Самоподобие изменения некоторых параметров флоры

Автор: Иванова А.В., Костина Н.В., Сенатор С.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Оригинальные статьи
Статья в выпуске: 3 т.23, 2014 года.
Бесплатный доступ
Показано подобие изменения различных параметров флоры [таксономической, биоморфологической и флорогенетической (на примере адвентивных видов) структуры] в зависимости от увеличения площади на примере двух территорий. Сформулированы выводы о флористических особенностях этих территорий по компонентам флоры.
Кривая "число видов - площадь", растительное сообщество, локальная флора, конкретная флора, самоподобие, самарско-ульяновское заволжье
Короткий адрес: https://sciup.org/148314939
IDR: 148314939 | УДК: 581.9
Текст научной статьи Самоподобие изменения некоторых параметров флоры
Анализ флористического разнообразия и флористической структуры на территориях разного уровня – от локальных (флора окрестностей географического пункта) и региональных флор (бассейн реки, административная единица) до флористических царств - подразумевает рассмотрение целого ряда параметров. В ботанических исследованиях эти параметры основываются на характеристиках, где базовым понятием служит ВИД. Каждый вид имеет таксономическую (систематика), биоморфологическую (шкала жизненных форм), эколого-ценотическую (шкала приуроченности к фитоценозу), флорогенетическую (происхождение) и прочие принадлежности.
Одним из методов выявления флористического разнообразия и его специфики на любой исследуемой территории является изучение зависимости возрастания числа видов от увеличения размеров площади (кривая «число видов - площадь»). Факт, что рост количества встреченных видов происходит медленнее, чем идет увеличение площади, известен давно. Описание этой зависимости приводится в работах Х. Уотсона (1859), О. Аррениуса (1921),
Г. Глизона (1922) и многих других зарубежных и отечественных исследователей (обзор работ приведен, например: Гелашвили и др., 2004, 2013). В начале хода кривой «виды-площадь» наблюдается резкое возрастание числа видов, замедляющееся при увеличении площади. В дальнейшем, за исключением единичных находок, прирост видов не наблюдается, что соответствует почти горизонтальному направлению хода кривой (так называемая «мертвая зона» или «плато»).
Большой фактический материал по форме вышеуказанной кривой для характеристики разных типов растительности накоплен фитоценологами, выявлены оптимальные размеры пробных площадок описания, основываясь на представлении о «минимум-ареалах» (Миркин, Розенберг, 1978; Миркин, 1986; Миркин, Наумова, 1998; рис. 1). Кривая «виды-площадь» имеет одинаковый вид и для тундры и для луга и для тропического леса (Розенберг и др., 1999, с. 242; Розенберг, 2013, с. 110). В водных сообществах накопление числа видов в зависимости от увеличения площади выглядит схожим образом (рис. 2).
Очевидно, что в зависимости от экологических условий и исторического развития территории, уровень видового богатства (число видов) в отдельных растительных сообществах различен. Например, для пустынной степи он значительно меньше, чем для луговой (см. рис. 1). Следовательно, и число видов на единицу площади в различных растительных сообществах неодинаково (см. Онипченко, 2013, с. 276).
Благодаря развитию понятия «конкретная флора», предложенного А.И. Толмачевым, и внедрению соответствующего метода (Толмачев, 1974, 1986; Юрцев, 1975; Ребристая, 1987), появилась возможность оценить полученные результаты, в том числе по построенным кривым «виды-площадь» (рис. 3).
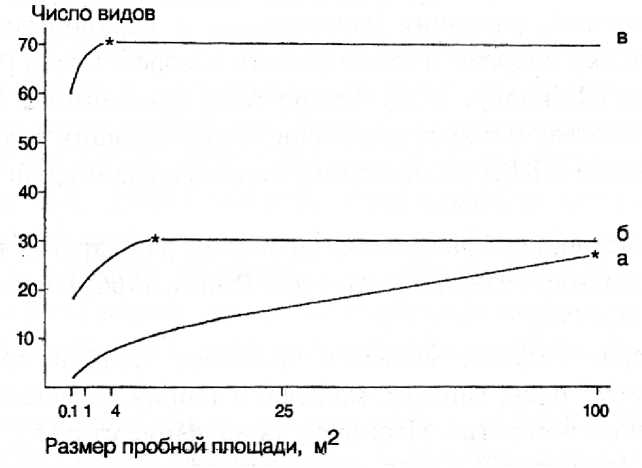
а – пустыня, б – пустынная степь, в – луговая степь,
* - точка кривой, соответствующая минимальному ареалу
Рис. 1. Кривые «число видов/площадь» (Миркин, Наумова, 1998, с. 157)
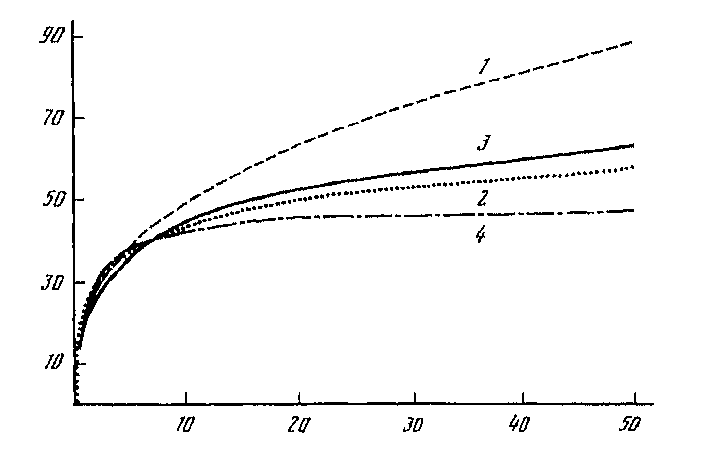
Цифрами обозначены кривые, построенные с помощью моделей:
-
1 – степенной, 2 – линейной логарифмической, 3 – нелинейной логарифмической, 4 – гиперболической.
По оси абсцисс – площадь местообитания, м2, по оси ординат – число встреченных видов.
Рис. 2. Зависимость видового богатства от площади местообитания для прибрежных фитоценозов водорослей-макрофитов зал. Посьета (Японское море) ( Кафанов, Кудряшов, 2000, с. 95)
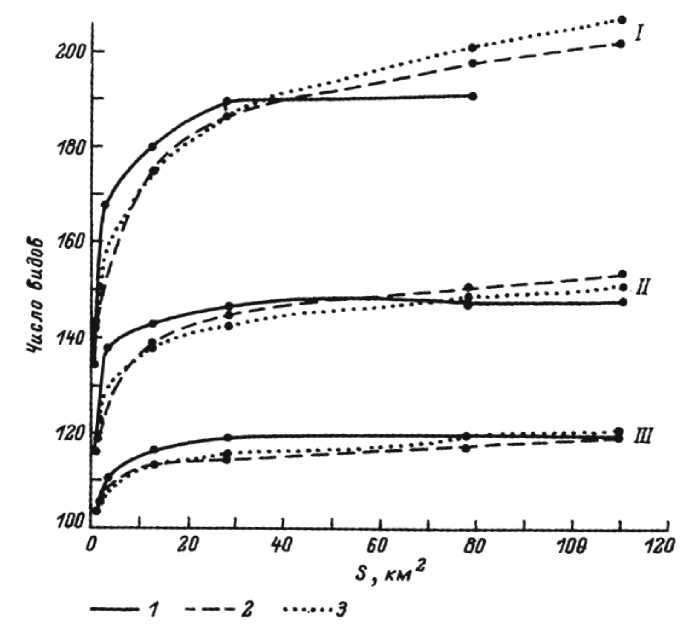
Конкретные флоры:
I - Лаптаяха (южные гипоарктические тундры),
II – Матюйяха (северные гипарктические тундры), III – Верхний Там-бей (арктические тундры)
Рассчитано:
-
2 – по уравнению Аррениуса, 3 – по уравнению Глизона
Рис. 3. Эмпирическое (1) и расчетное (2, 3) число видов на участках различной площади в пределах конкретных флор (Ребристая, 1987, с. 88)
Применение метода конкретных флор заключается в тщательном изучении некоторого(-ых) ключевого(-ых) участка(-ов). После охвата всех местообитаний с характерными для них сообществами, рост числа видов в исследуемой конкретной флоре замедляется.
Кривая «число видов - площадь» служит основой для определения таких важных характеристик, как уровень видового богатства или разнообразия, «минимум-ареал», «максимум - ареал» для отдельных сообществ, или же конкретных флор. Момент замедления соответствует «минимум - ареалу», а появление новых видов растений или исчезновение ранее регулярно встречавшихся, сигнализирует о достижении «максимум - ареала», то есть, полного охвата видового состава изучаемого сообщества или конкретной флоры.
Таким образом, количество видов, характерных для того или иного сообщества, весьма ограничено. Для конкретной флоры ограничено количество экотопов, следовательно, и растительных сообществ.
В случае увеличения площади, когда осуществляется переход из одного растительного сообщества в другое, наблюдается прирост видов и кривая «виды - площадь» приобретает ступенчатый вид (отступление 1).
Отступление 1
В геоботанических исследованиях обследованная площадь
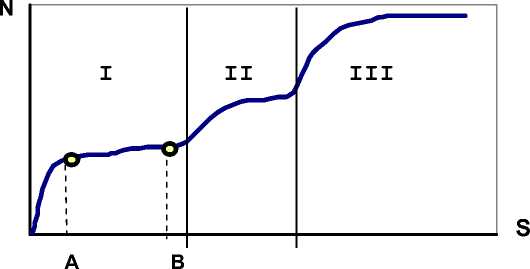
Кривая зависимости числа видов ( N ) от площади ( S ) с переходом в соседнюю "зону" ( I ^ II → III ). A - площадь "минимум-ареала", B - площадь "максимум-ареала"
измеряется в десятках и сотнях квадратных метрах и под соседней "зоной" понимается переход в другое растительное сообщество .
Во флористике размер участка составляет десятки и сотни квадратных километрах, а под соседней "зоной" понимается переход в другую конкретную флору.
При переходе из одной природно-климатической зоны в другую величина изучаемой площади увеличивается на по- рядок.
Таким образом, на разных масштабах площади имеем однотипную картину хода кривой зависимости числа видов от площади (масштабная инвариантность).
При исследовании конкретных флор с увеличением площади имеет место накопление не только (и не столько) отдельных видов, но и характерных сообществ. Если при дальнейшем наращивании площади наблюдается значительное увеличение числа видов, это свидетельствует о появлении экотопов, типичных для другой конкретной флоры. Существенный подъем кривой со- ответствует появлению не встреченных ранее видов, что свидетельствует о переходе в другую «флористическую зону» (Иванова и др., 2006), то есть в другую конкретную флору (отступление 2).
Флористические особенности территорий выявляются при анализе семейственных и родовых спектров. Иногда вычисляются показатели систематического разнообразия: среднее число видов в семействе, среднее число видов в роде и среднее число родов в семействе. Эти соотношения принято включать в основу рассуждений о степени флористического богатства рассматриваемой территории или ее флористическом разнообразии. Флористическое разнообразие любой территории напрямую зависит от экологического, исторического и антропогенного факторов. Как правило, если наблюдается «высокое» флористическое богатство, например, по числу видов, то и показатели систематического разнообразия находятся на «высоком» уровне.
Отступление 2
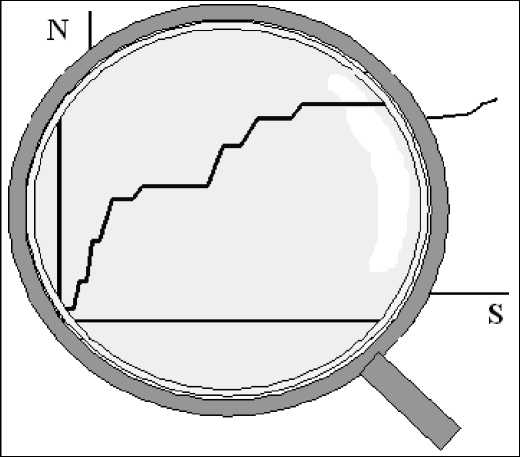
При изучении конкретных флор кривая "виды - площадь", позволяет заметить, что любая "ступень" состоит из более мелких "ступенек", каждая из которых отражает прибавление одного или нескольких типичных сообществ с характерными для них видами, а также видами, не встреченными ранее.
Заметим, что, в пределах одной "зоны" (сообщество, конкретная флора, природная зона) по мере увеличения площади ступеньки становятся ниже и длиннее.
Таким образом, определение однородности участка территории по кривой "виды-площадь" требует веских обоснований: достоверное различие между высотой "ступеней" и "ступенек".
Примечание: здесь имеем наглядное отражение принципа дискретной континуальности или непрерывной дискретности.
Показательным является пример для флор Курильских островов (Шмидт, 1980, с. 93-117; рис. 4). Полученные кривые зависимостей показателей флористического богатства (рис. 4а) и систематического разнообразия (рис. 4б) от географической широты демонстрируют определенное самопо-добие 1.
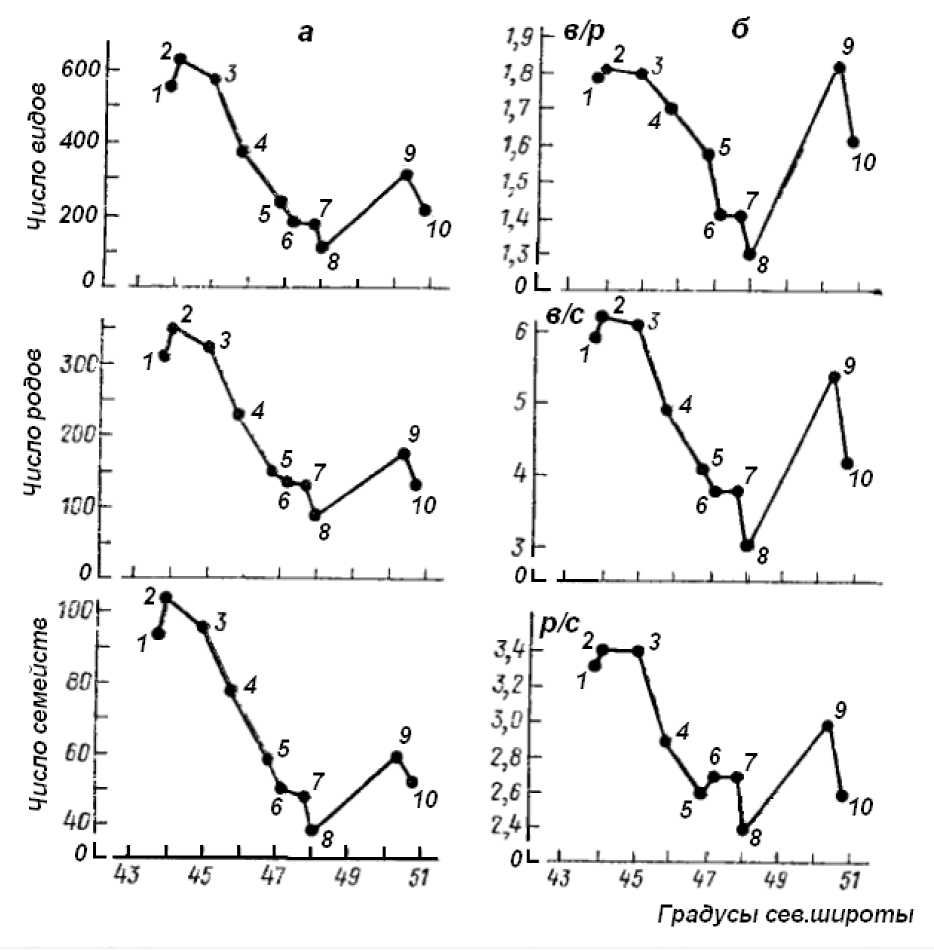
в/р - среднее число видов в роде, в/с - среднее число видов в семействе, р/с - среднее число родов в семействе.
Рис. 4. Зависимость флористического богатства (а) и показателей систематического разнообразия (б) флор Курильских островов от географической широты
(Шмидт, 1980 : 95)
Зависимость числа видов от площади для любых биологических сообществ имеет похожий вид. Она может быть аппроксимирована различными функциями: степенной, логарифмической, логистической, гиперболической и др. (Шитиков и др., 2012 : 54; рис. 2). Подбор функции зависит от целей и пристрастий исследователя.
Особой «популярностью» пользуется степенная зависимость: N = ASZ, где N - количество встреченных видов, S - площадь, А, Z - константы (отступление 3).
В настоящее время большинство ботанических работ (трудов) включает в свое содержание таксономический, биоморфологиче-ский, ареалогический эколого-ценотический, анализ флор, а также дается оценка процессов синантропизации и адвентизации территории. Уделяется внимание эндемичным, реликтовым, редким и охраняемым видам.
Рассмотрим изменения различных параметров флоры (таксономических, биоморфологиче-ских и флорогенетических) в зависимости от увеличения площади на конкретных примерах.
Отступление 3
В литературе по сравнительной флористике Z принято называть показателем пространственного разнообразия (по уравнению Аррениуса), показателем различия (по Престону). При этом следует не забывать о различиях в технике расчета этих показателей (Малышев, 1975). Согласно же фрактальной теории (Мандельброт, 2002) Z - фрактальная размерность (Федер, 1991; Гелашвили и др., 2013).
Фрактальность проявляется в таксономической структуре живых организмов, как в систематике, так и для отдельных сообществ и их совокупности какой-либо территории (Гелашвили и др., 2004, 2008, 2009, 2010, 2013).
В качестве «подопытных кроликов» были взяты две территории регионального уровня, находящиеся в одной природно-климатической зоне:
-
• бассейн реки Сок, представляющий собой преимущественно единый ландшафтный район;
-
• произвольно выбранная «прямоугольная» область, включающая части трех ландшафтных районов: Сокского, Мелекеско-Ставропольского и Ахтай-Майнского (Физико-географическое..., 1964), или Нижнесо-кского, Кондурчинского, Черемшано-Сокского (Чибилев, Дебело, 2006).
Обе изучаемые территории расположены в Самарско-Ульяновском Заволжье (рис. 5), характеризуются антропогенной нарушенностью, включают лишь «островки» естественной растительности (распаханность земель более 57%), обладают флористической неоднородностью, которая обусловлена историческими и ландшафтными (в том числе эдафическими) особенностями. Однако «прямоугольная» область имеет более высокий уровень сельскохозяйственной освоенности и плотность населения – 23,2 чел/км2. В то время, как на территории бассейна р. Сок плотность населения несколько ниже – 14,6 чел/км2.
В рамках определений конкретной флоры, согласно Толмачеву-Юрцеву 2, флора бассейна реки Сок, скорее всего, представляет одну конкретную флору, а флора «прямоугольной» области – «смешанную» флору трех ландшафтных районов.
Основой для построения кривых «виды-площадь» послужили списки локальных флор 3. Под локальной флорой нами понимается совокупность видов растений небольшого участка местности, например, окрестностей населенного пункта, озера, горы, урочища и т.д.
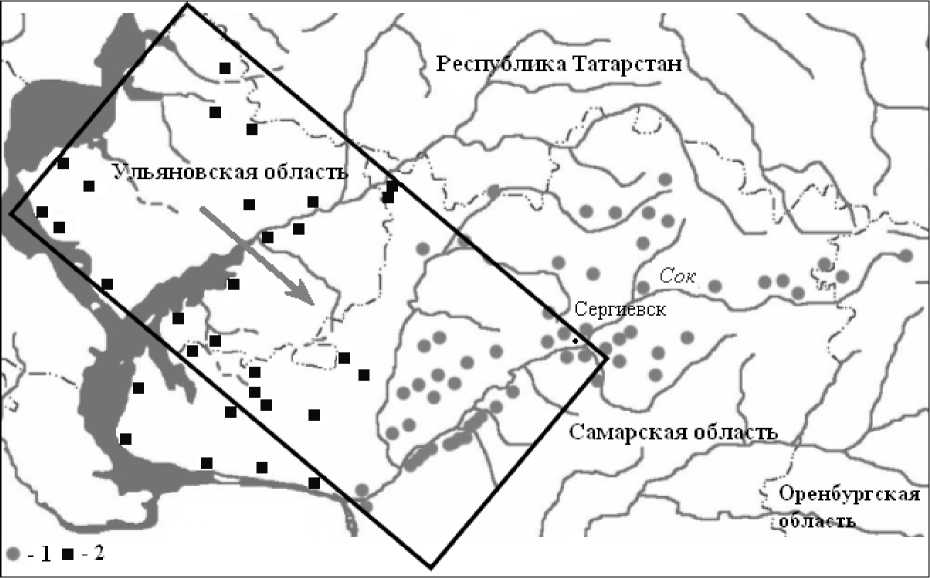
1 –локальные флоры бассейна реки Сок; 2 – локальные флоры прилегающей территории
Рис. 5. Картосхема расположения площадок флористических описаний
Базируясь на исходных описаниях локальных флор, содержащих списки видов, мы понимали, что:
во-первых , степень изученности каждой локальной флоры различна (от неоднократных посещений в течение нескольких лет и в разные периоды вегетационного сезона до одноразовых визитов), а списки описаний отражают только «выборку» из реально существующей флоры местности;
во-вторых , отдельное описание, как правило, охватывает только некоторый набор экотопов и экотонов, связанных с обследованным участком, но не включает виды полного набора экотопов, характерного для всей рассматриваемой территории;
в-третьих , месторасположение локальных флор в силу субъективных и объективных причин не представляет собой регулярную структуру.
Тем не менее, общая пространственная картина по имеющимся описаниям (мы надеемся) дает адекватную оценку распространения видов растений на изучаемой территории.
Исходя из вышеизложенных свойств рассматриваемых территорий, при построении зависимостей учитывалось следующее допущение: предполагалось, что в ближайшей окрестности изученных локальных флор встречаются или произрастали бы (в случае занятости сельскохозяйственными угодьями, рекреационными и другими антропогенно трансформированными территориями) те же виды растений.
Построение кривых «виды-площадь» происходило следующим образом. Изучаемые территории произвольно разделялись на соприкасающиеся между собой полосы (отступление 4). Площадь увеличивалась путем прибавления следующей полосы, а списки локальных флор, территориально принадлежащие к получаемой зоне, объединялись.
Заметим, что для предварительной оценки выявления нескольких флористических районов или для доказательства существования только одного района, метод выделения полос принципиального значения не имеет и особо не влияет на полученный результат.
Уточним, что нами использовалась квадратная сетка (площадь одного квадрата - 100 км2) и карта масштабом 1:2 000 000.
Отступление 4
При выделении отдельных полос можно использовать самые разные методы разбивки территории. Один из них - по градусной сетке (согласно географической широте и долготе), что позволяет выявить изменения флоры, в том числе по широтному и долготному градиенту. Выделение полос с учетом ландшафтных и экологических особенностей требует более тщательного вычисления площадей (см. отступление 5), что несложно сделать с помощью ГИС-технологий. Простым из методов является использование квадратной или прямоугольной сетки, что в дальнейшем без особых затрат позволяет приближенно определить площадь выделенной зоны. Преимущество такого подхода - возможность накладывать сетку как с привязкой к географическим координатам, так и в любой другой ориентации. Также несложно подсчитать площадь, если выделять полосы по радиальному принципу (окружности в круге или дуги в секторе).
Отступление 5
Имеется в виду определение площади по карте, т.е. в проекции "вид сверху". Точное вычисление площади, занимаемой растительным покровом, - непростая задача, т.к. любая изучаемая территория не представляет собой идеально ровную поверхность, т.е. имеет фрактальную размерность > 2.
Нами анализировалось не только изменение числа таксонов при увеличении площади (табл. 1). Также рассматривались зависимости в отношении многолетних трав, адвентивных видов и видов лидирующего семейства ( As-teraceae ). Отметим, что изменение числа видов в отдельных семействах головной части спектра и общего количества видов при увеличении площади происходит сходным образом (Иванова, 2013).
Параметры рассматриваемых флор
Таблица 1
|
Параметры |
Сокский бассейн 11870 км2, 60 лф |
«Прямоугольная» область 15300 км2, 55 лф |
|
Число семейств |
101 |
114 |
|
Число родов |
436 |
522 |
|
Число видов |
980 |
1253 |
|
Число адвентивных видов |
137 (13,9%) |
209 (16,7%) |
|
Число многолетних трав |
651 |
786 |
|
Число видов семейства Asteraceae |
159 |
179 |
Список литературы Самоподобие изменения некоторых параметров флоры
- Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С., Якимов В.Н., Шурганова Г.В. Степенной закон и принцип самоподобия в описании структуры сообществ // Поволж. экол. журн. 2004. № 3. С. 227-245.
- Гелашвили Д.Б., Якимов В.Н., Иудин Д.И. и др. Мультифрактальный анализ видовой структуры сообществ млекопитающих Нижегородского Поволжья. Экология. 2008. № 6. С. 456-461.
- Гелашвили Д.Б., Иудин Д.И., Солнцев Л.А. и др. Мультифрактальный анализ видовой структуры сообществ гельминтов мелких млекопитающих Самарской Луки // Докл. АН. 2009. Т. 427, № 5. С. 703-706.
- Гелашвили Д.Б., Якимов В.Н., Иудин Д.И. и др. Фрактальные аспекты таксономического разнообразия // Журн. общ. биол. 2010. Т. 71. С. 115-130.
- Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С., Якимов В.Н., Солнцев Л.А. Фракталы и мультифракталы в биоэкологи. Н.Новгород: Изд-во Нижегор. гос. ун-та, 2013. 370 с.
