Сезонная активность дождевых червей (Oligochaeta, Lumbriсidae) пояса широколиственных лесов Кабардино-Балкарского государственного высокогорного заповедника и прилегающих территорий (Центральный Кавказ)
 пояса широколиственных лесов Кабардино-Балкарского государственного высокогорного заповедника и прилегающих территорий (Центральный Кавказ)")
Автор: Рапопорт И.Б.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: фауна
Статья в выпуске: 1-5 т.12, 2010 года.
Бесплатный доступ
Исследованы видовое разнообразие, биотопическое распределение и сезонная динамика показателей обилия дождевых червей в среднегорных биотопах с различным гидротермическим режимом. Впервые на Центральном Кавказе найден Aporrectodea handlirschi (Rosa, 1897) - редкий исчезающий вид, занесенный в Красную книгу Российской Федерации. Впервые для условий Северного Кавказа у A. rosea (Savigny, 1826) зафиксирован переход летней диапаузы в зимнюю.
Дождевые черви, фауна, экология, сезонная динамика
Короткий адрес: https://sciup.org/148199159
IDR: 148199159 | УДК: 591.9:595.14
Seasonal activity of earthworms (Oligochaeta, Lumbricidae) from the belt of broad-leaved forests in Kabardino-Balkarian state high-mountain reserve and adjacent territories (Central Caucasus)
Species diversity, biotopic distribution and seasonal dynamics of abundance indexes in earthworms from middle mountains biotopes of various hydrothermal regimen are studied. Aporrectodea handlirschi (Rosa, 1897) has been first identified in the Central Caucasus. The change from summer diapause in A. rosea (Savigny, 1826) in first re-corded in the North Caucasus.
Текст научной статьи Сезонная активность дождевых червей (Oligochaeta, Lumbriсidae) пояса широколиственных лесов Кабардино-Балкарского государственного высокогорного заповедника и прилегающих территорий (Центральный Кавказ)
Широко известна целесообразность использования автохтонных видов дождевых червей в восстановлении почвенного плодородия, что не возможно без изучения фауны, экологии и ландшафтного распределения видов. Сезонная динамика численности отдельных видов люмбрицид исследована в Закавказье [1], на северном макросклоне Большого Кавказа работы не проводились. Целью работы являлось изучение видового разнообразия, биотопического распределения и сезонной динамики показателей обилия дождевых червей в среднегорных биотопах с различным гидротермическим режимом.
Краткое описание района исследований. Кабардино-Балкарский высокогорный заповедник (КБГВЗ) создан для сохранения и изучения природных комплексов высокогорья северного макросклона Центрального Кавказа. Пояс широколиственных лесов в пределах заповедника представлен на небольшой территории – в бассейне р. Черек Балкарский и его притоков. Ущелье р. Черек Балкарский южнее Нижнего Голубого озера образует закрытую со всех сторон высокими хребтами Верхне-Балкарскую котловину, в которую не проникают влажные воздушные течения. Климат ее отличается сухостью, склоны безлесны, покрыты в нижней части нагорно-ксерофитной растительностью. Выше по ущелью происходит переход на-горно-ксерофит-ной растительности в кустарниковые заросли орешника с рододендроном желтым и далее в лиственный лес. Среднегодовое количество осадков в среднем по лесному поясу составляет 600-750 мм, в районе Верхне-Балкарской аридной котловины – не более 400 мм. Почвы района преимущественно маломощные, с небольшим гумусовым
слоем и камененисто-щебнистым почвенным профилем.
Материал и методы. Сбор материала проводился на правом берегу р. Черек Балкарский с 2006 по 2007 гг. Исключение составил февраль, когда дороги были трудно проходимы из-за обильно выпавшего снега. Беспозвоночные отобраны из почвенных монолитов (25х25 см) в соответствии с общепринятой методикой [3]. Идентификация дождевых червей проведена по работе Т.С. Всеволодовой-Перель [4]. При выборе учетных площадок использовали метод линейной трансекты [5]. Мы исходили из того, что тип растительного сообщества является в определенной степени показателем гидротермических условий биотопа, поэтому выбирались такие элементы ландшафта, градиент температуры и влажности которых отражал бы крайние значения для исследуемых территорий. Все площадки закладывались на северо-западном макросклоне ущелья, на платообразных участках и педиментах рельефа, уклон которых не превышает 3-5°. Исследованы: нагор-но-ксерофитная растительность аридной котловины (1068 м над ур. м., 5 км ниже с. Верхняя Балкария), древесно-кустарниковые заросли лещины с рододендроном желтым (буферная зона КБГВЗ, 1160 м над ур. м., 2 км выше с. Верхняя Балкария), мезофитные и гигрофитные лесные сообщества ольшаника (КБГВЗ, 1320 м над ур. м., 6 км выше с. Верхняя Балкария).
Результаты и их обсуждение. В общей сложности на исследованной территории выявлено 9 видов дождевых червей: Aporrectodea handlirschi (Rosa, 1897), A. rosea (Savigny, 1826), Dendrobaena octaedra (Savigny, 1826), D. schmidti Мichaelsen, 1907, D. tellermanica Perel, 1966, Dendrodrilus ru-bidus tenuis (Eisen, 1874), Eisenia fetida (Savigny, 1826), Eiseniella tetraedra (Savigny, 1826), Lumbri-cus rubellus Hoffmeister, 1843.
A. handlirschi встречен в одной пробе – в ольшанике, в прирусловой части притока Черека Балкарского. На Центральном Кавказе отмечен впервые. Реликтовый, исчезающий почвенноподстилочный вид, занесенный в Красную книгу Российской Федерации [6]. Распространен в предгорьях и низкогорьях Северо-Западного Кавказа, за пределами России обитает в Альпах, Северных Карпатах и севере Балкан, а также на северо-западе Малой Азии [4]. Наиболее часто встречается в пробах D. schmidti – крымско-кавказский эндемик, мезофилл, широко распространенный от степной зоны до альпийского пояса. На Центральном Кавказе вид представлен тремя морфоэкологическими формами [7]. Автохтонным является D. tellermanica – собственно почвенный вид, хорошо приспособленный к обитанию в засушливых условиях. За пределами Кавказа pзарегистрирован в нагорных дубравах правобережья Хопра, на Калачской возвышенности, юговосточных склонах Среднерусской возвышенности, Алтае, Южном Урале [4]. Остальные виды относятся к числу космополитов. Обитание собственно почвенного A. rosea на Центральном Кавказе приурочено к аридным и лесостепным ландшафтам. Почвенно-подстилочный E. fetida тяготеет к местам скопления органики, предпочитает влажные почвы и околоводные биотопы. Мезофильный L. rubellus в пределах Центрального Кавказа редок, в основном сопутствует жилью человека. Подстилочные D. octaedra и D. rubidus tenuis характерны для пояса широколиственных лесов и субальпики, амфибионт E. tetraedra чаще встречается в среднегорных ландшафтах под лесной растительностью в почвах с проточным увлажнением.
Участок с нагорно-ксерофитной растительностью (Верхне-Балкарская аридная котловина). Почти вся поступающая влага расходуется на сток и испарение и даже в апреле после таяния снега верхний слой почвы представляет сухую спрессованную корку. В сообществах Верхне-Балкарской котловины зарегистрированы A. rosea и D. teller-manica – виды, хорошо приспособленные к жестким ксеротермическим условиям (рис. 1).
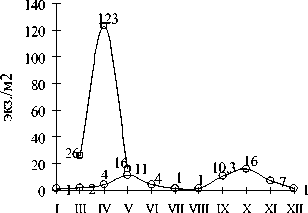
A. rosea
D. tellermanica месяц
Рис. 1. Сезонная динамика численности дождевых червей в биотопе с нагорно-ксерофитной растительностью
-
A. rosea в активном состоянии найден на глубине 15-30 см в конце марта, апреле (максимальная численность 123±19 экз./м2) и мае. Т.к.
распределение его в апреле – мае не являлось агрегированным, и он отмечался во всех пробах, мы считаем, что A. rosea доминировал в биотопе. Вероятно в данных условиях на уровне 15-35 см протекает репродуктивный период и развитие ювенильных особей. Переживание зимних месяцев и летняя диапауза происходят на большей глубине – 35-50 см. В апреле в пробах преобладали половозрелые особи, в мае были собраны в основном молодые черви. Осень 2007 г. была очень сухой, возможно, поэтому у A. rosea произошел переход летней диапаузы в зимнюю. Подобный факт ранее фиксировался в предгорно-степных ландшафтах Грузии и пустынно-степных светлоземах Средней Азии [1, 12].
-
D. tellermanica является экологическим викариатом A. rosea . За время наблюдений выявлено 2 периода активной жизнедеятельности – апрель-июнь и сентябрь-октябрь, с максимумом в октябре (16±8 экз./м2). В марте в пробах преобладали половозрелые особи и субадульты, в апреле, мае и октябре преобладали молодые черви. Диапази-рующие черви с июня по сентябрь отмечались в слое 25-35 см.
Древесно-кустарниковые заросли лещины с рододендроном желтым. Биотоп отличается жестким гидротермическим режимом и каменистощебнистым почвенным профилем. Найдены 2 вида дождевых червей – D. schmidti и D. octaedra , первый из которых доминировал на исследованной территории, второй встречался редко (рис. 2). Единичные особи D. octaedra отмечены в апреле– мае и в августе–октябре исключительно на затененных участках (под куртинами рододендрона и березы). В июне – июле спорадические находки вида приурочены к камням и корням деревьев. Фактором, ограничивающим распространение, вероятнее всего, служит постоянная инсоляция биотопа. Максимальная численность D. octaedra отмечена в сентябре – 10,7±3,3 экз./м2. Зимой в подстилке было собрано несколько коконов. D. schmidti встречался на микростациях рельефа с более развитым гумусовым слоем, наличием кротовин и древесным затенением. В конце марта едва вышедшие из диапаузы черви были собраны в пороях кабана. Период активной жизнедеятельности – с апреля по октябрь месяц. Пики численности наблюдались в мае (37±12 экз./м2) и сентябре (36±9 экз./м2). Летом D. schmidti концентрировался под камнями и у корней деревьев, поэтому в количественных пробах встречался редко.
Мезофитные и гигрофитные сообщества лиственного леса с преобладанием ольхи серой на берегу притока р. Черек Балкарский . Биотоп, в котором сезонные и суточные вариациями температуры и влажности наиболее сглажены. В мезофит-ных сообществах зарегистрированы: D. schmidti, D. octaedra , D. rubidus tenuis (рис. 3) . Доминировал D. schmidti , встречаясь с мая по октябрь на глубине 5-15 см. Минимум численности отмечен в мае (16,0±3,3 экз.м2), максимум в августе (34,7±4,5 экз.м2).
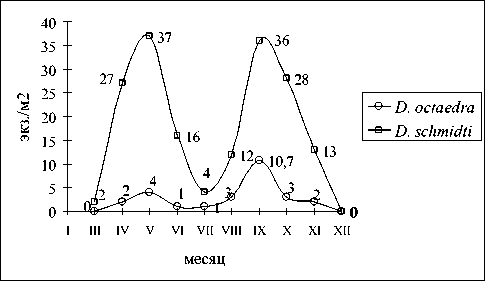
Рис. 2. Сезонная динамика численности дождевых червей в биотопе лещина-рододендрон желтый
D. octaedra – субдоминант исследованной территории. Отмечен в подстилке, верхнем (0-5 см) слое почвы, под корой разрушающейся древесины. Наибольшие показатели численности наблюдались в июне, августе, сентябре (около 16 экз./м2), наименьшие – в апреле. В ноябре – марте различные возрастные стадии вида зарегистрированы вблизи корней деревьев, под валежником. В подстилке под снегом в течение зимы удалось собрать только коконы D. octaedra . D. rubidus tenuis встречается с мая по ноябрь (до 16±6,5 экз./м2 в августе-сентябре). В апреле единичные особи были найдены у корней деревьев и в оттаявшей почве под проекцией ветвей валежника. Зимующих червей этого вида в мезофитных биотопах зарегистрировать не удалось.
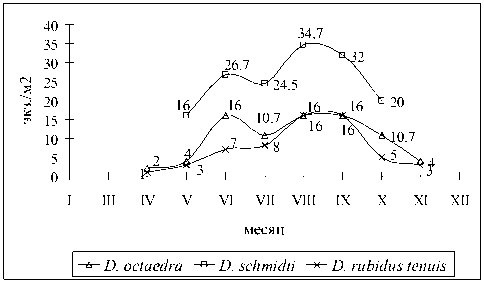
Рис. 3. Сезонная динамика численности дождевых червей в мезофитных сообществах ольшаника
В гигрофитных лесных сообществах (почва благодаря быстрому течению ручья зимой не промерзает) видовое богатство выше (рис. 4). Зарегистрированы: D. schmidti , D. octaedra , D. rubidus tenuis , E. fetida , L. rubellus , E. tetraedra tetraedra и A. handlirschi . Доминант D. schmidti фиксировался в течение всего года в верхнем слое почвы (0-10 см) с максимумом численности в августе (104±34,15 экз./м2). Скопления неполовозрелых червей отмечены в апреле (до 9 экз. на пробу 25х25 см2 при средней численности 17,3±6,15 экз./м2), июне, августе и ноябре.
В значительной степени колебания численности D. schmidti , D. octaedra и D. rubidus tenuis определялись концентрацией дождевых червей на прибрежных участках, как в летний период, так и зимой. Максимальное обилие D. octaedra – 28,8±8,1 экз./м2 – зарегистрировано в июне. В ноябре – марте различные возрастные стадии вида собраны в незамерзающей почве ручья. D. rubidus tenuis в гигрофитных сообществах чаще отмечался в мае – сентябре с максимумом (20±4,6 экз./м2) в августе. Зимой единичные особи D. rubidus tenuis отмечены в хорошо инсолируемых околоводных биотопах у корней деревьев. В зимних сборах преобладали коконы и половозрелые черви. E. fetida в течение всего года фиксировался в прибрежной почве. Встречался в русле ручья (в том числе ювенильные особи) и небольших островках по его центру. Наибольшая численность E. fetidа зарегистрирована в мае (40,0±16,0 экз./м2). L. rubellus отмечен с мая по сентябрь, чаще всего в мае и августе (24±9,2 экз./м2). Данные по численности A. handlirschi и E. tetraedra не приводятся. 2 особи A. handlirschi отмечены в июле в одной пробе. Агрегация E. tetraedra наблюдалась в декабре (6 особей в пробе 25х25 см2). В январе вид встречался спорадически, в остальное время года был приурочен к гидрофитным участкам биотопа.
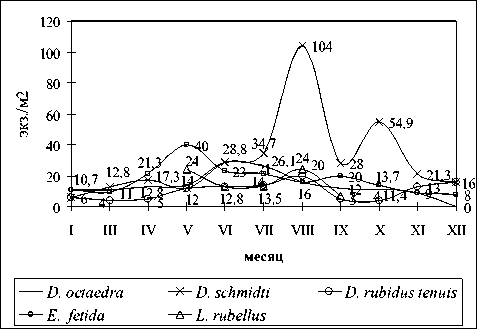
Рис. 4. Сезонная динамика численности дождевых червей в гигрофитных сообществах ольшаника
Таким образом, для большинства исследованных сообществ, за исключением гигрофитного биотопа, характерна зимняя диапауза продолжительностью от 4 (нагорно-ксерофитная растительность) до 5 месяцев (мезофитные лесные сообщества). Наличие летнего периода покоя определяется влажностью биотопов и может варьировать от полного отсутствия (лесные сообщества) до 4 месяцев (нагорно-ксерофитная растительность).
Выводы:
-
1. На исследованной территории найдено 9 видов дождевых червей. Наибольшее видовое разнообразие наблюдается в биотопе с наилучшим режимом влажности – ольшанике на берегу ручья (7 видов). В сообществах с нагорно-ксерофитной растительностью обитают D. tellermanica и A.
-
2. В ущелье р. Черек Балкарский впервые на Центральном Кавказе обнаружен A. handlirschi – реликтовый, исчезающий вид, занесенный в Красную книгу России.
-
3. Видовой состав, показатели обилия и биото-пическое распределение дождевых червей отражают гидротермические особенности сообществ пояса широколиственных лесов терского варианта поясности. Значительную роль в формировании структуры фауны играет также тип растительной формации. Он, в частности, определяет видовой состав люмбрицид, зависящий от их пищевой специализации (примером является D. octaedra , встречающийся исключительно под древостоем).
-
4. Изучение сезонной динамики численности в разных биотопах показывает разнообразие экологических стратегий дождевых червей при переживании неблагоприятных погодных условий: использование микростаций в зимний период (подстилочные D. octaedra, D. rubidus tenuis , E. tetraedra tetraedra , почвенно-подстилочные D. schmidti и E. fetida ), формирование в годичном жизненном цикле двух фаз активной жизнедеятельности (большинство дождевых червей, например – собственно почвенный D. tellermanica ), как вариант – переход летней диапаузы в зимнюю, в отдельные годы период покоя может достигать 810 месяцев (собственно почвенный A. rosea ).
rosea , хорошо приспособленные к переживанию жестких гидротермических условий. В экотонном по режиму увлажнения биотопе (орешник-рододендрон) собраны D. schmidti и D. octaedra.
Выражаю огромную благодарность Ф.А. Тем-ботовой за организацию экспедиций, Ф.А. Тембото-вой и В.И. Ланцову за постоянное внимание и руко- водство работой, Т.С. Всеволодовой-Перель – за бесценные для меня консультации и важные замечания. Благодарю В.И. Ланцова, О.В. Умерову, З.М. Юсупова, А.А. Мокаеву за помощь в разборе почвенных проб.
Работа выполнена при финансовой поддержке программы фундаментальных исследований Президиума РАН «Биоразнообразие и динамика генофондов» и гранта РФФИ-Юг № 06-04-96711.
Список литературы Сезонная активность дождевых червей (Oligochaeta, Lumbriсidae) пояса широколиственных лесов Кабардино-Балкарского государственного высокогорного заповедника и прилегающих территорий (Центральный Кавказ)
- Квавадзе, Э.Ш. Дождевые черви (Lumbricidae) Кавказа. -Тбилиси: Мецниереба, 1985. -283 с.
- Соколов, В.Е. Млекопитающие. Насекомоядные/В.Е. Соколов, А.К. Темботов. -М.: Наука, 1989. -С. 3-27.
- Гиляров, М.С. Учет крупных беспозвоночных (мезофауны)//Методы почвенно-зоологических исследований. -М.: Наука, 1975. -С. 12-29.
- Всеволодова-Перель, Т.С. Дождевые черви фауны России: кадастр и определитель. -М.: Наука, 1997. -102 с.
- Стриганова, Б.Р. Животное население почв бореальных лесов Западно-Сибирской равнины/Б.Р. Стриганова, Н.М. Порядина. -М.: Товарищество научных изданий КМК, 2005. -234 с.
- Красная книга Российской Федерации. -М.: АСТ, 2001. -С. 34.
- Rapoport, I.B. Morpfo-ecological forms of Dendro-baena schmidti Мichaelsen, 1907 (Oligochaeta, Lumbricidae) of North Caucasus//Fouth International Oligochaete Taxonomy Meetings. Book of Abstracts. -Diyarbakir, Turkey, 2009. -Р. 39.
- Димо, Н.А. Земляные черви в почвах Средней Азии//Почвоведение. -1938, 4. -С. 494-526.
