Сегетальная растительность агрофитоценозов зерновых культур Минской области

Автор: Терещенко Светлана Сергеевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология растительных сообществ
Статья в выпуске: 1-6 т.14, 2012 года.
Бесплатный доступ
В статье с использованием синтаксономии дан обзор сегетальной растительности посевов зерновых культур Минской области.
Сегетальное сообщество, агрофитоценоз
Короткий адрес: https://sciup.org/148200876
IDR: 148200876 | УДК: 581.524
Segetal vegetation of the agrophytocenosis of grain crops of the Minsk area
In the article using syntaxonomy the survey of segetal vegetation of grain crops of the Minsk area is given.
Текст научной статьи Сегетальная растительность агрофитоценозов зерновых культур Минской области
Сорно-полевые растения являются неотъемлемым естественным компонентом агрофитоценоза, численность и состав которых находится в тесной взаимосвязи с экотопом, и формируются под влиянием целого комплекса природных и антропогенных факторов. Группировки сегетальной растительности по своим свойствам близки к сообществам естественной растительности, однако отличаются более однородным флористическим составом, что связано с нивелированием естественных условий экотопа. Тем не менее, на пахотных землях происходит формирование ценоиндикационных комплексов [1 ] , виды которых позволяют оценить экологические условия произрастания фитоценоза.
Целью наших исследований было изучение се-гетальной растительности и выявление экологических закономерностей их распределения в агрофитоценозах зерновых культур Минской области с использованием эколого-флористической классификации.
Геоботанические исследования проводились в течение 2010-2011 гг. на ключевых участках, заложенных с использованием метода эколого-фитоценотических п рофилей (ри с. 1), разработанного в лаборатории геоботаники Института экспериментальной ботаники им. В.Ф. Купревича НАН Беларуси [2 ] .
Всего было выполнено 305 полных геоботанических описаний по стандартным методикам [3, 4]. При описании сегетальных сообществ на пробных площадях 100 м2 регистрировались все виды с указанием количественного соотношения между ними. Проективное покрытие определяли по 7-балльной шкале [4]. При обработке материалов использован метод Браун-Бланке [4, 5, 6]. Названия синтаксонов даны в соответствии с «Кодексом фитосоциологи-ческой номенклатуры» [7].
Для сравнения степени сходства таксономического состава рассматриваемых сообществ в различных агрофитоценозах применен коэффициент Жаккара [8].
Для определения экологических условий использовались экологические шкалы Элленберга [9]. По выбранным факторам рассчитаны балльные эколо-
гические оценки для каждого геоботанического описания. Расчет выполнялся методом определения средневзвешенного индекса выраженности фактора по экологическим формулам. Полученные экологические показатели анализировали методом одномерной ординации (градиентный анализ), по которому синтаксоны упорядочиваются вдоль конкретных осей экологических факторов, что отражает закономерный характер их изменений [4]. Проведенный таким образом экологический анализ видов дает возможность судить об экологических особенностях сообществ, и отражает экологические режимы их местообитаний.
Синтаксономическая схема сегетальной растительности в посевах зерновых культур на территории Минской области включает 1 класс, 4 порядка, 5 союзов и 8 ассоциаций.
Класс Stellarietea mediae R. Tx., Loheyer et Preising in R. Tx. 1950
Порядок Aperetalia J. et R. Tx. 1960
Союз Scleranthion annui (Krus. & Vlieg. 1939) Sissingh in Westhoff, Dijk & Passchier 1946 Ассоциация Spergulo-Scleranthetum annuі Kuhn 1937
Ассоциация Centaureo-Aperetum spicae venti V. Solo-makha 1989
Союз Chenopodio albi-Descurainion sophiae V. et T.
Sl. et Shelyag-Sosonko in V. Sl. 1988
Ассоциация Fallopio convolvulus-Chenopodietum albi V. Sl. 1990
Порядок Polуgono-Сhenopodietalia (R. Tx. et Lohm.
1950) J. Tx. 1961
Союз Panico-Setarion Siss. in Westhoff, Dijk et Passchier 1946
Ассоциация Echinochloo-Setarietum Krus. et Vlieg.
(1939) 1940
Ассоциация Echinochloetum crusgalli V.Sl. 1988
Союз Polygono-Chenopodion Siss. W. Koch 1926 em Siss. 1946
Ассоциация Galeopsietum speciosae Krusem et
Vlieg. 1939
Ассоциация Gnaphalio uliginosae-Matricarietum perfo-ratae Gamor 1987
Порядок Eragrostietalia J.Tx. ex Poli 1966
Союз Matricario-Chenopodion albi Timár 1954 Ассоциация Matricarietum perforatae Kepczyńska 1975.
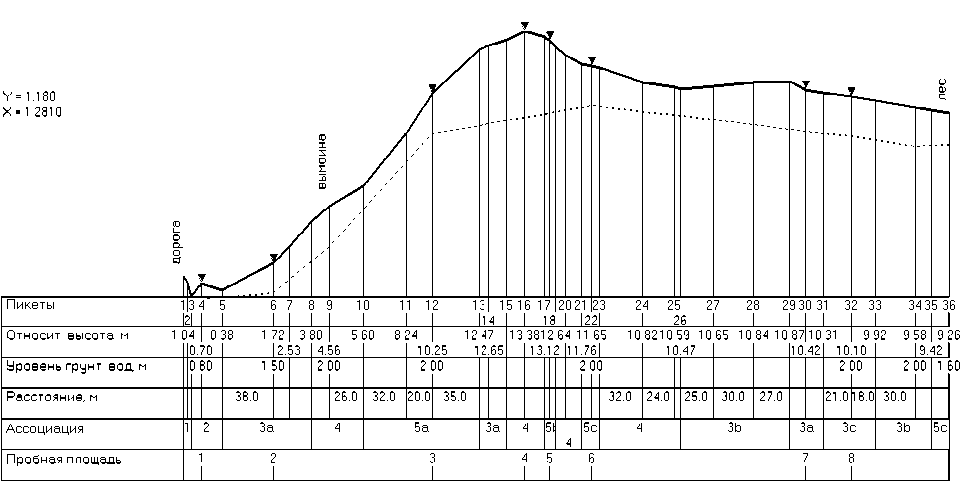
Рис. 1 . Эколого-фитоценотический профиль в агроэкосистеме на западе Минской возвышенности 0,4 км северо-восточнее д. Станюши Ивенецкого района Минской области. 2011 г. Протяженность 0,60 км.
Примечание: Ассоциации: 1 - Festucetum arundinaceae (subass. F. a. dactylidetosum glomeratae); 2 - Ranunculo re-pentis-Alopecuretum geniculati (subass. R.r.-A.g. bidentetosum tripartiti); 3 - Centaureo-Aperetum spicae-venti (subass.: a - C-A. sp.-v. elytrigietosum repentis; b - C-A. sp.-v. sonchetosum arvensis; с - C-A. sp.-v. menthetosum arven-sis); 4 - Spergulo-Scleranthetum annuі (subass. Sp.-Sc. a. agrostidetosum giganteae); 5 - Agropyretum repentis (subass.: a - A. r. cirsietosum arvensi; b - A. r. sonchetosum arvensis; c - A. r. stachietosum palustris)
Для анализа мы выбрали посевы зерновых культур (рожь, пшеница, тритикале, ячмень), которые по статистическим данным на 2010 год составляли около половины от всех посевных площадей – 575, 8 тыс. га (50,6 %), из них на долю озимых зерновых приходилось 47,7 %, яровых – 52,3 %.
Всего в посевах зерновых культур зарегистрировано 129 видов сорных растений, которые принадлежат к 84 родам, 26 семействам. Преобладают виды семейств Compositae (17,5 %), Gramineae (9,3 %) , Cruciferae (8.5 %) , что в целом характерно для синантропной растительности Беларуси [10]. Ведущие 13 семейств включают в себя 111 видов (86 %). Наличие такого высокого количества видов в сравнительно незначительном числе семейств свойственно территориям, условия развития которых могут рассматриваться как экстремальные [11]. В данном случае экстремальные условия создаются за счет агротехнических и агрохимических мероприятий.
Видовое разнообразие сорно-полевого комплекса в посевах озимых культур значительно выше, чем в яровых. Так, в посевах озимой пшеницы в среднем встречалось 24 вида сорных растений, яровой пшеницы – 17 видов, озимой ржи – 22 вида, озимой тритикале – 21 вид, яровой тритикале – 16 видов, ячменя – 15 видов. Агрофитоценозы ячменя были наиболее бедными по видовому составу. Полученные данные свидетельствуют о первостепенном влиянии эдафо-климатических условий на формирование сообществ, чем воздействие культуры. Видовое разнообразие сообществ в однородных экологических условиях определяется эдификатор- ной ролью культурных доминантов [12] и способами агротехнических приемов, применяемых для выращивания зерновых культур.
Анализ флористического состава сообществ се-гетальной растительности в районах Минской области показал, что коэффициенты сходства между группировками сообществ, относимых к одной ассоциации, варьируют незначительно. Так, для озимых культур коэффициент общности в среднем равен: в ассоциации Centaureo-Aperetum spicae venti 0,3 (посевы пшеницы); асс. Fallopio convolvu-lus-Chenopodietum albi – 0,29 (посевы пшеницы); асс. Matricarietum perforatae – 0,28 (посевы пшеницы); асс. Gnaphalio uliginosae-Matricarietum perforatae – 0,27. Наибольшее сходство видового состава наблюдается в ассоциации Centaureo-Aperetum spicae venti в посевах тритикале (0,45) и посевах ржи (0,35). Для яровых культур коэффициент однородности в асс. Fallopio convolvulus-Chenopodietum albi – 0,24 (посевы ячменя), 0,26 (посевы пшеницы); в асс. Galeopsietum speciosae – 0,28 (посевы ячменя); в асс. Echinochloo-Setarietum – 0,23 (посевы ячменя); в асс. Echinochloetum crusgalli – 0,21 (посевы пшеницы). Наибольшие различия выявлены в пределах ассоциации между сорно-полевыми сообществами озимых и яровых культур, формирующимися в сходных эдафических условиях: в асс. Fallopio convolvulus-Chenopodietum albi – 0,21 и асс. Galeopsietum speciosae – 0,18 между посевами пшеницы и ячменя; асс. Matricarietum perforatae – 0,17 между посевами озимой ржи и яровой пшеницы.
В эколого-ценотическом спектре сорного компонента агрофитоценозов зерновых культур доми- нирующей группой являются сорные виды (17,9 %), а также сорные сегетальные (10,9 %), сорные луговые (7 %), сорно-прибрежно-луговые (6,2 %), сорно-опушечно-луговые (5,4 %) и сорнорудеральные (4,7 %) растения. Лугово-опушечные (21 %), а также луговые (3,9 %) и лугово-болотные (3,7 %) виды растений занимают второе место. Это связано с тем, что луговые растения также имеют широкий экологический оптимум обитания, и, следовательно, могут входить в состав синантропных растительных сообществ. На третьем месте находятся прибрежно-сорные (11,6 %), прибрежноболотные (5,4 %) виды растений. Самый небольшой процент видов относится к культурным и интродуцированным видам (2,3 %). Отчасти такой расклад является характерным для сегетальной растительности, что определяется параметрами, характеризующими природную среду Минской области (расчлененность рельефа, распаханность территории, густота речной сети, различная кон-турность полей, близость прилегающих естественных растительных формаций и т.д.).
Флорогенетический анализ показывает, что во всех сообществах наиболее массово представлена группа апофитов: от 45,2 % в асс. Fallopio convolvu-lus-Chenopodietum albi , до 64,4 % Gnaphalio uliginosae-Matricarietum perforatae (55 % в общем), среди которых превалируют эвритопные виды с широкими ареалами как по долготному, так и по широтному спектру. Наибольшим количеством видов в ценоф-лорах ассоциаций представлена группа евроазиатских видов - 32; 45,1 %, большинство которых относится к бореальным элементам флоры. Значительна роль циркумполярных видов - 20; 28,2 %. Это свидетельствует об общности происхождения и истории формирования всей флоры Голарктики. Высоко участие европейско-западноазиатских (7; 10 %) и европейских видов (8; 8,5 %), что подтверждает тесную связь сорной флоры с флорами Европы и Азии. Значительно меньшей является группа видов европейско-западносибирского типа ареала (11 видов; 6 %).
Сегетальные сообщества являются постоянно открытыми группировками, обладающими большим числом ниш возобновления, что делает их весьма чувствительными к внедрению чужеродных видов [13]. Это объясняет высокое присутствие в составе ценофлор сообществ адвентивных видов (45 % в общем), среди которых археофитов - 74,2 % и кено-фитов - 25,8 %. Соотношение археофитов в сообществах колеблется от 28,8 % в асс. Gnaphalio uliginosae-Matricarietum perforatae до 48 % в асс. Echinochloetum crusgalli и кенофитов от 3,7 % в асс. Echinochloetum crusgalli до 10,5 % в асс. Matricarietum perforatae. Преобладают выходцы из средиземноморского и средиземноморско-западноазиатского регионов (23 вида, или 39,7 % от всей адвентивной фракции). Второй центр, оказавший влияние на формирование адвентивного компонента флоры Минской области - южные районы Евразии (8 видов; 13,8 %). Немаловажное значение в станов- лении видового состава адвентивной фракции флоры оказали североамериканские виды, занимающие 3 место (5 видов; 8,6 %), они на 100 % представлены кенофитами. Это обусловлено усилением торговоэкономических связей между Старым и Новым Светом в период XVII-XIX веков, а также сходством климатических условий, способствующих дальнейшей экспансии североамериканских видов. Особенность североамериканских видов проявляется в их высокой инвазийности и способности к натурализации.
Жизненные формы сегетальных растений в агрофитоценозах зерновых культур представлены 17 видами геофитов, 36 видами гемикриптофитов и 76 видами терофитов. В группировках сегетальной растительности независимо от культуры преобладают терофиты (от 54,8 % в асс. Gnaphalio uliginosae-Matricarietum perforatae до 80,6 % в асс. Fallopio convolvulus-Chenopodietum albi ), что характерно для агроэкосистем. Соотношение геофитов и гемикриптофитов значительно варьирует в зависимости от характера культурного доминанта. Гемикриптофиты по численности преобладают в посевах озимых культур, геофиты - в яровых. Такая закономерность отмечена при изучении соотношения жизненных форм в сообществах Spergulo-Scleranthetum annui, Fallopio convolvulus-Chenopodietum albi, Echinochloo-Setarietum, Galeopsietum speciosae, Matricarietum per-foratae.
Анализ сегетальных сообществ по шкалам экологических режимов Х. Элленберга указывает на решающее воздействие эдафических условий экотопа. Минимальные и максимальные значения позволяют установить широту экологической амплитуды выделенных группировок и выявить факторы, оказывающие наибольшее влияние на распространение сообщества, а среднее значение указывает на место данного синтаксона в общем экологическом ряду. Расчеты показателей приведены в таблице.
Согласно полученным показателям для каждого фактора построены схемы, отражающие обобщенные ряды синтаксонов в соответствии с изменением конкретного фактора.
Экологический ряд сообществ класса Stellarietea mediae в посевах зерновых культур по отношению к режиму освещения представлен следующим образом: Echinochloo-Setarietum ^ Galeopsietum specio-sae → Echinochloetum crusgalli → Fallopio convolvu-lus-Chenopodietum albi → Gnaphalio uliginosae-Matricarietum perforatae → Centaureo-Aperetum spi-cae venti ^ Spergulo-Scleranthetum annui ^ Matri-carietum perforatae .
Сорно-полевые растительные группировки при нарастании градиента влажности можно расположить в следующий экологический ряд: Spergulo-Scleranthetum annui ^ Centaureo-Aperetum spicae venti → Fallopio convolvulus-Chenopodietum albi → Echinochloo-Setarietum → Galeopsietum speciosae → Matricarietum perforatae → Echinochloetum crusgalli ^ Gnaphalio uliginosae-Matricarietum perforatae .
Таблица 1. Основные экологические показатели синтаксонов сегетальной растительности
|
Ассоциация |
Показатель экологического фактора (баллы) |
|||
|
Light (L) |
Moisture (M) |
Reaction (R) |
Nutrient (N) |
|
|
Spergulo-Scleranthetum annuі |
6,45-7,3/6,9 |
4,4-4,8/ 4,5 |
4,1-4,5 / 4,2 |
4,9-5,4 / 5,1 |
|
Centaureo-Aperetum spicae venti |
6,5-7 / 6,8 |
4,5-5,4 / 4,8 |
5,2-6,3 / 5,6 |
5,8-6,6 / 6,2 |
|
Fallopio convolvulus-Chenopodietum albi |
6,6-7,4 / 6,7 |
4,6-6,0 / 5,3 |
5-6,7 / 5,8 |
6,1-7 / 6,5 |
|
Galeopsietum speciosae |
6,4-7,2 / 6,6 |
4,6-5,8 / 5,6 |
6-6,4 / 6,2 |
6,2-6,6 / 6,4 |
|
Echinochloo-Setarietum |
6,1-6,9 /6,5 |
5-5,7 / 5,4 |
6-6,5 / 6,3 |
6,4-6,7 / 6,5 |
|
Echinochloetum crusgalli |
6,7-7,1 / 6,7 |
5,4-6,1 / 5,9 |
6-7 / 6,6 |
7,2-7,6 / 7,3 |
|
Gnaphalio uliginosae-Matricarietum perforatae |
6,6-6,9 / 6,8 |
5,7-6,4 / 6,2 |
5,6-6,3 / 5,9 |
6,4-7,0 / 6,8 |
|
Matricarietum perforatae |
6,7-7,4 / 6,9 |
5,3-6,2 / 5,8 |
5,8-6,4 / 6,0 |
6,0-6,6 / 6,3 |
Примечание. В числителе приведено минимальное и максимальное значение показателя в пределах синтаксона, в знаменателе – его среднее значение.
Приведенные выше данные свидетельствуют, что большинство сорно-полевых комплексов являются хорошо приспособленными к различным по уровню кислотности почвам. Их распределение в агрофитоценозах зерновых культур можно свести к обобщенному экологическому ряду: Spergulo-Scleranthetum annuі → Centaureo-Aperetum spicae venti → Fallopio convolvulus-Chenopodietum albi → Gnaphalio uliginosae-Matricarietum perforatae → Ma-tricarietum perforatae → Galeopsietum speciosae → Echinochloo-Setarietum → Echinochloetum crusgalli .
Содержание азота в почве является одним из определяющих факторов формирования растительности, поскольку именно плодородие почвы ограничивает развитие и распространение видов различной ценотической приуроченности. Экологический ряд сегетальных сообществ по отношению к содержанию азота в почве выглядит таким образом: Spergulo-Scleranthetum annuі → Centaureo-Aperetum spicae venti → Matricarietum perforatae → Galeopsietum speciosae → Echinochloo-Setarietum → Fallopio convolvulus-Chenopodietum albi → Gna-phalio uliginosae-Matricarietum perforatae → Echi-nochloetum crusgalli .
Таким образом, сообщества характеризуются экологическими особенностями, что связано с адаптированностью видов к специфическим условиям агрофитоценозов.
Наличие определенного комплекса сорных видов в агрофитоценозах позволяет выявить фактор влияния эдафотопа, использовать полученные сведения для ведения экологически обоснованного растениеводства.
Список литературы Сегетальная растительность агрофитоценозов зерновых культур Минской области
- Соломаха В.А., Соломаха Т.Д. О выделении ценоиндикационных комплексов сорняков пахотных земель//Ландшафтная индикация для рационального использования природных ресурсов. Москва, 1986. С. 79-81.
- Сцепановiч Я.М. Трансект-метад як аснова манiторынгу раслiнных экасiстэм (з нямецкага досведу)//Мiжнародны экалагiчны досвед i яго выкарыстанне на Беларусi. International Environmental Experience: Applications for Belarus (collected papers). Вiцебск, 2003. С. 226-230.
- Программа и методика биогеоценологических исследований. М.: Наука, 1974. 403 с.
- Миркин Б.М., Наумова Л.Г., Соломещ А.И. Современная наука о растительности. М.: Логос, 2001. 264 с.
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde. Wien-New York: Springer-Verlag, 1964. 865 s.
- Westhoff V., van der Maarel E. The Braun-Blanquet approach//Classification of plant communities. The Hague: Junk, 1978. P. 287-399.
- Weber H.E., Moravec J., Theourillat D.-P. International code of phytosociological nomenclature. 3rd editional//J. Veg. Sci. 2000. V. 11. N 5. P. 739-768.
- Мэгарран Э. Экологическое разнообразие и его измерение. М.: Мир, 1992. 184 с.
- Ellenberg H. et al. Zeigewerte von Pflanzen in Mitteleuropa//Scripta Geobotanica. 1992. 2 Aufl. S. 1-258.
- Третьяков Д.И. Роль синантропного компонента в формировании флоры Белоруссии: дис. … канд. биол. наук: 03.00.05. Минск, 1990. 365 л.
- Толмачев А.И. Введение в географию растений. Ленинград, 1974. 244 с.
- Туганаев В.В. Анализ сегетальной флоры Волжско-Камского края//Культурная и сорная растительность Удмуртии. Ижевск: Удмуртия, 1977. С. 33-35.
- Hart R. Why are biennials so few?//Amer. Natur., 1977. V. III. № 980. P.792-799.
