Сдвоенное гнездование зимородка Alcedo atthis

Автор: Котюков Ю.В.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 17 т.6, 1997 года.
Бесплатный доступ
Птицы, фауна, окский заповедник, среда обитания, гнездование
Короткий адрес: https://sciup.org/140148684
IDR: 140148684
Текст статьи Сдвоенное гнездование зимородка Alcedo atthis
Величина кладки и количество кладок за сезон размножения — важные демографические параметры популяций птиц. Эти показатели определяются как физиологическим состоянием самки, так и внешними факторами, в частности погодой и уровнем плотности населения и выживаемости в популяции. Высокая ежегодная смертность у обыкновенного зимородка Alcedo atthis, превышающая и у молодых, и у взрослых 76% (Morgan, Glue 1977; Bezzel 1980), компенсируется высокой плодовитостью. Величина кладки — наименее варьирующий демографический показатель у этого вида. Поэтому его плодовитость определяется прежде всего количеством кладок в течение одного гнездового сезона. Многочисленными исследованиями подтверждено многократное гнездование зимородка, причем во всех изученных популяциях преобладали самки, имеющие 2-3 кладки в сезон (Kumari 1939; Bezzel 1980; Zoller 1980; Подольский 1982; Котюков 1986). Очередная (2-я или 3-я кладка) начинается, как правило, за 10-20 сут до вылета птенцов предыдущего выводка, т.е. когда последним исполняется 4-17 сут (Нумеров, Котюков 1984), и в редких случаях — после вылета птенцов. Автором отмечен случай последовательной откладки и попеременного насиживания яиц одной самкой в двух разных гнездах.
Материал для настоящего сообщения собран в июне 1989 во время экспедиционного обследования р. Пры (левый приток Оки, Рязанская обл.) в верхнем и среднем течении, ежегодно проводимого Центральной орнитологической станцией (Окский заповедник). Условия гнездования и размещения гнезд зимородка здесь сходны с таковыми в нижнем течении Пры, где проводится многолетнее стационарное популяционно-экологическое исследование этого вида, основанное на полном учете нор и кольцевании размножающихся птиц и птенцов.
Взрослых зимородков отлавливали и метили по методике, описанной ранее (Гуревич и др. 1978). Яйца для измерения доставали через раскоп, проделываемый сзади гнездовой камеры. После окончания работы раскоп тщательно заделывали. Все измерения проведены одним человеком с помощью штангенциркуля с точностью до 0.05 мм. В работе использован показатель “индекс удлиненности яиц”: V = 100(£ - D^/D; где L — длина, D — диаметр яйца (Костин 1977).
В ходе обследования во второй половине дня 22 июня 1989 на левом берегу Пры в 5 км выше с. Деулино (Рязанский р-н) нашли нору зимородка (№ 3, см. рисунок), в которой поймали самца, насиживавшего кладку из 10 яиц. Так как столь большая кладка встречена нами впервые за 14 лет изучения зимородка, мы предприняли попытки отловить самку. В течение нескольких часов до наступления темноты у норы наблюдали только самца. Он же дважды пойман в норе и насиживал кладку ночью. Утром следующего дня, в 740, в норе был пойман самец. Наблюдения продолжили. В течение 2 ч самец трижды залетал в нору и насиживал по 30-40 мин, в перерывах охотился на ближайшей старице в 70-100 м от гнезда и дважды улетал на 5-8 мин вверх по течению реки. В 1000 снизу по течению прилетела самка и села рядом с самцом, отдыхавшим в прибрежных зарослях ивняка в 50 м от гнезда. После короткой переклички оба партнера пересели на ближнюю к норе Присаду, и вскоре самка залетела в гнездо. Самец 8-10 мин сидел возле норы, время от времени издавая тихий сигнал тревоги. Лишь после того, как он улетел вверх по течению, удалось поймать и окольцевать самку.
Вскрыв гнездовую камеру, мы обнаружили среди 10 яиц 7 сильно насиженных, дата откладки которых, судя по размерам пути и водному тесту, 31 мая (см. таблицу, кладка № 1). Другие 3 яйца были
Размеры и форма яиц обыкновенного зимородка
( L- длина, мм; D- диаметр, мм; V- индекс удлиненности, %)
|
Кладка № 1 |
Кладка № 2 |
Кладка № 3 |
|||||||
|
L |
О |
V |
L |
D |
V |
L |
D |
V |
|
|
21.5 |
18.2 |
18.1 |
21.1 |
19.0 |
11.1 |
21.7 |
18.8 |
15.4 |
|
|
22.4 |
18.3 |
22.4 |
21.7 |
18.9 |
14.8 |
22.0 |
18.3 |
20.2 |
|
|
22.0 |
18.7 |
17.6 |
22.0 |
18.6 |
18.3 |
21.6 |
18.6 |
16.1 |
|
|
21.4 |
18.7 |
14.4 |
21.4 |
18.5 |
15.7 |
||||
|
21.4 |
18.6 |
15.1 |
23.2 |
18.4 |
26.1 |
||||
|
21.2 |
18.3 |
15.8 |
22.2 |
18.5 |
20.0 |
||||
|
21.6 |
19.0 |
13.7 |
21.9 |
18.8 |
16.5 |
||||
|
X |
21.64 |
18.54 |
16.73 |
21.93 |
18.67 |
17.50 |
21.77 |
18.57 |
17.23 |
|
SE |
0.157 |
0.109 |
1.122 |
0.254 |
0.087 |
1.782 |
0.120 |
0.145 |
1.497 |
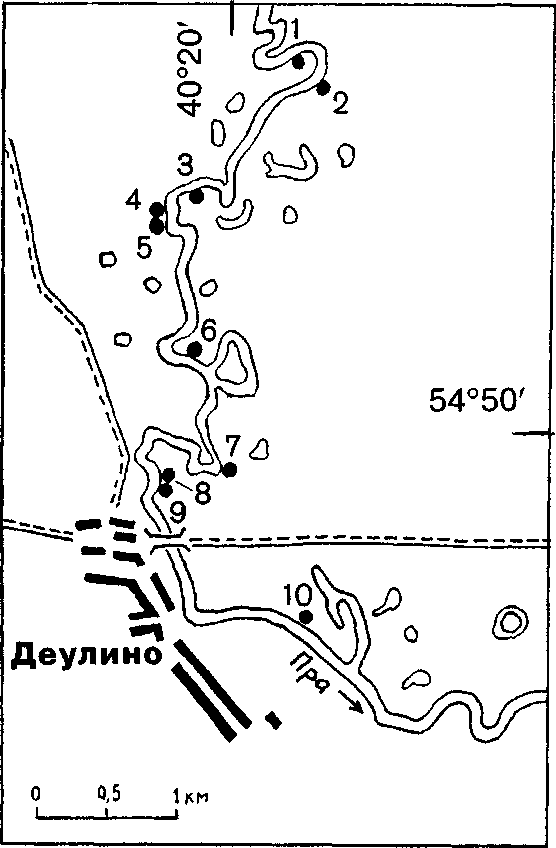
Схема размещения нор зимородка на участке реки Пры
насижены слабо —не более 2 сут (см. таблицу, кладка № 3), начало откладки —19 июня.
На правом берегу в 350 м ниже по течению найдены два гнезда другой самки: нора № 4 с кладкой из 7 яиц и нора № 5, рядом с которой держались слетки (см. рисунок). В 1.5 км ниже норы № 5 на левом берегу обнаружена нора № 6 с кладкой. В этой норе была поймана насиживающая птица, оказавшаяся самкой из норы № 3. В гнездовой камере было 7 яиц с насижен-ностью около 5 сут, т.е. начало их откладки — примерно 12 июня (см. таблицу, кладка № 2).
Таким образом, установлено попеременное насиживание одной самкой кладок в двух гнездах, отстоящих друг от друга на 1.1 км по прямой.
Поскольку одновременное обогревание самкой кладок в двух гнездах встречается у птиц довольно редко, а у зимородка от мечено впервые, представляет интерес выяснение причин столь необычного поведения. В сущности, оно сводится к ответу на вопрос: принадлежат ли кладки в обеих норах насиживающей их самке или же ее поведение — проявление альтруизма по отношению хотя бы к одному из гнезд. Предположение о том, что в описанном случае имеет место помощничество, должно быть отвергнуто, т.к. из более чем 150 видов птиц, для которых известны коммунальные ячейки с неразмножающимися помощниками, последние не участвуют в насиживании (Emlen 1982; Панов 1983). У зимородка возможность появления помощников подтверждена наблюдениями за мечеными особями (Bunzel, Druke 1986), при этом неполовозрелый самец, птенец-сеголеток из раннего выводка, кормил 22-24-сут птенцов неродственной ему пары. В отличие от молодых, для половозрелых зимородков характерно поведение, исключающее помощь в инкубации: посещение гнезда с яйцами чужой птицей всегда оканчивается гибелью кладки, т.к. чужаки прокалывают яйца клювом или повреждают их во время драки с хозяином гнезда, что доводилось наблюдать нам в Окском заповеднике и отмечено другими исследователями (Неуп 1963; Bezzel 1980). Именно из-за этой особенности поведения у зимородка со дня откладки первого яйца хотя бы один из партнеров должен постоянно находится в норе или рядом. Основанное на особенностях поведения доказательство может показаться недостаточно веским, поэтому приведем еще одно, не менее важное — результат анализа формы и размеров яиц. Из 17 найденных в двух норах яиц лишь одно, самое длинное, имело нормально-яйцевидную форму (К= 26.1%), тогда как остальные — укороченно-яйцевидную или близкую к ней форму (Исоставляет 11.1-22.4, в среднем 16.6±0.7). Выделяющееся значение V можно отнести к “выскакивающим” вариантам при оценке его по критерию Диксона (Земитис 1987): т = 0.298, п = 17. Длина и диаметр яиц всех 3 кладок незначительно отличаются друг от друга (см. таблицу). Попарное сравнение параметров яиц отдельных кладок не выявило значимых различий как средних (критерий Бейли, Р > 0.3), так и дисперсий (F-критерий, Р > 0.09). Это позволяет допускать, что кладки в обеих норах принадлежат одной самке.
Во второй норе рассматриваемой самки (нора № 6, см. рисунок) самец не был пойман, однако, судя по поведению, им не мог быть самец из норы № 3, т.к. он почти постоянно находился в норе или рядом и насиживал кладку ночью. Последнее наблюдение интересно тем, что противоречит данным автора, собранным в Окском заповеднике, и указаниям в литературе (Reinsch 1968; Bezzel 1980), согласно которым ночью обогревает кладку только самка.
Поскольку ночное насиживание самцом одной из двух синхронных кладок самки одинаково вероятно свидетельствует и в пользу моногамии, и в пользу биандрии самки, рассмотрим в доказательство нашего предположения о двубрачии самки следующую гипотетическую ситуацию. Допустим, кладки в норах № 3 и № 6 принадлежат одному самцу. Подобные случаи не представляют редкости в полигинических группировках зимородка (Котюков 1991), где самцу приходится попеременно насиживать 2-3 кладки разных самок, которые, в свою очередь, также активно участвуют в насиживании своих кладок. В рассматриваемой ситуации вскоре после завершения кладки в норе № 3 моногамный самец должен привлечь вторую самку, выкопать совместно с ней нору с тем, чтобы 12-18 июня была отложена кладка. В течение этого периода длительностью не менее 10 сут самка постоянно находится возле или в самой норе и самостоятельно не охотится. Самец значительную часть времени дневной активности вынужден тратить на кормление самки, постройку гнезда и охрану территории и партнера. Из-за недостатка времени у самца кладка в норе № 3 подолгу остается без наседки и неизбежно должна погибнуть. В действительности этого не произошло. Наси- живание, по-видимому, было достаточно плотным. Это подтверждает наше предположение о двубрачии самки.
На основании изложенного выше можно попытаться восстановить хронологическую картину размножения самки. Во второй половине мая самка образует пару с самцом и выкапывает нору длиной 57 см, где с 31 мая по 6 июня откладывает кладку из 7 яиц. Сразу после завершения первой кладки самка образует пару со вторым самцом и приступает к рытью норы в 1.1 км по прямой (2.3 км по фарватеру) от первой норы. Следует отметить, что самка, вероятно, образовала пару со вторым самцом на следующий день после завершения первой кладки, т.к. интервал между первой и второй кладками составляет 6 сут, а гнездостроение у зимородка длится 5-12, в среднем 7 дней (Гуревич и др. 1978). В готовую нору (№ 6, см. рисунок) длиной 64 см в период с 12 по 18 июня откладываются 7 яиц второй кладки, а 19-21 июня самка откладывает в своей первой норе (№ 3) еще 3 яйца в дополнение к 7 имеющимся. Таким образом, в течение 22 сут самка отложила в 3 кладках 17 яиц, масса которых составляет 172% от ее массы тела (39.6 г на 23 июня). Она эпизодически обогревала кладки в обоих гнездах.
Проанализировав ситуацию на описанном участке Пры в конце июня 1989, можно предположить, что самец, гнездившийся в норе № 6, все-таки был пойман. Этим самцом мог быть хозяин норы № 7 (23 июня — 14-сут птенцы) или нор № 8 (вылет в конце 2-й декады июня) и № 9 (23 июня — 6 яиц). В противном случае им мог быть моногамный самец, гнездившийся только в норе № 6, или же полигамный хозяин нор № 4 и № 5, или же хозяин норы №10 (23 июня — 3 яйца). Самец, гнездившийся в норе № 3, вероятно, тоже был полигамом и размножался в паре с самкой из нор № 2 (22 июня — 4 яйца) и № 1 (птенцы покинули гнездо примерно за сутки до осмотра). Подтверждением двубрачия этого самца может служить тот факт, что за время наблюдений у норы № 3 он несколько раз улетал вверх по течению (вероятно для того, чтобы покормить самку).
Столь редкое явление, описанию которого посвящено настоящее сообщение, интересно по крайней мере в трех аспектах. Во-первых, сдвоенное гнездование — эффективный способ повышения репродуктивного успеха. Судя по срокам гнездования и учитывая наложение двух последующих выводков, уместно предположить, что до конца гнездового сезона самка успеет отложить с каждым из самцов по второй кладке и выкормить птенцов. При более жестких заданных условиях, исключающих возможность второй кладки со вторым самцом, самка способна выкормить 3 выводка по 7 птенцов. В расчеты не включены три яйца, отложенные в дополнение к первой кладке в норе № 2, т.к. наиболее вероятно, что птенцы из них не 1 О Рус- орнитол. журн. 1997 Экспресс-выпуск №17
вылупятся или будут затоптаны птенцами первого выводка, которые примерно на 2 нед старше. Значительный репродуктивный потенциал многих самок зимородка, по крайней мере только “местной” популяции, можно считать характерной особенностью гнездового сезона 1989 года. Например, в Окском заповеднике в этот год 5 из 26 (19.2%) самок отложили по три нормальные кладки, в то время как за все предыдущие годы отмечено лишь 3 из 363 (0.8%) подобных случая. Первые кладки всех 5 самок были начаты в период с 1 по 7 мая. Если названную особенность считать определяющей стратегией для данного гнездового сезона, то последовательная откладка и одновременное насиживание яиц в 2 гнездах, вероятно, объясняется более поздним началом гнездования самки. Альтернативная гипотеза состоит в том, что для описываемой самки как индивидуума характерен высокий репродуктивный потенциал и своеобразная тактика размножения вне связи с условиями сезона и временем начала гнездования. Из-за недостаточности материала автор не может отдать предпочтения ни одной из гипотез.
Во-вторых, сдвоенное гнездование интересно с точки зрения репродуктивного поведения. Известно, что полиандрия у птиц — наиболее редкий из четырех основных типов отношений полов (Панов 1983; Моск 1983). По данным Джени (Jenni 1974), доля видов птиц мировой фауны, у которых отмечены хотя бы единичные случаи полиандрии как в природе, так и в условиях неволи, составляет менее 1%. Обыкновенный зимородок относится к числу тех немногих видов, у которых последовательная территориальная полиандрия отмечается регулярно: в Окском заповеднике в течение 15 лет зарегистрировано 43 подобных случая (Котюков 1991). В отличие от того, что было известно для зимородка ранее, описываемый случай является примером уникальной системы размножения — одновременной территориальной полиандрии. Об ее редкости свидетельствует тот факт, что до сих пор она найдена только у пятнистого перевозчика Actitis macularia, у которого самка на протяжении 2 последующих сезонов в течение короткого промежутка времени откладывала 2 кладки с разными самцами и эпизодически насиживала обе кладки (Oring, Maxson 1978). Сравнение с другими видами птиц, например, с американской яканой Jacana spinosa, здесь вряд ли уместно, поскольку у названного вида имеют место значительный обратный половой диморфизм (самки в два раза крупнее самцов) и реверсия половых ролей, а одновременная полиандрия для него — облигатный тип семейных отношений (Jenni, Collier 1972). У пятнистого перевозчика, хотя и имеется незначительный обратный половой диморфизм, но, в отличие от американской яканы, эти кулики в зависимости от места и условий сезона размножаются в составе монотус. орнитол. журн. 1997 Экспресс-выпуск №17 1 о гамных пар или полиандрических ячеек (Hays 1972; Oring, Knudson 1972; Maxson, Oring 1980). Вполне определенно, тасманийскую курочку Tribonyx mortierii, которую считают ярким примером одновре-менно-полиандрического вида без реверсии половых ролей (Ridpath 1972; Maynard Smith, Ridpath 1972) также нельзя отнести к подходящим для сравнения видам, т.к. у этой нелетающей птицы самки откладывают яйца, оплодотворенные разными самцами, в одно коммунальное гнездо.
В-третьих, в результате непрерывной откладки яиц (кладки № 2 и № 3) последовательно в два гнезда (норы № 6 и № 3), часть или все яйца кладки № 3 могли оказаться оплодотворенными самцом из норы № 6. Такой своеобразный “гнездовой паразитизм”, обусловленный сохранением спермы у самки в течение 3-31 сут, отмечен у полиандрического пятнистого перевозчика (Oring et al. 1992). Несмотря на то, что у зимородка вопрос отцовства путем исследования крови птенцов не изучен, нет оснований полностью отрицать возможность сохранения спермы в половых путях самок этого вида. Для самцов-реципиентов в подобных ситуациях существует определенный риск принять на воспитание чужое потомство.
И наконец, последнее замечание касается величины кладки у зимородка. Вторая кладка описанной самки, как и первая, содержала 7 яиц, тогда как 3 яйца третьей кладки служили продолжением кладки № 2 и были отложены с нормальным (суточным) интервалом, но в другом гнезде. Этот факт свидетельствует о том, что у обыкновенного зимородка, независимо от потенциальной плодовитости самки, величина кладки — параметр фиксированный.
Автор выражает искреннюю признательность сотруднику Окского заповедника С. В. Филяревичу за неоценимую помощь в проведении полевых исследований.
