Рыжая полевка (Clethrionomys glareolus Shreb.) в коренных и антропогенных ландшафтах Восточной Фенноскандии
 в коренных и антропогенных ландшафтах Восточной Фенноскандии")
Автор: Ивантер Эрнест Викторович, Жулинская Ольга Юрьевна, Курхинен Юрий Павлович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 8 (169), 2017 года.
Бесплатный доступ
На основании многолетних стационарных и экспедиционных исследований в Восточной Фенноскандии рассматриваются популяционно-экологические особенности рыжей полевки в условиях антропогенной трансформации, динамика численности таежных экосистем, закономерности пространственной структуры популяции, изменений численности, взаимоотношения с видами-конкурентами. Численность рыжих полевок находится под контролем большого числа факторов (демографический состав популяции в настоящий и предыдущий моменты времени, погодные и кормовые условия, состояние популяции совместно обитающих видов, качество местообитания и т. д.). Установлено, что концентрированные рубки лесов приводят к усилению нестабильности и крайнему упрощению популяционной организации и экологической структуры видового населения рыжей полевки.
Биотоп, вырубки, экосистема, популяция, грызуны
Короткий адрес: https://sciup.org/14751253
IDR: 14751253 | УДК: 599.33:591.5
The bank vole (Clethrionomys glareolus Shreb.) in indigenous and anthropogenic landscapes of Eastern Fennoscandia
Rodents are widespread in terrestrial ecosystems. These animals are bio indicators of the environmental conditions; therefore, it is convenient to use them for solving a number of applied and theoretical problems of animal ecology. The main aim of the research to study comprehensively the ecology of the bank vole inhabiting Karelia. In the study, the following methods of counting the number of small mammals were used: trap-lines and trapping grooves. The abundance of voles is controlled by a large number of factors (weather and feed conditions, habitat quality, the status of coexisting species etc.). It was revealed that concentrated forest felling leads to the increased instability and extreme simplification of the population organization, to the streamlining of the ecological structure of the population of the Bank vole species.
Текст научной статьи Рыжая полевка (Clethrionomys glareolus Shreb.) в коренных и антропогенных ландшафтах Восточной Фенноскандии
К числу наиболее важных факторов, воздействующих на таежные экосистемы Северной Евразии, относятся интенсивные рубки леса. Их активное влияние на природные комплексы началось уже довольно давно: в Скандинавских странах и Финляндии – во второй половине XIX века, в Карелии – в 1930-е годы. Промышленное освоение лесных ресурсов с применением тяжелой техники достигло пика в середине ХХ века. Все это не могло пройти мимо профессионального внимания экологов, однако оно было сосредоточено преимущественно на явлении трансформации самих лесных древостоев, тогда как судьба обитающих здесь животных, и в частности мелких лесных грызунов (а они всегда были и остаются весьма существенными компонентами лесного биоценоза), подчас не входила в их поле зрения.
Исследования в этом направлении проводились, были и публикации [5], [6], [13], [15], но все же вопросов оставалось не меньше, чем ответов. Это и определило главную цель настоящей
работы – восполнить образовавшийся пробел, по крайней мере, в отношении изучения мелких лесных грызунов, и прежде всего повсеместно доминирующей европейской рыжей полевки.
МАТЕРИАЛЫ И МЕТОДЫ
Учеты мелких грызунов проводились по унифицированной методике [1], [2], [4], [7], сочетавшей работу на полевых стационарах (Ладожский, Гомсельгский, Картешский, Каскеснаволок, Агробаза, заповедники «Кивач» и «Костомукшский») с широкими экспедиционными обследованиями, охватившими значительную часть так называемой Восточной Фенноскандии. К этой территории мы относим восточную часть Балтийского кристаллического щита: Кольский п-ов, Финляндию, Карелию и Карельский перешеек Ленинградской области.
Кроме того, в одном из наиболее исследованных в экологическом отношении Кондопожском районе Карелии (на территории Гомсельгского стационара) в пределах ландшафта одного типа (сельговый среднезаболоченный с преобладанием сосняков) было выделено 4 категории коренных сосновых лесов в соответствии со степенью их фрагментации: крупные массивы (25–30 га), относительно небольшие участки (6–7 га) и совсем мелкие (по 1–2 га) островки соснового леса. Для сравнения использовались данные учетов зверьков, полученные в крупных массивах нефраг-ментированных сосняков заповедника «Кивач» (общей площадью около 3 тыс. га). На всех этих участках работы велись одновременно, в равных объемах и по единой методике, а сравниваемые территории разделяли всего 20 км. При этом все фрагменты сосняков обладали здесь близкими характеристиками рельефа, почв, исходного типа растительности и были окружены сходными по структуре экосистемами (производные лиственные и смешанные насаждения). Это позволило осуществить достаточно корректный сравнительный анализ данных за большой временной ряд (более 30 лет) синхронных наблюдений.
Общий объем проведенных учетов превысил за период работы 470000 ловушко-суток и 8500 канавко-суток, а общее число добытых и исследованных грызунов составило 8900 экз., из которых 6270 приходится на исследованный вид.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Рыжая полевка – один из наиболее распространенных и многочисленных в регионе видов мелких млекопитающих, а среди лесных грызунов и вовсе абсолютный доминант. Средний (1966–2016 годы) многолетний показатель его учета (табл. 1) составил за весь период наблюдений 1,55 экз. на 100 ловушко-суток (с долей уловов среди грызунов – 79,2 % ). В отловах же канавками показатель численности рыжей полевки равнялся 1,1 экз. на 10 канавко-суток, что составляет 44,0 % от общего числа пойманных грызунов.
Что же касается влияния концентрированных рубок хвойных лесов на население рыжих полевок, то в целом оно отрицательное. На свежих лесосеках возрастом до 1 года численность этой полевки сокращается в 5 раз. Правда, в первые 3–4 года после рубки показатель учета заметно возрастает, однако продолжает (причем в 1,5 раза) уступать таковому в старовозрастных хвойных лесах. Тем не менее через 5–6 лет после рубки численность вида восстанавливается (табл. 2). Очень сильные изменения происходят с индексом доминирования рыжей полевки, особенно при расчетах этого показателя в долях улова мышевидных грызунов. Наблюдения за динамикой численности видов в биотопах сукцессионного ряда с различной давностью рубки (от 1 до 25 лет) доказали позитивную связь между этим показателем и индексом доминирования вида (коэффициент корреляции r = + 0,95, p < 0,05). При этом высокая численность рыжей полевки в не-дорубах и опушечных зонах сохраняется. Здесь создаются своеобразные резерваты и центры дальнейшего расселения и восстановления видового населения, что в результате существенно сглаживает отрицательное воздействие концентрированных рубок леса на состояние популяций рыжих полевок.
В антропогенном ландшафте «пики» и «депрессии» численности рыжей полевки не столь четко выражены, как на заповедной территории. Здесь обычно наблюдается более резкий подъем ее численности в начале лета, но к осени скорость роста популяции заметно снижается. Полученные данные показывают, что этот вид сравнительно легко переносит такие последствия рубок, как фрагментации хвойных лесов. Более того, в массивах сосняков экспериментальной территории (слабая степень фрагментации) численность рыжей полевки в среднем оказалась несколько выше, а коэффициент вариации – ниже, чем в нефрагментированных лесах (рис. 1). Сильная степень фрагментации (до мелких массивов) сокращает численность зверьков и нарушает ее стабильность (рост CV ). Тем не менее рыжая полевка абсолютно доминирует даже на небольших участках сосняков (семенных куртинах), окруженных свежими и зарастающими
Таблица 1
Сводные данные о численности и соотношении видов лесных мышевидных грызунов в Приладожье (1966–2016 годы)
|
Вид |
Учеты ловушко-линиями |
Учеты ловчими канавками |
||||
|
Общее число отловленных зверьков, экз. |
Экз. на 100 ло-вушко-суток |
Доля вида в уловах, % |
Общее число отловленных зверьков, экз. |
Экз. на 10 канавко-суток |
Доля вида в уловах, % |
|
|
Рыжая полевка |
5845 |
1,55 |
79,5 |
871 |
1,1 |
44,0 |
|
Красная полевка |
180 |
0,05 |
2,5 |
17 |
0,02 |
0,9 |
|
Темная полевка |
1123 |
0,31 |
15,3 |
298 |
0,38 |
15,0 |
|
Полевка-экономка |
65 |
0,02 |
0,9 |
41 |
0,05 |
2,0 |
|
Лесной лемминг |
7 |
0,002 |
0,1 |
162 |
0,21 |
8,2 |
|
Лесная мышовка |
131 |
0,03 |
1,7 |
589 |
0,75 |
29,9 |
|
Всего |
7357 |
1,962 |
1000,0 |
1978 |
2,51 |
100,0 |
Таблица 2
Изменение численности и соотношения фоновых видов мышевидных грызунов в коренных и трансформированных рубками леса биотопах
|
Биотоп |
Рыжая полевка |
Темная полевка |
Полевка-экономка |
||||||
|
I |
II |
III |
I |
II |
III |
I |
II |
III |
|
|
А. Учеты на ловушко-линиях: |
|||||||||
|
Сосняки зеленомошные, спелые |
2,8 |
60 |
94 |
0,14 |
2,5 |
5 |
0 |
0 |
0 |
|
Ельники зеленомошные, спелые |
2,8 |
62 |
98 |
0,02 |
0,4 |
0,7 |
0 |
0 |
0 |
|
Открытые вырубки |
1,9 |
22 |
37 |
2,3 |
26 |
45 |
0,8 |
8 |
16 |
|
Молодняки 6–20 лет |
2,9 |
39 |
66 |
10 |
14 |
23 |
0,4 |
5 |
9,1 |
|
21–40 лет |
2,0 |
72 |
100 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Вторичные древостои 50–70 лет |
2,1 |
43 |
92 |
0,06 |
1 |
2,6 |
0,1 |
3 |
4,4 |
|
Семенные куртины |
2,3 |
58 |
96 |
0,1 |
2 |
4,2 |
0 |
0 |
0 |
|
Недорубы |
3,8 |
55 |
89 |
0,4 |
5 |
9,4 |
0 |
0 |
0 |
|
Экотоны (лес-вырубка) |
4,7 |
39 |
55 |
2,9 |
25 |
34 |
0,9 |
8 |
10,6 |
|
Б. Учеты ловчими конусами |
|||||||||
|
Сосняки зеленомошные, спелые |
2,2 |
40 |
95 |
0,6 |
10 |
26 |
0 |
0 |
0 |
|
Ельники зеленомошные, спелые |
1,0 |
27 |
100 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Открытые вырубки |
0,1 |
2 |
5 |
1,4 |
32 |
76 |
0,1 |
3 |
5,4 |
|
Молодняки 6–20 лет |
0,5 |
9 |
40,6 |
0,6 |
14 |
49 |
0 |
0 |
0 |
|
20–30 лет (смешанные) |
2,0 |
45 |
77 |
0,6 |
10 |
23 |
0 |
0 |
0 |
|
30–40 лет (хвойные) |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Вторичные древостои 50–70 лет |
1,1 |
24 |
50 |
0 |
0 |
0 |
0,2 |
4 |
9,1 |
|
Семенные куртины |
1,6 |
23 |
59 |
0,8 |
14 |
29,6 |
0 |
0 |
0 |
|
Недорубы |
0,7 |
12 |
70 |
0,2 |
3 |
20 |
0 |
0 |
0 |
Примечание. I – численность (экз. на 100 ловушко-суток – А, на 10 конусо-суток – Б), II – % от улова мелких млекопитающих, III – % от улова мышевидных грызунов.
вырубками. Это происходит несмотря на то, что на вырубках многочисленны другие виды полевок (см. табл. 2).
Реакция рыжей полевки на концентрированную рубку хвойных лесов зависит также от породного состава вырубаемого древостоя (типа
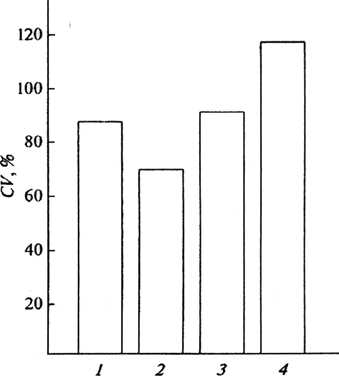
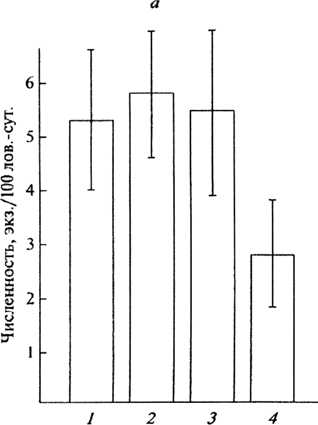
Рис. 1. Численность рыжей полевки и ее динамика в сосняках различной степени фрагментации: А – средние многолетние показатели учета (экз./100 ловушко-суток), Б – коэффициент вариации (CV, % ); 1 – заповедные территории, 2 – крупные массивы (> 30 га), 3 – небольшие массивы (6–7 га), 4 – мелкие выделы до 2 га
леса) и характера последующей сукцессии растительности. В средней тайге (Южная Карелия) при рубке высокопроизводительных сосняков и ельников и формировании злаковых типов вырубок происходит значительное снижение численности и индекса доминирования рыжей полевки. Близкий тип изменений численности рыжей полевки характерен также для вырубок в северной тайге Карелии. Он также преобладает при рубках хвойных лесов в таежных регионах к востоку от Карелии. При этом следует учитывать способ и степень очистки лесосек. На плохо очищенных вырубках численность этого вида и представленность его в уловах зверьков выше.
Большое значение имеет и близость стены леса, и наличие недорубов и семенных куртин. Заселение вырубок рыжей полевкой идет в первую очередь из недорубов, даже маленьких по площади. И в дальнейшем эти участки захламленного хвойного леса служат своеобразным «резерватом», который гарантирует сохранение популяции зверьков, особенно в годы с резкими изменениями погодных условий, увеличивающими гибель полевок в открытых стациях.
При анализе данных по изменениям численности рыжей полевки во время трехлетнего эксперимента (рис. 2) прослеживаются некоторые ее особенности, связанные с рубкой леса. Прежде всего уровень численности вида на открытых и зарастающих лесосеках все эти годы был ниже, чем на «контроле» (в коренном лесу) и участках на более поздних стадиях сукцессии. При этом амплитуда многолетних колебаний численности зверьков в молодняках и недорубах, как правило, была более резко выражена, чем в зрелых сосняках. Промежуточное положение между «контролем» и «опытом» занимают 40–50-летние древостои.
Сезонная динамика численности рыжей полевки в бесснежный период в молодняках (рис. 3) характеризуется более резким, чем в «контроле», нарастанием численности зверьков к июлю – августу и высоким ее уровнем в середине лета. К осени показатель учета рыжей полевки в мо-лодняках оказывается ниже, чем в сосняке-зеле-номошнике. Снижение же численности зверьков в молодняках в период межсезонья, возможно, связано с ухудшением экологических условий в этом биотопе в «переходные» периоды.
На открытой злаковой вырубке рыжие полевки отсутствовали в начале сезона, но в небольшом количестве появлялись к осени (молодые расселяющиеся животные).
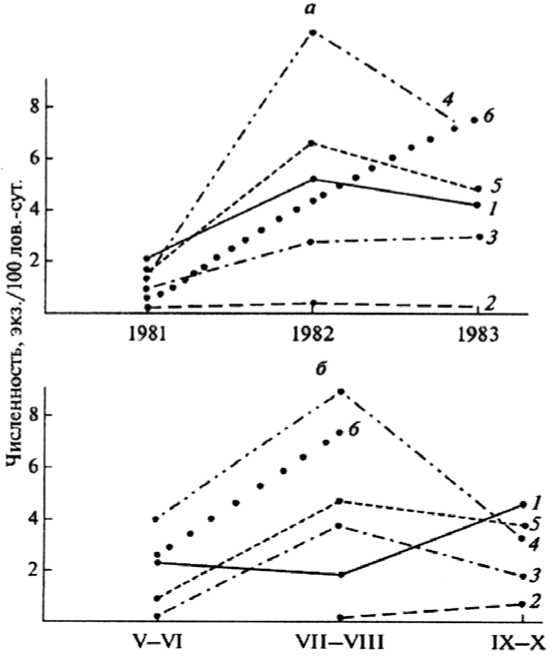
Рис. 2. Изменение численности рыжей полевки в различных биотопах по годам (а) и по средним данным в течение бесснежного периода (б): 1 – сосняк зеленомошный, 2 – открытая вырубка, 3 – молодняк 6–15 лет, 4 – молодняк 16–20 лет, 5 – древостой 50–70 лет, 6 – недорубы и семенные куртины
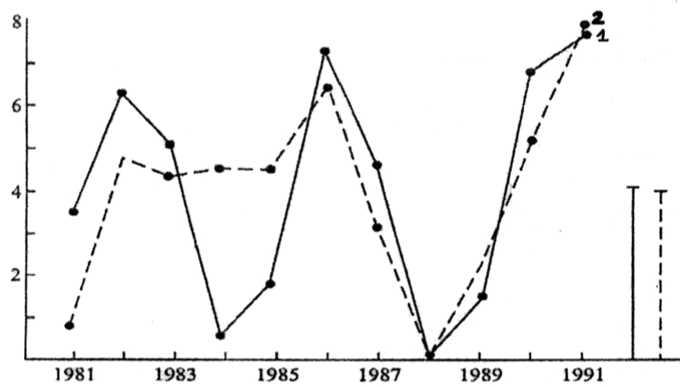
Рис. 3. Изменение численности рыжей полевки на заповедной территории (1) и в трансформированном рубками ландшафте (2)
Анализ возрастной структуры населения показывает, что с увеличением давности рубки доля зимовавших особей возрастает от 0 на вырубках до 33 % в сомкнутых молодняках (что, возможно, связано со снижением здесь зимней смертности зверьков). Сомкнутые древостои вообще характеризуются более высокой долей зимовавших рыжих полевок (17,2–33,3 % ), чем открытые вырубки и молодняки 5–10 лет (0–7,6 %). Доля участвующих в размножении самок (из числа прибылых ранних выводков) в молодняках выше, но снижение численности вида к концу сезона может указывать на повышенную смертность зверьков или перемещение части животных в коренные хвойные леса.
Статистически достоверное преобладание самцов среди прибылых зверьков отмечено только в недорубах и семенных куртинах. Очевидно, именно в эти биотопы мигрирует наиболее подвижная часть популяции, тем более, что среди взрослых особей (составляющих почти 30 % всех отловленных в недорубах рыжих полевок) преобладают самки (около 55 % ). Средняя величина выводка у рыжих полевок несколько выше в стациях с благоприятными условиями жизни в молодняках 16–20 лет, в старовозрастных сосняках, а также во вторичных смешанных и лиственных лесах 40–50 лет.
Анализ данных по другим регионам показал, что сходная реакция рыжей полевки (как и большинства остальных представителей наших лесных полевок) на сплошную рубку леса наблюдается и в других частях лесной зоны Европейской России [5], [8], [9], [10], а также в Финляндии [15] и Польше [12].
Таким образом, в первые годы после концентрированной рубки леса происходит снижение численности и доминирования рыжей полевки, особенно заметное на злаковых вырубках. В дальнейшем, по мере восстановления лесной растительности, численность вида возрастает. Лишь в жердняках (древостои возрастом 20– 40 лет) отмечен минимальный показатель численности рыжей полевки при сохранении высокого индекса доминирования в отловах грызунов.
Амплитуда колебаний численности вида в смежные годы, а также от весны к лету и осени в молодняках была несколько выше, чем на «контроле» (старовозрастные сосняки). Сохранение при рубке семенных куртин, недорубов, а также успешное лесовосстановление способствуют более быстрому заселению вырубок рыжей полевкой, особенно при формировании незлаковых типов лесосек с обилием порубочных остатков.
ЗАКЛЮЧЕНИЕ
Говоря об общих экологических последствиях наблюдающейся после рубки фрагментации коренных лесов и формирования мозаичного ландшафта, можно выделить два последовательных этапа трансформации таежных экосистем после начала интенсивной лесоэксплуатации. На первом этапе трансформации крупных массивов лесов происходит их существенная фрагментация, сопровождающаяся инсуляризацией местообитаний лесных видов животных. На втором этапе, спустя некоторое время после начала фрагментации, формируется мозаичный ландшафт, представляющий собой совокупность вырубок, молодняков, производных лиственных и смешанных лесов, а также сравнительно небольших фрагментов сохранившихся хвойных лесов.
Соответственно выделенным этапам, рассмотрение данной проблемы следует вести в двух аспектах – по пути сравнительной оценки изменений видового состава, разнообразия и численности животных в ходе фрагментации массивов хвойных (сосновых) лесов и анализа изменений в фаунистических комплексах и численности животных, происшедших после формирования массивов мозаичного, трансформированного рубками ландшафта. В Средней Европе териологические исследования в этой области чаще публикуются в виде кратких обзоров и, как правило, либо вовсе не касаются таежной зоны, либо связаны исключительно с экзотическими группами, например с летучими мышами. Работы же, выполненные в таежной Скандинавии, чаще всего носят гипотетический или обзорный характер [11], [14] и обычно не опираются на конкретный экспериментальный материал. Скорее всего, это связано и с тем, что в этом регионе не просто найти достаточно большую серию расположенных рядом, в сходных условиях массивов незатронутых рубками таежных лесов. Тем более, актуально изучение данной проблемы там, где такие массивы еще сохранились, и прежде всего в таежных сосновых лесах российской части Восточной Фенноскандии.
Анализ связанных с рубками леса характерных процессов трансформации населения доминирующей здесь рыжей полевки позволил вскрыть как внутривидовые, так и межвидовые изменения в общей массе формирующихся здесь биоценотических группировок мелких млекопитающих. Эти исследования, в частности, показали, что с сокращением размеров фрагментов хвойных лесов сокращается и количество видов, зафиксированных в среднем за год (фактически это индекс видового богатства), причем наиболее резкий переход (в 2 раза) наблюдается от крупных к небольшим (6–7 га) массивам. Последовательно растет и индекс Паркера – Бергера (доминирование наиболее многочисленного вида, в данном случае рыжей полевки), что свидетельствует о росте «несбалансированности» сообщества в пользу одного доминирующего вида. Характерно, что показатель обилия мелких млекопитающих в небольших и мелких участках сосняков также оказался примерно в 1,5 раза ниже, чем в крупных массивах. Заметные изменения наблюдаются и при сокращении размеров фрагментов от 20–30 до 6–7 га. При этом не только снижается показатель учета, но и растет коэффициент его вариации, рассчитанный за 11 лет наблюдений. Это можно рассматривать как еще одно, хотя и косвенное, свидетельство связанного с рубками снижения стабильности населения зверьков.
Симптоматично и то, что тренды многолетней динамики численности мелких млекопитающих в сосняках различной степени фрагментации в целом имеют сходную направленность. При этом следует отметить два аспекта. Во-первых, размах колебаний (особенно максимумы) даже в слабо фрагментированных сосняках (20–30 га) оказался несколько выше, чем на заповедной территории, во-вторых, подчеркнута закономерность, согласно которой дальнейшая фрагментация сосняков (до 6–7 га и особенно до 1–2 га) резко усиливает отклонения в характере динамики популяций мелких млекопитающих от общих тенденций, характерных для крупных лесных массивов. Это прежде всего проявляется в ином чередовании пиков обилия и в размахе колебаний.
Четкие различия выявляются и при сопоставлении уровней численности двух фоновых видов-содоминантов (обыкновенной бурозубки и рыжей полевки), которые вместе составляют более 90 % суммарного отлова. С сокращением размеров лесных массивов показатель обилия обыкновенной бурозубки растет, а рыжей полевки падает. Показатель же коэффициента вариации этих видов демонстрирует обратную зависимость. Характерные различия наблюдаются и в динамике численности. В крупных массивах заповедника волны популяционной динамики обыкновенной бурозубки имеют более сглаженный характер, а спады численности выражены слабее, чем в небольших по площади сосняках. Сходные тенденции в общих чертах отмечаются и для рыжей полевки, однако уровень численности этого вида в больших массивах «опытного» участка оказался несколько выше, а коэффициент вариации и размах колебаний в смежные годы – ниже, чем даже в сосняках заповедника. Это можно объяснить соседством и влиянием в целом благоприятных для вида резервных местообитаний типа молодняков и опушечных лент, характерных для мозаичного ландшафта опытного участка. На популяцию рыжей полевки это, по-видимому, оказывает стабилизирующее влияние. Дальнейшая фрагментация сосняков приводит к сокращению показателя численности рыжей полевки, причем особенно сильно (в 2 раза) в семенных куртинах сосны площадью 1–2 га. Что же касается периодов депрессии численности, то в мелких лесных массивах у обоих видов они выражены более резко и нередко принимают многолетний характер.
В сосняках разной степени фрагментации у рыжей полевки зафиксировано неполное совпадение волн динамики численности. В основном это касается пиков, но в некоторых случаях – и периодов депрессии численности. Таким образом, фрагментация сосняков в целом приводит к обеднению видового состава, сокращению численности видов, усилению нестабильности населения и упрощению структуры сообществ мелких млекопитающих. Максимальная степень фрагментации (сокращение размеров участков до 1–2 га) приводит к формированию довольно нестабильных и упрощенных группировок с 1–2 абсолютно доминирующими видами. Характерно, что при сохранении собственной структуры даже мелких массивов сосняков (ярусность, сомкнутый полог, специфичный напочвенный покров) массового заселения семенных куртин сосны зверьками с окружающей территории молодняков не наблюдалось. В итоге острова сосняков существенно отличаются по составу населения и численности мелких млекопитающих от смежных территорий.
Доминирующая повсеместно рыжая полевка несколько легче, чем землеройки, переносит дробление массивов спелых сосняков, хотя изменения все же значительны. Характерно, что, как и в случае с землеройками, заселения небольших по площади участков сосняков другими видами полевок (темная, экономка) не наблюдалось, хотя они и присутствовали на прилегающих территориях производных лесов. Лишь однажды в небольшом массиве сосняка-черничника площадью всего 2 га удалось обнаружить изолированное поселение лесного лемминга – вида, известного своим тяготением к крупным массивам коренных хвойных лесов Фенноскандии.
Kurhinen Yu. P., Forest Institute of the Karelian Research Center of the Russian Academy of Sciences (Petrozavodsk, Russian Federation)
THE BANK VOLE ( CLETHRIONOMYS GLAREOLUS SHREB.) IN INDIGENOUS AND ANTHROPOGENIC LANDSCAPES OF EASTERN FENNOSCANDIA
Список литературы Рыжая полевка (Clethrionomys glareolus Shreb.) в коренных и антропогенных ландшафтах Восточной Фенноскандии
- Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 244 с.
- Ивантер Э. В. Экологическая структура и динамика населения мелких млекопитающих Приладожья//Фауна и экология млекопитающих Северо-Запада СССР. Петрозаводск, 1983. С. 72-99.
- Ивантер Э. В., Курхинен Ю. П. Влияние антропогенной трансформации лесных экосистем Восточной Фенноскандии на популяции малой и средней бурозубок//Принципы экологии. 2014. № 2. С. 21-25. DOI: 10.15393/j1.art.2014.3561.
- Ивантер Э. В., Макаров А. М. Территориальная экология землероек-бурозубок. Петрозаводск, 2001. 271 с.
- Керзина М. Н. Влияние вырубок и гарей на формирование лесной фауны//Роль животных в жизни леса. М.: Изд-во МГУ, 1956. С. 21-304.
- Курхинен Ю. П. Воздействие сплошных концентрированных рубок на кормовые ресурсы и численность растительноядных млекопитающих//Влияние хозяйственного освоения лесных территорий Европейского Севера на население животных. М.: Наука, 1987. С. 18-31.
- Курхинен Ю. П., Данилов Ю. П., Ивантер Э. В. Млекопитающие Восточной Фенноскандии в условиях анетропогенной трансформации таежных экосистем. М.: Наука, 2006. 206 с.
- Наумов С. П., Руковский Н. Н. Влияние рубки леса и его возобновления на полевок рода Clethrionomys и Microtus//Новые проблемы зоологической науки. Ставрополь, 1979. Ч. 2. С. 303-306.
- Попов И. Ю. Структура и динамика населения мелких млекопитающих в связи с сукцессиями растительности в Европейской тайге: Автореф. дис.. канд. биол. наук. М., 1998. 27 с.
- Турьева В. В. Фауна мышевидных грызунов различных типов леса и ее изменения под влиянием рубок//Труды Коми филиала АН СССР. Вып. 4. 1956. С. 112-115.
- Andren H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A revie//Oikos. 1966. Vol. 71. P. 355-366.
- Bonach F., Kazakewicz M. Tentative comparison of small mammals communies in poor pine stand of various//Bulletin of Polish Academy of Sciences. 1980. Vol. 28. P. 43-48.
- Glowacinski Z. Some ecological parameters of avian communities in the successional series of a cultivated pine forest//Bulletin of Polish Academy of Sciences. 1979. Vol. 27. P. 169-177.
- Hansson Z. Landscape ecology of boreal forest//Trends in ecology and evolution. 1992. Vol. 7. P. 299-302.
- Henttonen H., Kaikusalo A., Tast J., Viitala J. Interspecific competition between small rodents in Subarctic and boreal ecosystems//Oikos. 1977. Vol. 29. P. 581-590.
