Селекционная оценка и генотипирование клонов плюсовых деревьев Рinus sylvestris L. в Северном Казахстане

Автор: Крекова Я.А., Чеботько Н.К., Каган Д.И.
Журнал: Лесохозяйственная информация @forestry-information
Рубрика: Лесная селекция и генетика
Статья в выпуске: 2, 2023 года.
Бесплатный доступ
Приведены результаты селекционно-генетической оценки и молекулярной паспортизации 15 клонов плюсовых деревьев Pinus sylvestris L. Объекты исследования расположены в Акмолинской обл. Северного Казахстана. Клоновое потомство плюсовых деревьев представлено 5 биотопами. Рассматривается комплексная оценка клонов в возрасте 37 лет по ряду показателей. Размещение рамет клонов при посадке по схеме 3×3 м оказало влияние на формирование древостоя в целом. Средняя сохранность 15 исследуемых клонов - 77,1 %. На обследуемых участках доля рамет без признаков ослабления составила 41,1 %. У 50,5 % рамет обнаружены значительные механические повреждения коры в нижней части стволов на высоте до 2 м, которые, предположительно, нанесены дикими животными. На первоначальном этапе проанализированы основные таксационные показатели, параметры кроны и проведено генотипирование 15 клонов. Средняя высота всех исследуемых клонов - 18,9±0,3 м, средний диаметр ствола на высоте 1,3 м - 20,3±0,4 см, средний диаметр кроны - 3,7±0,2 м. Оценка качественных признаков позволила выявить клоны с высоким качеством ствола (прямоствольность), густым охвоением и наибольшей устойчивостью: № 6, 17, 25, 30, 22, 34, 38. Установлено слабое плодоношение (от 20 до 100 шишек на одну рамету), которое обусловлено небольшими размерами крон и плодоносящего яруса. На основе изоферментного и микросателлитного анализа определены генотипы клонов плюсовых деревьев, а также частота встречаемости выявленных аллелей. Диагностические аллельные сочетания отдельных локусов были обнаружены для 73,3 % клонов. Генетическая идентификация позволила установить, что 98,7 % рамет соответствует исследуемым клонам. Несоответствие клоновой принадлежности оставшихся рамет подтверждено как при изоферментном, так и при микросателлитном анализах.
Pinus sylvestris l, клоны плюсовых деревьев, высота дерева, диаметр ствола, размеры кроны, генотип
Короткий адрес: https://sciup.org/143180080
IDR: 143180080 | УДК: 630.165.6 | DOI: 10.24419/LHI.2304-3083.2023.2.09
Selection evaluation and genotyping of clones of plus trees Pinus sylvestris L. Northern Kazakhstan
The article presents the results of selection and genetic evaluation and molecular passportization of 15 clones of plus trees Pinus sylvestris L. at the age of 37 years. The objects of research are located in the Akmola region of Northern Kazakhstan. The initial placement of the ramets during their planting (3×3 m) had an impact on the formation of the stand. The average preservation of 15 studied clones was 77,1 %. The proportion of ramets without signs of weakening was 41.1 %. During the examination, significant mechanical damage to the bark in the lower part of the trunks (no higher than 2 m in height) was found in 50 % of the ramets. At the initial stage, the main taxation indicators, crown parameters and genotyping of 15 clones were analyzed. The average height of all the studied clones is 18.9±0.3 m, the average diameter of the trunk at a height of 1.3 m is 20.3±0.4 cm, the average diameter of the crown is 3.7±0.2 m. The assessment of qualitative features allowed us to identify clones with high trunk quality (straightness), with dense crowns: 6, 17, 25, 30, 22, 34, 38. In the study of clones, weak fruiting was found (from 20 to 80-100 cones for rameta), which is due to the small size of the crowns and the fruiting tier. On the basis of isoenzyme and microsatellite analysis, the genotypes of clones of plus trees were determined, as well as the frequency of occurrence of the identified alleles. As a result of the conducted studies, diagnostic allelic combinations of individual loci were identified for 73.3 % of clones. Genetic identification made it possible to establish that 98.7 % of the trees correspond to the studied clones. The discrepancy between the clonal affiliation of two trees was confirmed by both isoenzyme and microsatellite analyses.
Текст научной статьи Селекционная оценка и генотипирование клонов плюсовых деревьев Рinus sylvestris L. в Северном Казахстане
В лесной селекции с целью повышения продуктивности и качества лесов проводят отбор плюсовых деревьев по ряду ценных признаков: высота, диаметр ствола, полнодревес-ность, прямоствольность, устойчивость к вредителям и болезням и пр. Для сохранения генофонда и изучения наследственных свойств плюсовых деревьев их вегетативное потомство испытывают в архивах клонов. Проявление тех или иных качеств плюсовых деревьев, выявленных при фенотипическом отборе, во многом определяется их наследственностью (генотипом), а также влиянием условий произрастания.
Несоответствие размещения рамет клону одного генотипа на делянках может искажать итоговые данные по количественным и качественным характеристикам вегетативного потомства плюсовых деревьев. Для исключения ошибок на созданных селекционно-генетических объектах проводят генетическую паспортизацию. Так, например, определенный на основе метода аллозимного анализа и результатов фенетического этапа паспортизации уровень ошибок при маркировке клонов сосны обыкновенной в Озерском лесничестве Алтайского края составил 11 % [1].
С помощью методов молекулярно-генетического анализа, в том числе изоферментного маркирования, можно получить большое количество генетических портретов плюсовых деревьев. В Беларуси проведена работа по созданию базы данных, в которой собраны генотипы плюсовых деревьев с подробным описанием их фенотипов, используемых при закладке лесосеменных плантаций. На основе накопленной генотипической и фенотипической информации по плюсовым деревьям появилась возможность создавать лесосеменные плантации с заданными генетическими характеристиками [2].
В Казахстане на объектах селекционно-генетического значения ДНК-паспортизация до настоящего времени не проводилась, что обусловило актуальность проводимых исследований. Важное значение имеет и фенетическая характеристика, на основании которой осуществляется отбор наиболее продуктивных и урожайных генотипов.
Цель исследования – выявление клонов плюсовых деревьев сосны обыкновенной, отличающихся хозяйственно ценными признаками, а также уточнение клоновой принадлежности рамет в соответствии со схемами селекционно-генетических объектов при помощи молекулярно-генетических методов анализа.
Объекты и методы исследований
Объектами исследования являлись клоны плюсовых деревьев сосны обыкновенной Pinus sylvestris L. в возрасте 37 лет. Плюсовые деревья отобраны на территории Казахского Мелкосопоч-ника из 5 биотопов (Боровской, Воробьевский, Катаркольский, Урумкайский I, Урумкайский II). Привитые саженцы высажены на территории двух архивов со схемой размещения 3 x 3 м. Набор клонов был одинаковым.
Архивы клонов сосны обыкновенной № 1 и 2 площадью 2,7 и 3,3 га соответственно заложены В.И. Мосиным и А.И. Бреусовой в Акмолинской обл. (Северный Казахстан) в 1984 г. Преобладающим типом рельефа является мелкосопочник увалистый. Микрорельеф – слабая возвышенность с микропонижениями. Почвы темно-серые лесные. Климат территории резко континентальный. Среднегодовая температура воздуха колеблется в пределах от 1,2 до 2,0 °С. Годовое количество осадков варьирует от 250–295 мм в равнинной части до 400 мм в возвышенной.
Сохранность архивов клонов определена в соответствии с правилами проведения инвентаризации лесных культур [3].
Высота деревьев и диаметр стволов на высоте 1,3 м измерены с использованием мерной вилки Haglof (точность до 1 мм) и высотомера электронного Haglof (точность ±0,1 м). При оценке качественных признаков клонов плюсовых деревьев сосны обыкновенной были исследованы: качество ствола [4], санитарное состояние [5], плодоношение [6], устойчивость к вредителям и болезням, плотность кроны. Устойчивость к вредителям и болезням оценивалась по 5-балльной шкале: 5 баллов – органы растения не поражены; 4 – поражено 10–20 %; 3 – 21–30 %; 2 – 31–40 %; 1 балл – 41–50 % и более органов. Густоту (плотность) кроны определяли по 3-балльной шкале: 3 – густая (скелетные ветви просматриваются на 10–20 %); 2 – средней густоты (на 50 %); 1 – редкая или ажурная (видны все скелетные ветви). Молекулярно-генетический анализ и паспортизация исследуемых клонов сосны обыкновенной сделаны по 20 изофермент-ным локусам (Gpi; Fe; Pgm-1, -2; Dia-1, -2; Gdh; Adh-1, -2; 6-Pgd-1, -2; Mdh-1, -2, -3; Aat-1, -2, -3; Lap-1, -2; Idh.) и 6 микросателлитным локусам ядерной ДНК (PtTx4001, PtTx4011, Psyl2, Psyl17, Psyl36, Psyl42). Изоферментный анализ выполнен методом электрофореза в крахмальном геле [8]. Выделение ДНК при микросателлитном (SSR) анализе проведено с использованием СТАВ-метода [9]. Амплификация ДНК основывается на методе полимеразной цепной реакции.
Изменчивость изучаемых показателей оценена по шкале С.А. Мамаева [7]. Весь объем собранного материала был статистически обработан с помощью программы MS Excel 2019.
Результаты и обсуждение
В рамках исследований была проведена инвентаризация всех клонов, произрастающих в архивах. Установлено, что средняя сохранность архивов клонов № 1 и 2 на 2022 г. составляет 70,1 и 67,7 % соответственно. Общее количество рамет на объектах составляло 3 286 шт., при этом каждый клон представлен от 11 до 58 раметами (деревьями, генетически идентичными клону).
Согласно средневзвешенному баллу санитарного состояния клоны плюсовых деревьев характеризуются как ослабленные (1,7 и 1,6 баллов для архивов клонов № 1 и 2 соответственно). Доля ра-мет без признаков ослабления составляла 41,1 % общего количества на исследуемых участках.
При обследовании клонов плюсовых деревьев были обнаружены механические повреждения стволов, среди которых присутствовали обдиры различной площади. Обдиры коры доходили до камбия, в результате чего на стволах клонов выявлены следы смолотечения. Можно предположить, что повреждения регулярно наносят дикие животные (косули), для которых тонкая кора в нижней части дерева является источником дополнительных питательных веществ. Доля деревьев с такими повреждениями в архивах клонов № 1 и 2 составила 56,8 и 44,7 % соответственно. Большее количество деревьев с сильными механическими повреждениями (высотой до 2 м и шириной от 1/2 диаметра ствола и более) отмечено в архиве клона № 1, где на их долю приходилось 20,9 %. В архиве клона № 2 деревьев с такой же степенью повреждений в 2 раза меньше (10,3 %). Увеличение площади повреждений по всему диаметру ствола и полное повреждение луба (флоэмы) может привести к нарушению движения нисходящего потока растворов органических веществ, снижению устойчивости и жизнеспособности растений и затем к их гибели [10].
Первоначально селекционно-генетическая оценка и молекулярно-генетическая паспортизация были проведены для 15 клонов плюсовых деревьев сосны обыкновенной. На рис. 1 представлены биометрические показатели и сохранность изучаемых клонов.
Размеры ствола (высота и диметр) основных лесообразующих пород являются важнейшими показателями продуктивности. Исследованиям по отбору и изучению роста потомства плюсовых деревьев посвящено много работ, в которых отображены основные результаты по выделению наиболее быстрорастущих и продуктивных генотипов. Провести сравнительную характеристику высоты и диаметра потомств плюсовых деревьев в различных частях естественного ареала сосны обыкновенной в определенном возрасте довольно трудно. Однако из ранее обобщенных материалов С.А. Мамаева известно, что значительная разница показателей высоты и диаметра обусловлена географической изменчивостью, при которой наблюдается трансформация совокупности экологических факторов, влияющих на таксационные показатели [11].
Высота исследуемых клонов варьировала от 16,4±0,3 м (CV 9,28 %)уклона № 27до 20,7±0,3 м (CV 6,03 %) у клона № 22. Наибольшая средняя
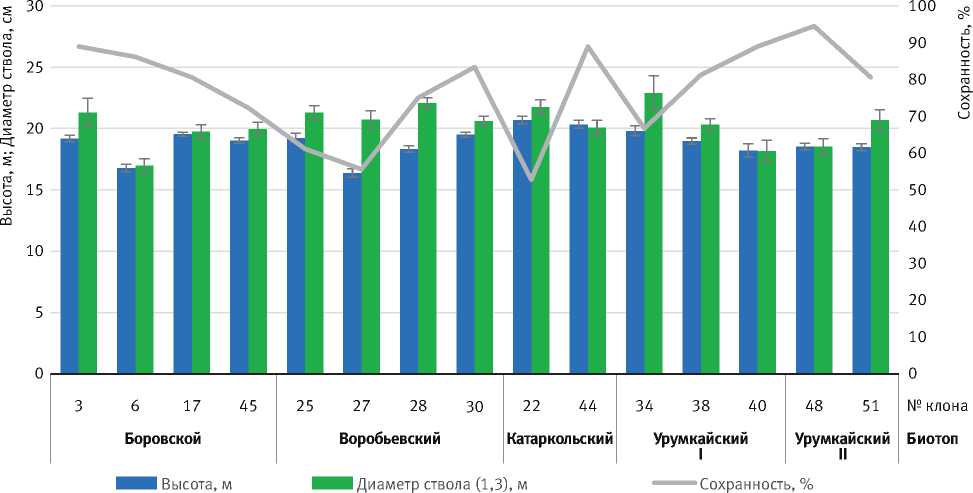
Рис. 1. Средние биометрические показатели клонов плюсовых деревьев и их сохранность в 37 лет
высота выявлена у клонов Катаркольского биотопа – 20,5±0,2 м. Другие клоны в зависимости от происхождения в среднем достигали высоты от 18,4±0,3 до 19,0±0,2 м. Клоны Воробьевского биотопа отличались низкорослостью (среднее по биотопу – 18,4±0,2 м), но при этом имели самые большие значения диаметра ствола (среднее по биотопу – 21,2±0,1 см). Наиболее низкие показатели высоты и диаметра зафиксированы у клона № 6 – 16,8±0,3 м (CV 7,65 %) и 17,0±0,6 см (CV 13,64 %) соответственно.
По результатам наблюдений высота деревьев клонового потомства сосны обыкновенной во многом обусловлена их генотипом, а также воздействием абиотических факторов.
Сохранность исследуемых клонов в среднем составила 77,1 %, при этом наиболее высокая – у клона № 48 (94,4 %). Немногим более половины рамет сохранилось у клона № 22 (52,8 %). Самая низкая средняя сохранность – у Воробьевского биотопа (68,8 %). Около 30 % рамет у клонов этого биотопа выпали в первой половине испытания, как и у клонов Катаркольского и Урумкай-ского I биотопов. Незначительная гибель рамет в первой половине жизни наблюдалась у клонов Урумкайского II и Боровского биотопов (9,7 и 13,9 % соответственно).
Состояние клонов было оценено визуально, на основе признаков, характеризующих ствол, крону и устойчивость к неблагоприятным факторам (табл. 1).
Слабое искривление ствола (прогиб менее 10 см на 2 м длины ствола) встречалось у 46,7 % клонов всех биотопов. Достаточно большой наклон ствола (75–80°) и сильное его искривление зафиксировано у клона № 40. Высокое качество ствола отмечено у клонов № 6, 17, 25, 30, 22, 34, 38, раметы которых отличались прямоствольностью.
Значительная доля клонов (60,4 %) отнесена к категории ослабленных. На состояние клонов могло повлиять механическое повреждение стволов (объедание коры животными). В кроне некоторых рамет происходило усыхание отдельных ветвей и пожелтение хвои. Массовых повреждений вредителями и болезнями не обнаружено, за исключением незначительного поражения ассимиляционного аппарата. Наибольшая устойчивость к вредителям и болезням выявлена у клонов № 6 и 25, признаков ослабления у которых не наблюдалось.
Крона большинства клонов (80,3 %) средней густоты или редкая. Только у клонов № 27, 28, 30 Воробьевского биотопа сформировалась густая
Таблица 1. Оценка качественных признаков клонов плюсовых деревьев сосны обыкновенной
|
№ |
Качество ствола, балл |
Санитарное состояние, балл |
Устойчивость к вредителям и болезням, балл |
Плотность кроны, балл |
Плодоношение, балл |
|
клона |
Х±mx |
Х±mx |
Х±mx |
Х±mx |
Х±mx |
|
Боровской биотоп |
|||||
|
3 |
4,39±0,16 |
1,94±0,13 |
3,78±0,10 |
2,44±0,17 |
0,26±0,03 |
|
6 |
4,71±0,14 |
1,41±0,15 |
4,59±0,15 |
1,41±0,12 |
0,18±0,03 |
|
17 |
4,82±0,13 |
1,41±0,12 |
4,29±0,11 |
1,41±0,12 |
0,19±0,02 |
|
45 |
4,46±0,18 |
1,77±0,12 |
4,23±0,12 |
1,38±0,14 |
0,13±0,02 |
|
Воробьевский биотоп |
|||||
|
25 |
5,00±0,00 |
1,13±0,09 |
4,87±0,09 |
1,80±0,11 |
0,25±0,05 |
|
27 |
4,40±0,17 |
1,70±0,11 |
4,00±0,00 |
2,60±0,11 |
0,19±0,04 |
|
28 |
4,24±0,16 |
1,82±0,10 |
3,47±0,12 |
2,59±0,12 |
0,45±0,04 |
|
30 |
5,00±0,00 |
1,06±0,06 |
4,50±0,12 |
2,56±0,12 |
0,42±0,06 |
|
Катаркольский биотоп |
|||||
|
22 |
4,59±0,17 |
1,76±0,11 |
4,18±0,10 |
2,24±0,14 |
0,23±0,02 |
|
44 |
4,29±0,14 |
1,59±0,12 |
4,41±0,12 |
1,53±0,12 |
0,26±0,04 |
|
Урумкайский I биотоп |
|||||
|
34 |
4,55±0,21 |
1,63±0,15 |
4,36±0,15 |
1,73±0,14 |
0,24±0,04 |
|
38 |
4,65±0,12 |
1,26±0,09 |
4,13±0,13 |
2,39±0,15 |
0,49±0,05 |
|
40 |
3,53±0,13 |
2,20±0,17 |
3,80±0,17 |
1,07±0,07 |
0,26±0,06 |
|
Урумкайский II биотоп |
|||||
|
48 |
4,12±0,17 |
2,18±0,13 |
3,82±0,13 |
1,24±0,11 |
0,24±0,05 |
|
51 |
4,50±0,16 |
1,25±0,11 |
4,13±0,09 |
2,38±0,15 |
0,25±0,03 |
крона с хорошим охвоением. Плодоношение клонов слабое. В среднем в кроне формируется от 20 до 100 шишек (клоны № 28, 30, 38). Наличие определенного количества шишек у клонов в настоящее время обусловлено в большей степени сформированностью кроны. В возрасте 37 лет при изначальном размещении рамет 3 x 3 м крона клонов находится на вершине ствола. Для наиболее полной характеристики был измерен диаметр кроны, а также рассчитаны доля ее протяженности от высоты ствола и протяженности мужского яруса (табл. 2).
Диаметр кроны клонов варьировал от 2,85±0,41 м (клон № 45) до 4,82±0,26 м (клон № 27). У большей доли клонов (40,0 %) этот показатель имел повышенный уровень изменчивости. Очень высокая изменчивость диаметра кроны отмечена у клонов № 6, 40 и 51, при том что их сохранность варьировала в пределах
80,6–88,9 %. Наиболее широкая крона сформировалась у клонов Воробьевского и Катаркольского биотопов – в среднем 4,06±0,27 и 4,37±0,19 м соответственно. Средний диаметр кроны клонов других биотопов был меньше на 21,2–26,8 %. Протяженность кроны всех клонов в среднем составляет 9,01±0,24 м, или 47,7 % протяженности ствола. При этом шишки формировались только в верхней освещенной части кроны в женском ярусе, доля протяженности которого в среднем составляла 28,7 % общей высоты ствола. Наиболее короткая крона у клонов Урумкайского I биотопа – 7,98±0,46 м. Показатель протяженности кроны всех клонов имеет широкий диапазон меж- и внутриклоновых значений, о чем свидетельствует разброс уровней изменчивости признака от среднего до очень высокого. Более половины кроны клонов (61,1 %) занимал плодоносящий ярус, в котором наблюдалось небольшое
Таблица 2. Характеристика размеров крон исследуемых клонов плюсовых деревьев сосны обыкновенной
|
Клон |
Диаметр кроны, м |
Протяженность кроны, м |
Протяженность мужского яруса, м |
|||
|
Х±mx |
CV, % |
Х±mx |
CV, % |
Х±mx |
CV, % |
|
|
Боровской биотоп |
||||||
|
3 |
4,04±0,26 |
27,43 |
10,81±0,93 |
36,52 |
5,28±0,87 |
69,86 |
|
6 |
3,11±0,24 |
32,58 |
8,09±0,54 |
28,25 |
2,88±0,29 |
40,93 |
|
17 |
3,35±0,23 |
29,67 |
8,51±0,60 |
30,02 |
3,26±0,32 |
39,83 |
|
45 |
2,85±0,41 |
58,91 |
5,73±0,82 |
58,97 |
2,27±0,31 |
48,95 |
|
Воробьевский биотоп |
||||||
|
25 |
3,04±0,42 |
61,97 |
6,82±0,93 |
60,93 |
2,52±0,31 |
48,00 |
|
27 |
4,82±0,26 |
23,81 |
8,95±0,56 |
28,16 |
5,30±0,51 |
42,89 |
|
28 |
4,42±0,29 |
28,30 |
9,76±0,82 |
35,69 |
5,12±0,32 |
25,74 |
|
30 |
3,94±0,12 |
12,39 |
10,07±0,33 |
13,88 |
3,89±0,38 |
41,26 |
|
Катаркольский биотоп |
||||||
|
22 |
4,56±0,17 |
15,33 |
14,12±0,58 |
16,93 |
6,18±0,51 |
33,98 |
|
44 |
4,17±0,21 |
21,05 |
10,41±0,64 |
25,49 |
4,53±0,46 |
42,12 |
|
Урумкайский I биотоп |
||||||
|
34 |
2,93±0,50 |
65,53 |
7,13±1,23 |
67,00 |
1,56±0,21 |
43,87 |
|
38 |
3,27±0,13 |
18,91 |
9,46±0,35 |
17,59 |
3,70±0,22 |
28,77 |
|
40 |
3,59±0,33 |
36,46 |
7,36±0,65 |
35,11 |
1,93±0,19 |
37,38 |
|
Урумкайский II биотоп |
||||||
|
48 |
4,06±0,21 |
21,46 |
9,66±0,51 |
21,72 |
2,76±0,22 |
33,30 |
|
51 |
3,59±0,30 |
34,30 |
8,29±0,78 |
38,64 |
2,56±0,33 |
50,82 |
количество шишек на периферии верхней части кроны (средний балл плодоношения 0,28±0,06).
Таким образом, исследуемые клоны можно охарактеризовать как деревья среднепирамидальной формы кроны средней плотности со смешанным половым типом, где плодоносящий ярус занимает большую половину верхней части кроны.
Исследования роста клонов плюсовых деревьев сосны обыкновенной и показателей их урожайности направлены на выявление наиболее перспективных генотипов для создания лесосеменных плантаций. Данные исследования были проведены рядом ученых в субъектах Российской Федерации [12–15]. Так, известны результаты исследований, которые показывают, что в самых крупных шишках содержатся семена большей массы [16]. Масса семян способна оказывать сильное влияние на рост однолетних сеянцев в высоту, затем это воздействие уменьшается и исчезает к 5–7 годам [17].
При проведении изоферментного анализа рамет плюсовых деревьев сосны обыкновенной было выявлено 49 аллельных вариантов. Преобладающим являлся аллель 1.00, за исключением локуса 6-Pgd-2, у которого частота аллельного варианта 0.90 выше частоты основного аллеля для сосны обыкновенной. Из 20 проанализированных локусов 18 оказались полиморфными.
На основе молекулярно-генетического анализа 15 клонов сосны обыкновенной и их паспортизации составлены генотипы по 20 изо-ферментным локусам. Для большинства клонов выявлены диагностические аллельные сочетания отдельных локусов, характерные только для представителей данного клона. Для клонов № 28, 30, 34 и 44 такие специфические комбинации аллелей отдельных локусов обнаружены не были.
Среди проанализированных клонов наибольшее количество изоферментных локусов, находящихся в гетерозиготном состоянии, установлено у клонов № 48 и 51 (8 шт.), наименьшее – у клонов № 40 (4 шт.), 28 и 34 (по 5 шт.).
Необходимо отметить, что только 4 проанализированных генотипа (5,3 %) не соответствуют своей предполагаемой клоновой принадлежности (по одной рамете из клонов № 25, 27, 28, 38). Из них один генотип показал свою идентичность другому клону сосны обыкновенной. Так, рамета, отнесенная к клону № 25, по сочетанию аллельных вариантов 20 изоферментных генов была сходна с раметой клона № 38. В случае раметы клона № 45 для изоферментного анализа была представлена смесь семян от разных индивидуумов, о чем свидетельствуют генотипы по локусам 6-Pgd-1 (0.85/0.95/1.00) и Aat-2 (1.00/1.10/1.25). Генотип этой раметы был установлен при проведении ДНК-анализа.
Микросателлитный анализ клонов плюсовых деревьев сосны обыкновенной позволил проверить и уточнить полученные результаты для 4-х рамет, не соответствующих своей клоновой принадлежности.
При проведении микросателлитного анализа рамет плюсовых деревьев сосны обыкновенной выявлено 25 аллельных вариантов. По каждому локусу выявлен 1 преобладающий аллель, а для локуса Psyl42 – 2 (175 и 177 п.о.).
На основании выявленных ПЦР-фенотипов и полиморфизма аллельных вариантов были составлены соответствующие им генотипы клонов сосны обыкновенной (рис. 2).
Проведение SSR-анализа показало, что раметы клонов № 28, 38 и 45 (представленные в изоферментном анализе смесью семян разных деревьев) идентичны заявляемой клоновой принадлежности. Различия, выявленные при проведении изоферментного анализа, могли быть связаны со сложностью определения диплоидных генотипов взрослых деревьев по смеси различных гаплоидных (семена), в том числе с наличием примесей семян от других деревьев.
Для двух деревьев сосны обыкновенной, как и в случае изоферментного анализа, выявлено несоответствие их предполагаемой клоновой принадлежности. Было подтверждено, что рамета, отнесенная к клону № 25, по сочетанию аллельных вариантов 6-ти микросателлитных локусов
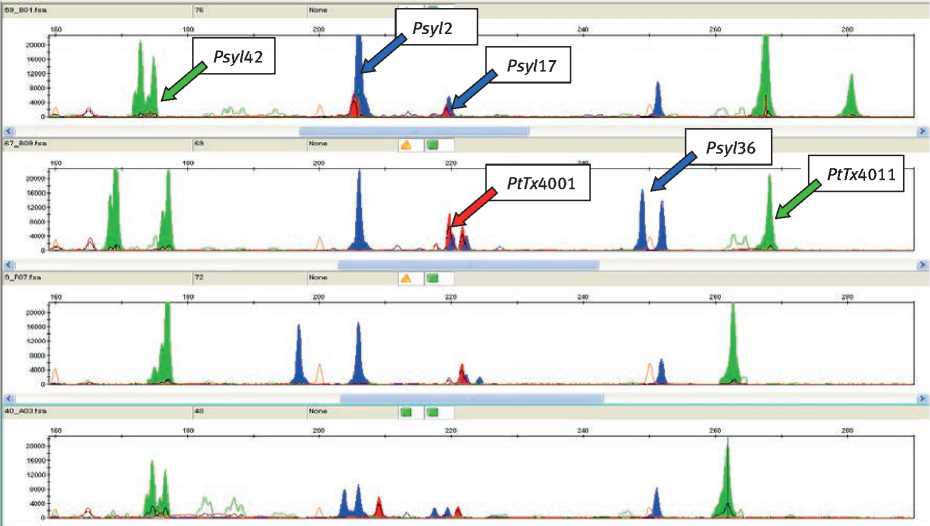
Рис. 2. Генотипы клонов сосны обыкновенной по SSR-локусам ядерной ДНК
(SRR-спектры: 1 – клон № 25; 2 – клон № 30; 3 – клон № 48; 4 – клон № 51)
ядерной ДНК, как и изоферментных, идентична раметам клона № 38. При проведении обоих видов молекулярно-генетического анализа из проанализированных рамет клонов плюсовых деревьев сосны обыкновенной совпадений не было выявлено только для одной раметы клона № 27.
Паспортизация и идентификация 15 клонов сосны обыкновенной на основе сформированного набора молекулярно-генетических маркеров (20 изоферментных и 6 микросателлитных) характеризуются высокой достоверностью дифференциации генотипов (вероятностью ошибки выявления сходных генотипов).
Выводы
В результате проведенных исследований получены данные об общем состоянии и таксационной характеристике клонов плюсовых деревьев сосны обыкновенной в возрасте 37 лет в Северном Казахстане. Установлено, что раме-ты клонов Воробьевского биотопа отличались самой низкой средней сохранностью (68,8 %) и наименьшей высотой ствола (среднее по биотопу – 18,4±0,2 м), наибольшими значениями диаметра ствола (среднее по биотопу – 21,2±0,1 см) и высоким средним баллом густоты кроны (2,4 балла). Самые высокие клоны отмечены в Катаркольском биотопе (среднее по биотопу – 20,5±0,2 м), максимальная высота – у клона № 22 (20,69±0,30 м). Высокое качество ствола выявлено у 46,7 % клонов. Проведенная оценка качественных признаков позволила выявить клоны с высоким качеством ствола (прямо-ствольность), густым охвоением и наибольшей устойчивостью: № 6, 17, 25, 30, 22, 34, 38. По результатам молекулярно-генетического анализа соответствие клоновой принадлежности согласно схемам селекционно-генетических объектов выявлено у 98,7 % проанализированной выборки рамет.
Данное исследование финансируется Министерством экологии, геологии и природных ресурсов Республики Казахстан (ИРН BR10263776) This study is funded by the Ministry of Ecology, Geology and Natural Resources of the Republic of Kazakhstan (№BR10263776)
Список литературы Селекционная оценка и генотипирование клонов плюсовых деревьев Рinus sylvestris L. в Северном Казахстане
- Зацепина, К.Г. Генотипирование деревьев на клоновых плантациях хвойных лесообразующих видов в Западной Сибири / К.Г. Зацепина, А.К. Экарт, В.В. Тараканов // Хвойные бореальной зоны. - 2012. - Т. XXX. - № 1-2. - С. 67-71.
- Ковалевич, А.И. Моделирование лесосеменных плантаций с заданной популяционно-генетической структурой семенного потомства / А.И. Ковалевич, А.П. Кончиц, А.И. Сидор // Труды БГТУ. - Серия I, Лесное хоз-во. - Минск: УО БГТУ, 2008. - Вып. XVI - С. 412-416.
- Об утверждении Правил проведения инвентаризации лесных культур, питомников, площадей с проведенными мерами содействия естественному возобновлению леса и оставленных под естественное заращивание в государственном лесном фонде. - Текст: электронный. - Приказ Министерства сельского хозяйства Республики Казахстан от 19.10.2012 № 17-02/532. - Режим доступа: URL: https://adilet.zan.kz/rus/docs/V1200008095/ (дата обращения: 27.09.2022/
- Проказин, Е.П. Изучение имеющихся и создание новых географических культур (программа и методика работ) / Е.П. Проказин. - Пушкино, 1972. - 52 с.
- Об утверждении Правил рубок леса на участках государственного лесного фонда. - Тескст: электронный. - Приказ Министерства сельского хозяйства Республики Казахстан от 30.06.2015 № 18-02/596. - Режим доступа: URL: https://adilet.zan.kz/rus/docs/V1500011894/(дата обращения: 20.12.2022).
- Бреусова, А.И. Прогноз и учёт урожая шишек и семян сосны обыкновенной на ПЛСУ и ЛСП / А.И. Бреусо-ва // Информационный листок № 05. - 2003. - 4 с.
- Мамаев, С.А. Основные принципы методики исследования внутривидовой изменчивости древесных растений / С.А. Мамаев // Индивидуальная эколого-географическая изменчивость растений. - Вып. 94. - Свердловск: УНЦ АН СССР, 1975. - С. 3-14.
- Гончаренко, Г.Г. Руководство по исследованию хвойных видов методом электрофоретического анализа изоферментов / Г.Г. Гончаренко, В.Е. Падутов, В.В. Потенко. - Гомель, 1989. - 164 с.
- Падутов, В.Е. Методы молекулярно-генетического анализа / В.Е. Падутов, О.Ю. Баранов, Е.В. Воропаев. -Минск: Юнипол, 2007. - 176 с.
- Сапанов, М.К. Причины гибели сосны обыкновенной в старовозрастных культурах Нарынских песков / М.К. Сапанов, М.Л. Сиземская // Лесоведение. - 2018. - № 5. - С. 381-388. - DOI 10.1134/S0024114818050078.
- Мамаев, С.А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале) / С.А. Мамаев. - Москва: Наука, 1972. - 284 с.
- Шейкина, О.В. Фенотипическая и генетическая изменчивость клонов плюсовых деревьев сосны обыкновенной на лесосеменной плантации в Чувашской Республике / О.В. Шейкина, Э.П. Лебедева, З.Х. Шигапов. -Йошкар-Ола: Поволжский государственный технологический университет, 2013. - 160 с.
- Коновалов, В.Ф. Состояние и закономерности роста сосны обыкновенной на лесосеменных объектах Республики Башкортостан / В.Ф. Коновалов, Э.Р. Насырова // Лесной вестник. - 2016. - Т. 20. - № 2. -С. 60-65.
- Багаев, С.С. Результаты проверки наследственности плюсовых деревьев сосны обыкновенной и лиственницы Сукачева на быстроту роста и репродуктивную способность в Волжском спецсемлесхозе / С.С. Багаев // Плодоводство, семеноводство, интродукция древесных растений. - 2017. - Т. 20. - С. 15-17.
- Бессчетнова, Н.Н. Таксационные показатели вегетативного потомства плюсовых деревьев сосны обыкновенной (Pinus sylvestris L.) в архивах клонов в Нижегородской области / Н.Н. Бессчетнова, В.П. Бессчетнов, Т.А. Оганян // Экономические аспекты развития АПК и лесного хозяйства. Лесное хозяйство Союзного государства России и Белоруссии: матер. междунар. науч.-практ. конф. - Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2019. - С. 115-122.
- Aniszewska, M. Connection between shape of pine (Pinus sylvestris) cones and weight, color and number of seeds extracted from them / M. Aniszewska // Electronic Journal of Polish Agricultural Universities. - 2006. -9 (1). - Режим доступа: URL: http:.www.ejpau.media.pl/articles/volume9/issue1/art-03.pdf / (дата обращения: 23.12.2022).
- Reich, P.B. Seed mass effects on germination and growth of diverse European Scots pine populations / P.B. Reich, J. Oleksyn, M.G. Tjoelker // Canadian Journal of Forest Research. - 1994. - 24(2). - Р. 306-320. - Doi:10.1139/ x94-044.
