Сезонная динамика активности и жизненные циклы жужелиц подрода Petrophilus chaudoir Восточного Саяна (Coleoptera, Carabidae, Pterostichini)
")
Автор: Хобракова Л.Ц.
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Зоология
Статья в выпуске: 3, 2007 года.
Бесплатный доступ
В этой работе приводятся описания сезонной активности 4-го вида наземных жуков из подродов Petrophilus родов Pterostichus: P. subaeneus Chaud., P. montanus (Motsch.), P. turanensis Jedl., P. dilutipes (Motsch. ) из горной тайги Центрально-Саянского плато. Жизненные циклы наземных жуков реконструируются на основе демографической структуры популяций.
Короткий адрес: https://sciup.org/148183739
IDR: 148183739
Текст научной статьи Сезонная динамика активности и жизненные циклы жужелиц подрода Petrophilus chaudoir Восточного Саяна (Coleoptera, Carabidae, Pterostichini)
Изучение онтогенеза и жизненных циклов почвообитающих беспозвоночных - одно из актуальных направлений в отечественной почвенной зоологии, основанной академиком М.С. Гиляровым (1957). После работы Ларссона (Larsson, 1939), предложившего первую классификацию жизненных циклов жужелиц, это направление в изучении жужелиц заметно прогрессировало. Однако деление жужелиц по типам развития на весенние и осенние виды оставалось неизменным. Основным диагностическим признаком оставалась зимующая стадия: у весенних -имаго, а у осенних - личинки. 40 лет спустя был выделен еще один тип сезонной активности -мультисезонный (Шарова, Душенков 1979). Исследования последних лет показывают, что эти схемы активности и развития жужелиц меняются в зависимости от географического положения занимаемого биотопа и климатических условий (Шарова, 1990; Шарова, Филиппов, 2002). Одногодичное развитие жужелиц широко распространено в умеренной зоне. В бореальных (Houston, 1981) или альпийских условиях развитие жужелиц может растягиваться на несколько лет (Forsskal, 1972; De Zordo, 1979; Refseth, 1980, 1984; Schatz, 1994).
Расшифровкам жизненных циклов жужелиц Европы и западной части России на основе данных по динамике демографической структуры имаго посвящено значительное количество работ (Andersen, 1984; Wallin, 1987; Refseth, 1986; Макаров, Черняховская, 1989, 1990; Sharova, Denisova, 1995; Шарова, Денисова, 1996, 1997; Маталин, 1997а, 19976, 1998а, 19986; Шарова, Филиппов, 2003).
На территории Сибири такие исследования практически не проводились. Биология размножения у широко распространенных сибирских видов жужелиц в условиях Южного Прибайкалья изучалась ВТ. Шиленковым (1978), а в горных условиях практически не исследована. Имеются данные о разведении отдельных видов сибирских жужелиц и развитии их личинок (Берлов, Берлов, 1984,1989).
При расшифровке жизненного цикла обращалось внимание на общий ход сезонной динамики и ее продолжительность, сроки размножения и пики яйцепродукции самок, что позволило выявить сезонность размножения жужелиц. По составу перезимовавших стадий развития можно судить о поливариантности жизненного цикла жужелиц, то есть о составе разновозрастных группировок в популяции и их сезонном развитии (Макаров, Черняховская, 1989, 1990; Макаров, 1991; Шарова, Денисова, 1997; Маталин, 1997а).
Материал и методы
Основой для статьи послужил материал, собранный стационарно с мая по октябрь 2001 г. в горно-таежном лиственничном поясе Окинского (Центрально-Саянского) нагорья на профиле в долине р. Ехэ-Хэрэгтэ (правый приток р. Ока) на разных склонах и высотах.
Для характеристики демографической структуры вида проанализированы популяции из 9 биотопов на склонах северной и южной экспозиций от вершины до его подножия.
Склон северной экспозиции: вершина, h=2000 м, лиственничник овсяницево-лишайниковый (2а); верхняя транзитная часть склона, h=1850 м, лиственничник кустарниковый (2г); нижняя транзитная часть склона, h=1650 м, лиственничник овсяницево-мохово-злаковый (2е); аккумулятивная часть склона, h=1500 м, лиственничник бруснично-зеленомошно-лишайниковый (2ж).
Склон южной экспозиции: вершина, Ь=2000 м, лиственничник кедровый лишайниково-моховый (2о); верхняя транзитная часть, h=185O м, лиственничник лишайниково-брусничнозеленомошный (2н); нижняя транзитная часть склона, h = 1650 м, лиственничник кустарниковоразнотравный (2и); аккумулятивная часть склона, h=1500 м, лиственничник моховоразнотравный (2к). -
В долине: h=l500 м, лиственничник кустарниково-разнотравный (2и).
Кроме того, учитывались данные предварительных исследований сезона 2000 г. Жуков и личинок отлавливали почвенными ловушками с 4%-ным формалином. В каждом биотопе устанавливалось по 10 ловушек. Выборку жуков проводили один раз в декаду. Для доминантных видов был проведен анализ демографической структуры популяций по репродуктивному состоянию и возрасту имаго обоих полов по методике X. Валлина (Wallin, 1987).
Для анализа сезонной динамики активности и расшифровки жизненных циклов жужелиц рода Pterostichus выбраны 4 массовых вида из подрода Petrophilus-. Р. montanus (Motsch.) (637 экз.), Р. dilutipes (Motsch.) (420 экз.), Р. subaeneus Chaud. (203 экз.), Р. turanensis Jedi. (91.экз.).
При вычислении индекса соотношения полов (ИСП) применяли формулу Шустека: ИСП = (f-mW+m), где/- число самок, т - число самцов.
Результаты ■ .
Pterostichus dilutipes (Motschulsky, 1844)
Таежный вид, широко распространенный в Сибири от Предуралья на западе до Южной Якутии на востоке и гор Монголии на юге. В Прибайкалье Р. dilutipes - один из наиболее распространенных лесных видов. Он поселяется в самых различных по составу лесах, но избегает слишком сухих и светлых лесов и чистых мертвопокровных сосняков (Шиленков, 1978).
В долинах рек Ока и Ехэ-Хэрэгтэ Р. dilutipes отмечается в 30 биотопах и обитает практически во всех типах лиственничных лесов на разных склонах. В районе исследования этот вид активен с конца мая (третья декада мая) до конца лета (третья декада августа) на склоне северной экспозиции (2а) и низкой пойме в долине (2и), а у подножия склона южной экспозиции (2к) активность вида смещается на период с первой декады июня до первой декады сентября. Таким образом, активность жуков на вершине склона северной экспозиции смещена на середину лета, в пойме долины активность вида приходится на весенне-раннелетний период, у подножия склона южной экспозиции - на раннелетний период. В исследованных биотопах продолжительность сезонной активности вида везде насчитывает 10 декад (табл. 1).
Таблица 1
Сроки и продолжительность сезонной активности Pterostichus dilutipes
|
Биотопы |
Период активности |
Продолжительность |
|
2а |
3 дек V - 3 дек VIII |
10 дек |
|
2и |
3 дек V - 3 дек VIII |
10 дек |
|
2к |
1 дек VI - 1 дек IX |
10 дек |
Графики сезонной активности включают два пика. Первый пик активности имеет разные сроки в зависимости от высоты и экспозиции склона, на вершине склона северной экспозиции (2а) он приходится на первую декаду июля, в низкой пойме в долине (2и) -- на первую декад)' июня и у подножия склона южной экспозиции - на вторую декаду июня. Этот пик активности жуков раньше отмечается в долине, а позже наступает на вершине склона северной экспозиции. Второй пик активности на склоне северной экспозиции (2а) отмечен в третьей декаде июля, в долине (2и) зарегистрирован во второй декаде июля и на склоне южной экспозиции (2к) приходится на первую декаду августа. Этот пик сезонной активности раньше отмечается в долине, а в более поздние сроки - у подножия склона южной экспозиции.
Пики сезонной активности Pterostichus dilutipes в различных биотопах
|
Пики активности |
Биотопы |
||
|
2а |
2и |
2к |
|
|
1 |
1 дек. VII |
1 дек. VI • |
2 дек. VI |
|
' 2 |
3 дек. VII |
2 дек. VII |
1 дек. VIII |
В половозрастной структуре популяций Р. dilutipes соотношение самок и самцов на склонах с разной экспозицией меняется. На вершине склона северной экспозиции преобладают самки, в долине преобладают самцы и только у подножия склона южной экспозиции отмечается более выровненное соотношение полов и здесь наблюдается наибольшая численность вида (табл. 3). Во всех изученных биотопах в начале сезона активности отмечаются самки и самцы имматур-ного, генеративного и постгенеративного возрастов, только на вершине склона северной экспозиции в начале сезона отмечаются постгенеративные имаго обоих полов.
Таблица 3
Индексы соотношения полов Pterostichus dilutipes __________________
|
Биотопы |
Число экземпляров |
исп |
||
|
всего |
самок |
самцов ■ |
||
|
2а |
66 |
41 |
25 |
0,24 |
|
2и |
116 |
46 |
70 |
-0,20 |
|
2к |
126 |
64 |
62 |
0,01 |
В Восточном Саяне активность Р. dilutipes проявляется с конца весны - начала лета и заканчивается в конце лета - начале осени. Продолжительность сезонной активности составляет 10 декад, несмотря на разные сроки вступления и завершения активности. С конца мая отмечен большой подъем активности, связанный с размножением, который длится 6 декад. В размножении участвуют имматурные, генеративные и постгенеративные имаго. Несмотря на растянутый период размножения в первой половине сезона, отмечаются два следующих друг за другом пика с небольшим преобладанием последнего. Максимум яйцепродукции (в третьей декаде июня - первой декаде июля), совпадает по времени со вторым пиком размножения. В течение всего августа и начала осени жуки становятся малоактивными, и уходят на зимовку в куколочных колыбельках.
Сезонная динамика демографической структуры Pterostichus dilutipes
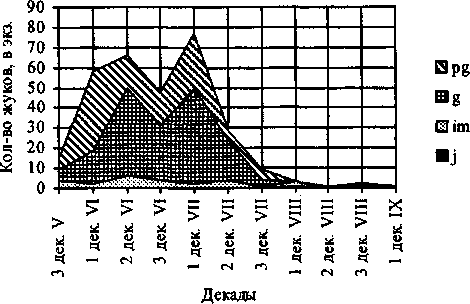
Рис. 1.
В районе исследования Р. dilutipes имеет одногодичный жизненный цикл с весенне-летним размножением и максимумом яйцепродукции в летний период. Характерно отсутствие активности жуков осенью в связи с ранним уходом на зимовку. Вид с поливариантным развитием, с зимующими имматурными, генеративными и постгенеративными имаго, размножающими в следующем сезоне.
В лесах Южного Прибайкалья этот вид имеет одногодичный цикл развития с весенним и осенним размножением (Шиленков, 1978). Жуки активны с середины мая, достигают максимума численности в июле. Яйцекладка начинается уже в конце мая и сильно растянута вплоть до осени. Наиболее интенсивно откладка яиц происходит в июле. В связи с сильно растянутым периодом размножения зимуют как молодые жуки, которые отрождаются в августе - сентябре, так и личинки. Взрослые личинки зарегистрированы в середине мая, окукливание происходит в первой декаде июня, а имаго отрождаются во второй декаде июня. Таким образом, первый пик активности дают жуки, зимовавшие в имагинальной стадии и приступающие к яйцекладке весной, а второй пик - жуки, зимовавшие в личиночной фазе, отродившиеся в начале лета и приступающие к яйцекладке только в июле.
Горно-лесной вид, обитающий в Алтае-Саянской горной системе от Восточного Алтая до Хамар-Дабана. В районе исследования в долинах рек Ока и Ехэ-Хзрэггэ этот вид обитает в 19 биотопах
В районе исследования этот вид многочислен в лиственничных лесах верхних транзитных частей обоих склонов, и активен с конца мая до конца августа (рис. 2). Общая продолжительность сезонной активности составляет до 10 декад (табл. 4). Начало сезонной активности отмечено на склоне северной экспозиции на вершине (2а) и верхней транзитной части (2г) с третьей декады мая, а в нижней транзитной части склона (2е) - с первой декады июня. Продолжительность активности составляет 3 и 6 декад, причем наибольшая её длительность - на вершине северной экспозиции. На склоне южной экспозиции сезонная активность начинается в третьей декаде мая, и заканчивается в третьей декаде июня (на вершине склона), или в третьей декаде августа (в верхней транзитной части).
Таблица 4
Сроки и продолжительность сезонной активности Pterostichus subaeneus
|
Склон северной экспозиции |
Склон южной экспозиции |
||||
|
Биотопы |
Сроки активности |
Продолжительность |
Биотопы вниз по склону |
Сроки активности |
Продолжительность |
|
2 а |
3 дек. V - 2 дек.VII |
6 дек. |
2о |
3 дек. V - 3 дек. VI |
4 дек. |
|
2г |
3 дек. V-2 дек.VI |
3 дек. |
2н |
3 дек. V - 3 дек. VII! |
10 дек. |
|
2е |
1 дек. VI - 3 дек. VI |
3 дек. |
- |
- |
• |
В сезонной динамике активности вида отмечается 2-3 пика сезонной активности (табл. 5). На склонах северной и южной экспозиций первый пик отмечен в третьей декаде мая - первой декаде июня, причем на вершинах этот срок наступает раньше. Второй пик активности наступает в третьей декаде июня на всех склонах и только на вершине северной экспозиции приходится на один месяц позже. Третий пик активности регистрируется только на склоне южной экспозиции - в третьей декаде августа.
Таблица 5
Пики сезонной активности Pterostichus subaeneus
|
Пики активности |
Склон северной экспозиции |
Склон южной экспозиции |
|||
|
2а |
2г |
2е |
2о |
2н |
|
|
1 |
3 дек. V |
3 дек. V |
1 дек. VI |
3 дек. V |
1 дек. VI |
|
2 |
1 дек. VII |
- |
3 дек. VI |
3 дек. V] |
3 дек. VI |
|
3 |
• |
- |
- |
- |
3 дек. VIH |
Половозрастная структура популяций Р. subaeneus меняется в зависимости от положения биотопа. На вершинах северной и южной экспозиций склонов преобладают самцы (ИСП соответственно составляет -0,10 и -0,07 (табл. 6). Ниже по склонам обоих экспозиций отмечено преобладание самок.
Индексы соотношения полов Pterostichus subaeneus
|
Склон северной экспозиции |
Склон ЮЖНОЙ ЭКСПОЗИЦИИ |
||||||||
|
Биотопы |
Число экземпляров |
исп |
Биотопы |
Число экземпляров |
исп |
||||
|
всего |
самок |
самцов |
всего |
самок |
самцов |
||||
|
2а |
29 |
13 |
16 |
-0,10 |
2о |
39 |
18 |
21 |
-0,07 |
|
2г |
31 |
17 |
14 |
0,09 |
2н |
54 |
33 |
21 |
0,22 |
|
2е |
50 |
26 |
24 |
0,04 |
- |
- |
• |
- |
- |
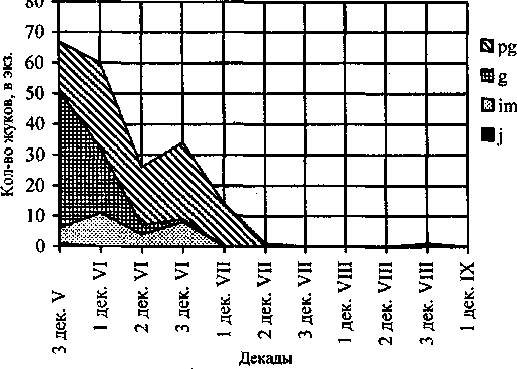
По нашим данным Р. subaeneus активен с конца мая до конца августа, общая продолжительность периода активности составляет 3-10 декад. Яйцекладка длится с третьей декады мая по третью декады июня с максимумом яйцепродукции в конце мая - середине июне. Часть генеративных особей, после размножения погибает, что подтверждается находками мертвых жуков. Период репродуктивной активности составляет 5 декад, затем жуки становятся малоактивными, и в конце лета (третья декада августа) появляются ювенильные особи. Оптимальные условия обитания этого вида - верхние транзитные части северной и южной экспозиций склонов, что отмечается на особенностях жизненного цикла.
Таким образом, жизненный цикл является одногодичным с весенним размножением. Максимальный пик активности жуков отмечен в мае, активность к середине лета завершается. Зимуют имаго в имматурном и постгенеративном состоянии. Размножение всех группировок происходит одновременно - при переходе в генеративное состояние. Приспособление к суровым условиям обитания в горах проявляется в укорочении сроков активности в период размножения до 5 декад; синхронности размножения всех перезимовавших разновозрастных групп имаго и развития преимагинальных фаз; слабая активность имаго во второй половине сезона в связи с их ранним уходом на зимовку. .
В Южном Прибайкалье (Шиленков, 1978) это вид с весенним одногодичным развитием. Период яйцекладки отмечается в июне. Период отрождения молодых жуков наблюдается в июле. Зимуют в имагинальной стадии.
Pterostichus montanus (Motschulsky, 1844)
Горно-лесной вид, распространенный от Полярного Урала до Охотского побережья, на юге до Северной Монголии. В районе исследования этот вид отмечен в 34 биотопах в долинах рек Ока и Ехэ-Хэрэгтэ.
Сезонная активность Р. montanus проявляется с третьей декады мая по первую декад)' сентября (рис. 3). Длительность этого периода составляет 8-11 декад (табл, 7). Хотя во всех исследованных биотопах начало активность совпадает, её окончание активности приходится на разные сроки и зависит от экспозиции склона, его освещенности и от микроклиматических условий, что особенно заметно на склоне северной экспозиции. Так, на вершине этот срок приходится на вторую декаду августа, в верхней транзитной части - на первую декаду сентября, в нижней транзитной части - на третью декаду августа и в аккумулятивной части - на первую декаду августа. На склоне южной экспозиции период активности везде заканчивается в конце лета (в третьей декаде августа) и только у его подножия - в начале осени (первая декада сентября). Таким образом, продолжительность сезонной активности на склонах северной экспозиции составляет 8-11 декад, а на склоне южной экспозиции - 10-11 декад.
Таблица 7
Сроки и продолжительность сезонной активности Pterostichus montanus
|
Биотопы |
Склон северной экспозиции |
Кол-во декад |
. Биотопы |
Склон южной экспозиции |
Кол-во декад |
|
■ 2а |
3 дек V - 2 дек VIII |
9 |
2о |
3 дек V - 3 дек VIII |
10 |
|
2г |
3 дек V - 1 дек IX |
И |
2н |
3 дек V - 3 дек VIII |
10 |
|
2е |
3 дек V - 3 дек VIII |
10 |
2и |
3 дек V - 3 дек VIII |
10 |
|
2ж |
3 дек V - 1 дек VIII |
8 |
2к |
3 дек V - 1 дек IX |
11 |
Сезонная динамика демографической структуры Pterostichus subaeneus
Рис, 2.
В сезонной динамике вида на разных склонах отмечено 3 пика активности (табл. 8). Первый пик на склоне северной экспозиции отмечен с конца мая до середины июня. Причем в верхних частях склона (2а, 2г) он запаздывает на 1-2 декады и регистрируется в начале и середине июня, в отличие от нижних частей склона (2е, 2ж), где этот пик приходится на конец мая.
На склоне южной экспозиции первый пик активности приходится на конец мая - начало июня, причем на вершине этот пик активности отмечен на одну декаду раньше (в конце мая), а во всех остальных частях склона приходится на начало июня. Второй пик активности на склоне северной экспозиции приходится на конец июня - начало июля. В верхних частях склона (2а, 2е) он приходится на середину лета - первую декаду июля, а у подножия склона отмечается на декаду раньше - в конце июня. В верхней транзитной части склона этот пик не отмечается. На склоне южной экспозиции второй пик активности регистрируется в конце июня - начале июля, причём на вершине он отмечен в начале июля - первая декада июля, а в нижних частях склона приходится на одну декаду раньше (третья декада июня). У подножия склона такой пик не отмечен. Третий пик активности в транзитных частях склона северной экспозиции не выражен, а на вершине и у подножия приходится на конец июля. На склоне южной экспозиции третий пик активности выражен нечетко и приходится с первой по третью декады августа, у подножия склона он не отмечается.
■ Таблица 8 Пики сезонной активности Pterostichus montanus
|
Пики активности |
Склон северной экспозиции |
Склон южной экспозиции |
||||||
|
2а |
2г |
2е |
■ 2ж |
2о |
2н |
2и |
2к |
|
|
1 |
1дек. VI |
2 дек. VI |
3 дек. V |
3 дек. V |
3 дек. V |
1 дек. VI |
1 дек. VI |
1 дек. VI |
|
2 |
1 дек. VII |
- |
1 дек. VII |
3 дек. VI |
1 дек. VII |
3 дек. VI |
3 дек. VI |
- |
|
3 |
3 дек. VII |
- |
- |
3 дек. VII |
1-3 дек. VIII |
2-3 дек. VIII |
2-3 дек. "VIII |
■ |
При анализе половозрастной структуры популяций Р. montanus отмечено, что на склоне северной экспозиции число самок превалирует везде, кроме подножия склона (2ж), где преобладают самцы (табл. 9). Наиболее выровненная половая структура вида отмечена в верхней транзитной части склона (2г). На склоне южной экспозиции преобладают самцы, причем их численное обилие нарастает сверху вниз по склону. Наиболее выровненное соотношение самок и самцов отмечено в верхней транзитной части склона. В целом, на склоне северной экспозиции преобладают самки, а на склоне южной экспозиции - самцы. Соотношение полов, близкое к 1, на обоих склонах зарегистрировано в верхней транзитной части (2г и 2н).
Индексы соотношения полов Pterostichus montanus
|
Склон северной экспозиции |
Склон южной экспозиции |
||||||||
|
Биотопы |
Число экземпляров |
исп |
Биотопы |
Число экземпляров |
ИСП |
||||
|
всего |
самок |
самцов |
всего |
самок |
самцов |
||||
|
2а |
94 |
58 |
36 |
0,23 |
2о |
56 |
25 |
31 |
-0,10 |
|
2г |
157 |
77 |
80 |
0,01 |
2н |
59 |
27 |
32 |
-0,08 |
|
2е |
54 |
28 |
26 |
0,03 |
2и |
59 |
9 |
16 |
-0,28 |
|
2ж |
73 |
29 |
44 |
-0,20 |
2к |
67 |
19 |
48 |
-0,43 |
На склоне северной экспозиций в начале сезона отмечаются иммасгурные и генеративные, иногда ювенильные имаго самок и самцов, а на склоне южной экспозиции - имматурные, генеративные и постгенеративные имаго обоих полов. У самцов продолжительность генеративного возраста больше, чем у самок на склонах северной экспозиции и верхних частях южной экспозиции. В конце сезона активности отмечаются молодые жуки.
По нашим данным Р. montanus активен с третьей декады мая по первую декаду сентября и общая продолжительность сезонной активности составляет 8-11 декад (рис. 3). Весной, с конца мая, в ловушках отмечаются имаго всех возрастов, которые переходят в генеративное состояние и с конца второй декады июля отмечается период репродуктивной активности, связанный с весенне-летним размножением вида, который длится 5 декад. Продолжительность яйцекладки на обоих склонах отличается сроками, на склоне северной экспозиции она длится с конца мая до конца июля, а на склоне южной экспозиции заканчивается раньше на одну. Смещение сроков максимальной яйцепродукции более заметно по склону северной экспозиции: от вершины до его подножия они меняются с третьей декады мая по вторую декаду июня. На склоне южной экспозиции максимум яйцепродукции во всех биотопах в основном приходится на первую декаду июля.
Сезонная динамика демографической структуры Pterostichus montanus
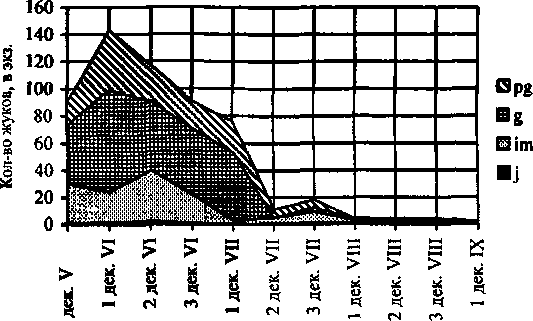
Декады
Рис. 3.
После большого и продолжительного периода размножения, с середины июля до начала августа, отмечается очень небольшой пик активности, представленный имматурными и постгенеративными имаго. В течение всего августа отмечаются в ловушках только ювенильные и имматурные особи.
Таким образом, жизненный цикл Р. montanus является одногодичным с весеннераннелетним размножением. Зимуют имаго в имматурном, постгенеративном состояниях, которые размножаются одновременно в мае, июне с пиком яйцепродукции в июне. Молодое поколение появляется в середине июля, но малоактивно. Можно предположить, что большинство ювенильных жуков остается на зимовку в куколочных колыбельках. Жизненный цикл полива- риантен по возрастному составу размножающихся жуков, зимующих в имматурном и постгенеративном состоянии.
Приспособление жизненного цикла к суровым горным условиям проявляется в ускорении популяционного развития (весенне-раннелетний период размножения в среднем занимает 5 декад), а молодое поколение во второй половине сезона мало активно и остается в куколочных колыбельках на зимовку.
По данным В.Г. Шиленкова (1978), в условиях Южного Прибайкалья этот вид с весенним одногодичным развитием. Период яйцекладки отмечен в июне. Период отрождения молодых жуков в июле - августе. Зимуют имаго.
Pterostichus turanensis Jedlicka, 1959
Горно-лесной вид - эндемик Восточного Саяна. В районе исслсдойния зарегистрирован в 4 биотопах, преимущественно на склоне северной экспозиции на каменистых участках.
Сезонная активность Р. turanensis отмечена с третьей декады мая до второй декады августа, её общая длительность составляет 9 декад (рис. 4). Единственный пик активности приходится на первую декаду июня.
В половозрастной структуре популяции Р. turanensis имеются некоторые особенности. В половой структуре вида самцы заметно преобладают над самками (индекс соотношения полов -0,78). В возрастной структуре самок и самцов заметны следующие отличия: у самок в размножение вступают имаго, зимовавшие в имматурном и постгенеративном, а у самцов в начале сезона отмечены только зимовавшие имматурные имаго, которые переходят в генеративное состояние.
В начале сезона отмечается репродуктивная активность Р. turanensis длительностью до 4 декад. Яйцекладка наблюдается с третьей декады мая по вторую декаду июня. Максимум яйце-продукции отмечен в первой декаде июня. Вероятно, после размножения жуки погибают, так как постгенеративные имаго после середины июня не отмечались. В середине лета, вероятно, происходит преимагинальное развитие вида, так как в августе появляются ювенильные и имматурные жуки, отродившиеся из летних личинок.
Сезонная динамика демографической структуры Pterostichus turanensis
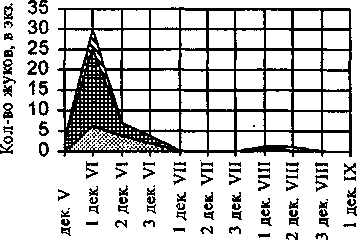
spg Sg Elim ■j
Декады
Рис. 4
Таким образом, жизненный цикл Р. turanensis в Восточном Саяне - одногодичный с весенне-раннелетним размножением, с зимующими имматурными имаго.
В.Г. Шиленков охарактеризовывал этот вид как весенний с одногодичным развитием. Период размножения приходится на июнь, а отрождение молодых жуков отмечалось в июле (Шиленков, 1978). ■ ■
Обсуждение результатов ■ .
Изучена сезонная динамика активности и демографическая структура популяций 4 доминантных видов жужелиц горно-таежного пояса в долине р. Ехэ-Хэрэгтэ на склонах северной и южной экспозиций. Полученные данные сведены в графики сезонной активности половозрастной структуры популяций, репродуктивного потенциала самок в разных частях склонов (вершина, верхняя транзитная, нижняя транзитная и аккумулятивная части склона). Выявлены 2 основных типа сезонной динамики активности жужелиц в условиях горно-таежного пояса: весенний и мультисезонный.
Весенние виды — Р, subaeneus, Р. montanus, Р. turanensis - имеют высокий пик активности весной в период размножения (конец мм - начало июня). У этих видов во второй половине сезона отмечается небольшой подъем активности за счет молодого поколения имаго. По сравнению с сезонной динамикой весенних видов европейской части России, жужелицы в горах Сибири имеют более поздние сроки весеннего размножения и слабую активность молодого поколения во второй половине сезона в связи с ранним уходом на зимовку. Такой ход сезонной активности весенних видов жужелиц определяется более коротким вегетационным сезоном в Сибири, холодной весной и ранними осенними заморозками.
МультисезонныЙ тип сезонной динамики активности характерен для Pterostichus dilutipes. Активность этого вида охватывает наиболее теплую часть сезона с начала июня по август. Мультисезонные циклы со средне-летним максимумом (Pterostichus dilutipes) характеризуются 2 пиками размножения: большим в середине лета за счет зимовавших имаго и малым - во второй половине лета за счет размножения имаго, отродившихся из зимовавших личинок. Зимуют имаго разных возрастов и личинки.
Сезонная динамика видов изменяется на разных склонах и высотах. Доминантные виды жужелиц, широко встречающиеся на разных склонах и высотах, обнаруживают изменение сезонной динамики активности. Так, например, у одного и того же вида в биотопах вершинной части склона по сравнению с биотопами в нижней части склона сроки сезонной динамики активности укорачиваются, а пики максимальной активности в период размножения смещаются. Такая закономерность изменения сезонной динамики активности на разных высотах склона отмечена для многих лесных видов.
Выявлены адаптации к развитию доминантных видов жужелиц в горных условиях Южной Сибири. Для одногодичных видов с весенним размножением характерно ускорение популяционного развития в связи поздней весной и ранними осенними заморозками.
Список литературы Сезонная динамика активности и жизненные циклы жужелиц подрода Petrophilus chaudoir Восточного Саяна (Coleoptera, Carabidae, Pterostichini)
- Берлов О. Э. Вернов Э.Я. К биологии прибайкальских жужелиц рода Carabus L. (Coleoptera Carabidae)//"Жесткокрылые Сибири". Иркутск, 1984. С. 67-77.
- Коробейников Ю.И. О некоторых особенностях сезонной динамики активности и размножения жужелиц в условиях Субарктики//"Проблемы почвенной зоологии. Матер, докл. X Всесоюз. совещ." Новосибирск, 1991. С. 127-128.
- Макаров К.В. Поливариантность жизненного цикла жужелиц (Coleoptera, Carabidae)//"Проблемы почвенной зоологии. Матер, докл. X Всесоюз. совещ." Новосибирск, 1991. С. 132.
- Макаров К.В., Черняховская Т.А. Изменчивость сезонной динамики жужелицы Pterostichus melanarius (Coleoptera, Carabidae) в разных типах леса//"Экологические вопросы рационального природопользования: Тез. докл. VIII конф. молод, уч.-биол." Рига, 1989. С. 55-56.
- Макаров К.В., Черняховская Т.А. Фенология развития и структура популяции Loricera pilicornis (F.) (Coleoptera, Carabidae) в условиях агроценоза//"Структура и динамика популяций почвенных и наземных беспозвоночных животных". М.: МПГУ, 1990. Т. 1. С. 21-32.
- Маталин А.В. Особенности жизненного цикла Pseudoophonus (s. str.) rufipes Deg. (Coleoptera, Carabidae) в Юго-Западной Молдове//Изв. АН. Сер. биол., 1997. № 4. С. 455-466.
- Маталин А.В. Жизненные циклы жужелиц трибы Harpalini (Coleoptera, Carabidae) в условиях Европейского субконтинента: возможные пути их становления и трансформации//"Проблемы энтомологии в России. Сб. научи, тр. XI съезда РЭО 23-26 сентября 1997". СПб., 1998. Т. 2. С. 25-26.
- Маталин А.В. Поливариантность жизненного цикла Harpalus (s. str.) affinis Schrank и ее адаптивное значение//Изв. АН. Сер. биол., 1998. № 4. С. 496-505.
- Хобракова Л.Ц. Жизненный цикл жужелицы Carabus loschnikovi F.-W. в лиственничных лесах Окинского нагорья (Восточный Саян)//"Проблемы почвенной зоологии. Матер. III (XIII) Всерос. сов. по почв, зоол." Йошкар-Ола, 2002. С. 187-188.
- Шарова И.Х., Денисова М.И. Поливариантность сезонного развития двух видов жужелиц рода Calathus Bonelli (Coleoptera, Carabidae) в лесостепи Центральной России//Докл. АН СССР. 1996. Т. 348. № 1. С. 140-142.
- Шарова И.Х., Денисова М.И. Сезонная динамика лесных популяций жужелиц рода Pterostichus (Coleoptera, Carabidae)//Зоол. журн. 1997. Т. 76. № 4. С. 418-427.
- Шарова И.Х., Душенков В.М. Типы развития и типы сезонной активности жужелиц (Coleoptera, Carabidae)//"Фауна и экология беспозвоночных". М.: МГПИ, 1979. С. 15-25.
- Шарова И.Х., Филиппов Б.Ю. Особенности жизненных циклов жужелиц (Coleoptera, Carabidae) в северной тайге//Зоол. журн., 2003. Т. 82. № 2. С. 229-238.
- Шарова И.Х., Хобракова Л.Ц. Сезонная динамика жизненного цикла Carabus loschnikovi F.-W. (Coleoptera, Carabidae) в высокогорьях Восточных Саян//"Тез. докл. XII съезд РЭО". СПб., 2002. С. 379-380.
- Шарова И.Х., Хобракова Л.Ц. Особенности жизненных циклов Pterostichus montanus (Motschulsky, 1844) и Carabus loschnikovi (Fischer-Waldheim, 1822) (Coleoptera, Carabidae) в условиях горно-таежного пояса Восточного Саяна//Известия РАН. Сер. биол. Зоология. 2005. № 1. С. 1-11.
- Шиленков В.Г. Особенности биологии массовых видов жужелиц (Coleoptera Carabidae) фауны южного Прибайкалья//Энтом. обозр. 1978. Вып. 2. С. 290-301.
- Andersen J.M. A re-analysis of the relationships between life cycle patterns and the geographical distribution of Fennoscandian carabid beetles//J. Biogeography. 1984. V. 11. № 6. P. 479-489.
- Brandmayr P., Zetto Blandmayr T. Phenology of ground beetles and its ecological significance in some of the main habitat types of Southern Europe//"Carabids beetles: Their adaptations and dynamics". Stuttgart-New York: Gustav Fisher Verl., 1986. P. 195-220.
- Butterfield J.E.L Changes in life-cycle strategies of Carabus problematicus over a range of altitudes in Northern England//Ecological Entomology. 1986. V. 11. №. 1. P. 17-26.
- Cardenas A.M., Hidalgo J.M. Seasonal activity and reproductive biology beetle of the ground beetle Carabus dufouri (Coleoptera, Carabidae)//European J. Entomol. 2000. V. 97. P. 329-338.
- Dijk Th.S. van. The age-composition of populations of Calathus melanocephalus L. Analysed by Studying Marked Individuals kept within fenced sites//Oecologia (Berlin). 1973. V. 12. P. 213-240.
- Dülge R. Seasonal activity of carabid beetles in wooded habitats in northwest Germany (Coleoptera, Carabidae)//"Carabid Beetles: Ecology and Evolution". Kluwer Academic Publ.: Dordrech. Series Entomologica. 1994. V. 51. P. 125-131.
- Hemmer J., Klenner M., Niesing H., Weber F. Life history phenomena in a subpopulation of Carabus auronitens F. from the Westphlian Lowland//"Carabids beetles: Their adaptations and dynamics". Stuttgart-New York: Gustav Fisher Verl., 1986. P. 439-464.
- Houston W.K. The life cycles and age of Carabus glabratus Paykull and C. problematicus Herbst (Col.: Carabidae) on moorland in northenrn England//Ecological Entomology. 1981. V. 6. P. 263-271.
- Jorum P. Life cycle and phenology of Carabus problematicus Herbst, 1786 in Denmark (Coleoptera, Carabidae)//Ent. Meddr. (K0benhavn). 1985. V. 53. P. 27-30.
- Kalas J.A. Species composition and seasonal activity patterns of Carabidae (Col.) in a small deciduous forest in western Norway//Fauna Norvegica (Oslo). Ser. B. 1985. V. 32. №. 1. P. 28-32.
- Kurka A. The life cycle of Agonum ruflcorne (Goeze) (Coleoptera, Carabidae)//Acta ent. Bohemoslov. 1976. V. 73. P. 318-323.
- Larsson S.G. Entwicklungstupen und Entwicklungszeiten der danischen Carabiden//Entomologiske Meddelelser. 1939.Bd. 20. P. 277-560.
- Neudecker Ch., Thiele H.U. Die jahreszeitliche Synchronisation der Gonadenreifung bei Agonum assimile Payk. (Coleopt. Carab.) durch Temperatur und Photoperiode//Oecologia (Berlin). 1974. V. 17. P. 141-157.
- Paarmann W. Seasonality and its control by environmental factors in tropical ground beetles (Col., Carabidae)//"Carabids beetles: Their adaptations and dynamics". Stuttgart-New York: Gustav Fisher Verl., 1986. P. 157-171.
- Refseth D. The life cycles and growth of Carabus glabratus and C. violaceus in Budalen, central Norway//Ecological Entomology. 1984. V. 9. №. 4. P. 449-455.
- Refseth D. Annual patterns of activity, reproduction and development in some Norwegian Carabidae (Col.)//Fauna Norvegica (Oslo). Ser. B. 1988. V. 35. №. 1. P. 21-30.
- Schats I. Life strategy of alpine carabid: Pterostichus jurinei (Coleoptera, Carabidae)//"Carabid Beetles: Ecology and Evolution". Kluwer Academic Publ.: Dordrech. Series Entomologica. 1994. V. 51. P. 213-217.
- Sharova I.Kh., Denisova M.I. Life cycles and reproduction rhythms of Calathus\erratus C.R. Sahlberg and C. micropterus Duftschmidt (Coleoptera Carabidae) in the forests of Russian forest-steppe zone//Russian Entomol. J. 1995. V. 4. №. 1-4. P. 159-165.
- Sota T. Life history patterns of Carabid beetles belonging to the subtribe Carabina (Coleoptera, Carabidae) in the Kinki district, Western Japan//Kontyu (Tokyo). 1985. V. 53. №. 2. P. 370-378.
- Szyszko J. Male-to-female ratio in Pterostichus oblongopunctatus (Coleoptera, Carabidae) as one characteristic of a population, P. 51-57. 1976.
- Thiele H. U. The control of larval hibernation and of adult aestivation in the Carabid beetles Nebria brevicollis F. and Patrobus atrorufus Stroem//Oecologia (Berlin). 1969. V. 2. P. 347-361.
- Wallin H. Distribution, movement and reproduction of Carabid beetles (Coleoptera, Carabidae) inhabiting cereal fields//"Plant protection reports and dissertation of the Swedish Univ. for Agric. Sci." Uppsala, 1987. V. 15. P. 3-19.
