям величин обилия доминирующих и субдоминирующих групп
(данные трансформированы как координаты на плоскости в correspondence analysis в программе BioDiversity Pro. Объединение проведено по методу Уорда, по оси ординат – евклидово расстояние, по оси абсцисс - станции) с учетом абиотических факторов (V-скорости и Н-глубины)
Несмотря на то, что течение во всех водотоках может быть охарактеризовано как незначительное (т.к. не превышает 0,3 м/с), что должно было привести к формированию сходных лимнофильных сообществ, но достоверные отличия по данному фактору, а также по глубине послужили причиной формирования различной структуры сообществ. Поскольку оценка экологического состояния проводилась на основе данных о составе и величинах обилия донных беспозвоночных, то, как следствие, классы качества вод, оцененные по совокупности индексов, также зависели от скорости течения (r=0.76, p<0,05) (табл.1). Тем самым была подтверждена общая закономерность, известная на основе разницы сапробных значений в для реофильных и лимно- фильных сообществ формируемы соответственно на перекатах и в заводях (Основы геоэкологии…, 2004).
|
Индекс/ст анция |
Hn |
Hb |
S |
D1 |
Wd |
Kch |
ККВ |
V, м/с |
Н, см |
|
RAL1 |
1,4 |
1,6 |
1,3 |
1,4 |
6 |
6,1 |
II-III |
0,183 |
5 |
|
RPM1 |
1,8 |
2,3 |
3 |
2,8 |
3 |
0,1 |
0,286 |
14 |
|
|
RAN1 |
2 |
1,8 |
3,8 |
2,7 |
4 |
1,7 |
0,214 |
3,5 |
|
|
RAN2 |
2,3 |
2,3 |
2,6 |
1,2 |
9 |
4,3 |
0,214 |
3,5 |
|
|
RSHI1 |
2,9 |
2,3 |
2,5 |
0 |
6 |
0,4 |
0,25 |
3,5 |
|
|
RYAB |
1,8 |
1,7 |
3,3 |
3 |
6 |
- |
II-III |
0,25 |
6,5 |
|
RPZh1 |
2,3 |
1,2 |
3,7 |
39,9 |
2 |
2 |
0,143 |
5,5 |
|
|
RZHD |
2,4 |
2,4 |
3 |
36,8 |
3 |
1,6 |
0,15 |
22 |
|
|
RPZh2 |
2,1 |
2 |
3,4 |
12 |
2 |
2,4 |
0,073 |
5 |
|
|
RSHI2 |
2,1 |
2 |
3,4 |
12 |
5 |
2,4 |
III-IV |
0,2 |
2 |
|
RPM2 |
2 ,4 |
2 ,7 |
3, 3 |
2 6,5 |
3 |
1 ,5 |
0 ,11 1 |
42 |
|
|
RBRU1 |
2,2 |
2,1 |
3,8 |
43,5 |
2 |
5,5 |
0,15 |
45 |
|
|
RBRya1 |
0,4 |
0,7 |
3,4 |
2,8 |
3 |
5,7 |
0,12 |
3 |
|
|
RAL2 |
2,8 |
2,9 |
3,4 |
28,2 |
6 |
6,2 |
III-IV |
0,143 |
15 |
|
RR |
1,7 |
1,9 |
3,8 |
42,4 |
2 |
6,3 |
0,001 |
17 |
|
|
RH |
1,6 |
1,5 |
3,7 |
66,4 |
4 |
3,6 |
0,001 |
35 |
|
|
RBRU2 |
1,6 |
2,7 |
3,6 |
86,9 |
6 |
6,4 |
0,11 |
27 |
|
|
RBRya2 |
1,3 |
0,8 |
3,7 |
94,1 |
6 |
1,2 |
0,1 |
3 |
|
|
RAL0 |
2,1 |
1,2 |
3,4 |
77 |
5 |
3,1 |
IV-V |
0,001 |
35 |
Табл. 1. Оценка качества вод по структуре макрозообентоса с использованием различных индексов.
Hn – индекс Шеннона по численно с ти, Hb – индекс Ше н нона по би о массе, S-индекс сапроб н ости , D1- олигохетный индекс, Wd –индекс Вудив и сса, Kch – х и ономидный индекс Балушкиной, ККВ – класс качества вод, V – скорость течения м/с, Н – глубина см.
Как видно из таблицы (табл.1) расхождения по качеству вод между ручьями принадлежащими территории национально парка и города нет, т.к. станции одного ручья различающиеся по своим морфометрическим характеристикам и находящиеся на незначительном удалении друг от друга (не более ста метров) могут принадлежать к разным сапробным зонам, испытывая одинаковую антропогенную нагрузку (RPM, RAL). Модальным классом качества вод является переходная зона между III-IV, что соответствует умеренно загрязненным и загрязненным водам, в то время как большинство малых рек Самарской области относят к II-III классам (Зинченко, 1994; Филиппов, Бар-башова, 2006 Голубая книга…, 2007; Чужекова, Филиппов, 2008). Такая ситуация может быть связана с размерами водотока, т.к. из-за не большой протяженности реокрена относительная нагрузка на его водосбор будет больше, чем на водосбор малой реки. Увеличение размеров предоставляет большее число возможных для обитания биотопов, что соответственно влияет на общее β-биоразнообразие, что как следствие ведет за собой более высокие показатели по качеству вод (Vannote, Minshall, 1980).
Приложения .Табл.
Величины обилия значимых групп беспозвоночных в сообществах макрозобетоса
|
Oligochaeta |
Hirudinae |
Tipulidae |
Limoniidae |
Chironomidae (Diptera) |
Все Chironomidae |
Весь макрозообенто |
||||||||||||||||||
|
(Diptera) |
(Diptera) |
Prodiamesinae |
Deamesinae |
Othocladinae |
Chironominae |
Tanypodinae |
Pupa |
|||||||||||||||||
|
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
N |
B |
|
|
RAL0 |
1,14± 0,67 |
2,2± 1,6 |
0,02± 0,02 |
0,0± 0,0 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
0,06± 0,02 |
0,1± 0,1 |
0,14± 0,14 |
0,2± 0,2 |
0,02± 0,02 |
0,0± 0,0 |
0,22± 0,14 |
0,3± 0,2 |
1,48± 1,16 |
9,2± 8,5 |
|
RAL1 |
0,12± 0,04 |
0,4± 0,3 |
0,04± 0,00 |
0,3± 0,1 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
5,39± 3,57 |
1,6± 1,1 |
0,51± 0,23 |
0,7± 0,4 |
0,07± 0,04 |
0,0± 0,0 |
5,96± 3,58 |
2,4± 1,1 |
8,68± 4,32 |
53,5± 35 |
|
RAL2 |
2,35± 0,53 |
1,9± 0,5 |
0,47± 0,09 |
3,7± 0,6 |
0,40± 0,21 |
44,5± 26,9 |
0,03± 0,03 |
0,0± 0,0 |
- |
- |
- |
- |
0,11± 0,11 |
0,1± 0,1 |
4,95± 1,58 |
1,9± 0,6 |
0,31± 0,16 |
0,4± 0,2 |
0,08± 0,0 |
0,0± 0,0 |
5,44± 1593 |
2,4± 0,6 |
8,32± 1,1 |
8,3± 1,1 |
|
RBrya1 |
0,09± 0,05 |
0,1± 0,0 |
0,01± 0,01 |
0,0± 0,0 |
- |
- |
- |
- |
- |
- |
- |
- |
0,04± 0,02 |
0,0± 0,0 |
3,12± 1,52 |
2,9± 1,5 |
- |
- |
0,04± 0,02 |
0,1± 0,1 |
3,20± 1,52 |
3,1± 1,5 |
3,32± 1,6 |
3,2± 1,6 |
|
RBrya 2 |
12,99± 7,48 |
33,2± 22,2 |
0,01± 0,01 |
0,0± 0,0 |
- |
- |
- |
- |
0,12± 0,08 |
0,0± 0,0 |
- |
- |
0,08± 0,04 |
0,1± 0,0 |
0,17± 0,06 |
0,0± 0,0 |
- |
- |
- |
- |
0,37± 0,11 |
0,1± 0,0 |
13,80± 6,83 |
37,1± 21,5 |
|
RYAB |
0,04± 0,00 |
0,0± 0,0 |
0,04± 0,04 |
0,4± 0,4 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
1,34± 0,06 |
15,1± 3,3 |
|
RAN1 |
0,04± 0,03 |
0,5± 0,5 |
- |
- |
- |
- |
- |
- |
0,08± 0,02 |
0,1± 0,0 |
1,15± 0,13 |
10,1± 1,3 |
- |
- |
0,05± 0,04 |
0,0± 0,0 |
- |
- |
- |
- |
1,28± 0,13 |
10,2± 1,3 |
1,47± 0,11 |
12,4± 1,4 |
|
RAN2 |
0,18± 0,02 |
1,2± 1,0 |
- |
- |
- |
- |
- |
- |
3,60± 0 |
7,2± 1,8 |
4,64± 1,89 |
50,8± 22,7 |
- |
- |
6,10± 1,00 |
5,3± 1,8 |
0,08± 0,06 |
0,1± 0,0 |
- |
- |
14,42± 2,14 |
63,4± 22,8 |
15,18± 3,38 |
72,9± 27 |
|
RSHI1 |
- |
- |
0,01± 0,01 |
0,0± 0,0 |
- |
- |
0,03± 0,02 |
0,4± 0,3 |
0,13± 0,13 |
0,1± 0,1 |
1,52± 0,65 |
25,8± 11 |
0,68± 0,25 |
0,8± 0,3 |
0,03± 0,03 |
0,0± 0,0 |
- |
- |
0,01± 0,01 |
0,0± 0,0 |
2,37± 0,71 |
26,7± 11 |
5,19± 0,87 |
90,8± 25,4 |
|
RSHI2 |
0,39± 0,24 |
1,4± 0,9 |
0,17± 0,14 |
6,3± 6,0 |
- |
- |
2,01± 1,09 |
11,6± 6,4 |
- |
- |
- |
- |
0,09± 0,05 |
0,2± 0,1 |
0,45± 0,16 |
2,1± 0,9 |
0,07± 0,07 |
0,3± 0,3 |
2,63± 1,11 |
14,2± 6,5 |
3,21± 1,64 |
22,0± 12,6 |
||
|
RH |
1,86± 1,14 |
9,6± 5,2 |
- |
- |
0,06± 0,05 |
6,1± 5,6 |
0,64± 0,22 |
7,4± 2,7 |
- |
- |
- |
- |
0,04± 0,04 |
0,1± 0,1 |
0,64± 0,28 |
0,9± 0,4 |
0,16± 0,12 |
0,3± 0,2 |
- |
- |
0,84± 0,31 |
1,2± 0,4 |
2,80± 1,32 |
13,9± 2,9 |
|
RBRU1 |
8,34± 4,46 |
23,4± 12,0 |
- |
- |
- |
- |
0,13± 0,07 |
0,8± 0,5 |
0,16± 0,02 |
0,6± 0,2 |
- |
- |
0,09± 0,09 |
0,1± 0,1 |
9,31± 1,49 |
93,5± 9,2 |
1,08± 0,35 |
3,8± 1,1 |
0,05± 0,01 |
0,2± 0,1 |
10,69± 1,53 |
98,2± 9,2 |
19,18± 2,68 |
122,4± 4,5 |
|
RBRU2 |
9,52± 0,15 |
25,6± 0,3 |
0,32± 0,85 |
14,9± 2,4 |
- |
- |
0,16± 0,00 |
0,8± 0,5 |
- |
- |
- |
- |
0,10± 0,02 |
0,1± 0,0 |
0,82± 0,18 |
10,2± 2,4 |
0,02± 0,02 |
0,1± 0,1 |
- |
- |
0,94± 0,19 |
10,5± 2,4 |
10,96± 0,04 |
56,9± 6,1 |
|
RPZH1 |
1,45± 0,29 |
54,9± 23 |
- |
- |
0,44± 0,02 |
40,6± 7,8 |
0,25± 0,05 |
2,0± 0,5 |
1,63± 0,36 |
8,7± 2 |
0,31± 0,09 |
5,5± 1,1 |
- |
- |
0,05± 0,04 |
0,0± 0,0 |
0,16± 0,07 |
0,3± 0,2 |
- |
- |
2,15± 0,38 |
14,5± 2,3 |
3,64± 0,17 |
70,9± 21,5 |
|
RPZH2 |
0,39± 0,24 |
1,4± 0,9 |
0,17± 0,14 |
6,3± 6,0 |
- |
- |
- |
- |
2,01± 1,09 |
11,6± 6,4 |
- |
- |
- |
- |
0,09± 0,05 |
0,2± 0,1 |
0,45± 0,16 |
2,1± 0,9 |
0,07± 0,07 |
0,3± 0,3 |
2,63± 1,11 |
14,2± 6,5 |
3,21± 1,64 |
22,0± 12,6 |
|
RR |
2,77± 1,64 |
9,4± 5,2 |
- |
- |
- |
- |
- |
- |
1,20± 0,49 |
4,0± 1,7 |
- |
- |
- |
- |
2,44± 1,25 |
7,4± 3,6 |
0,08± 0,06 |
0,4± 0,3 |
0,03± 0,03 |
0,2± 0,2 |
3,75± 1,35 |
12,0± 4 |
6,53± 0,89 |
22,1± 2,6 |
|
RZHD |
1,31± 0,58 |
3,6± 1,9 |
1,63± 0,5 |
15,6± 5,5 |
- |
- |
- |
- |
0,37± 0,35 |
2,0± 1,9 |
- |
- |
- |
- |
- |
- |
0,08± 0,06 |
0,4± 0,3 |
0,01± 0,01 |
0,1± 0,1 |
0,47± 0,36 |
2,5± 1,9 |
3,55± 0,84 |
24,4± 9,6 |
|
RPM1 |
0,08± 0,05 |
2,6± 2,5 |
- |
- |
- |
- |
- |
- |
0,21± 0,1 |
0,4± 0,3 |
0,04± 0,02 |
0,8± 0,4 |
2,07± 1,25 |
2,9± 1,8 |
- |
- |
0,13± 0,06 |
0,1± 0,1 |
0,01± 0,01 |
0,0± 0 |
2,47± 1,26 |
4,2± 1,9 |
2,91± 1,58 |
35,8± 21,5 |
|
RPM2 |
0,36± 0,19 |
0,9± 0,4 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
0,05± 0,05 |
0,0± 0,0 |
0,15± 0,09 |
0,4± 0,4 |
0,71± 0,17 |
1,0± 0,3 |
0,03± 0,01 |
0,1± 0,1 |
0,93± 0,2 |
1,6± 0,5 |
1,36± 0,22 |
3,2± 0,8 |
Прим.: N – численность тыс. экз./м2, В – биомасса г/м2
Таким образом, скорость течения и глубина даже при их незначительных величинах могут совместно выступать как фактор, определяющий структуру сообществ и приводящий к ее дифференциации при достаточно сходных условиях среды, тем самым, влияя на средний класс качества вод, что соответствует закономерностям известным для более крупных водотоков.
Список литературы Ручьевые сообщества макрозообентоса и оценка экологического состояния малых водотоков Самарской Луки
- Балушкина Е.В. Функциональное значение личинок хирономид в континентальных водоемах. Л.: Наука, 1987. 180с.
- Галимзянова А.В., Тахтеев В.В, Окунева Г.Л. Таксономическая структура и сезонная динамика сообщества зообентоса Олхинского незамерзающего источника (Южное Прибайкалье)// Материалы III всероссийской научной конференции «Принципы и способы сохранения биоразнообразия». 2008. С. 127-129.
- Голубая книга Самарской области: редкие и охраняемые гидробиоценозы/ под ред. Г.С. Розенберга, С.В. Саксонова. Самара, 2007. 200с
- Давыдов Л.К., Дмитриева А.А., Конкина Н.Г. Общая гидрология. Л.: Гидрометиздат, 1973. 462 с.
- Дгебуадзе Ю.Ю. Малые реки как объект экологических исследований: некоторые итоги и перспективы // Тезисы докладов Всероссийской конференции: Экосистемы малых рек: биоразнообразие, биология, охрана. 2004.С. 21-22
