Сезонная динамика дегидринов и температуры замерзания воды в талломах лишайника Lobaria pulmonaria в условиях средней тайги

Автор: Т.К. Головко, Н.Е. Коротаева, Р.В. Малышев, Г.Н. Табаленкова, М.А. Шелякин
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Статья в выпуске: 7 (83), 2025 года.
Бесплатный доступ
Исследовали сезонные изменения содержания белковдегидринов, оводненности и температуры замерзания воды в лишайнике Lobaria pulmonaria (лобария легочная). Впервые идентифицировано два дегидрина с мол. массой 40 и 43 кДа. Они присутствовали в талломах в течение всего года, что свидетельствует о конститутивной экспрессии данных белков. Наименьшее их содержание отмечали в наиболее благоприятный для роста весенний период. Показано увеличение содержания этих белков при десикации талломов в дневные часы летом и их повышенный уровень при переходе к холодному времени года. Содержание воды варьировало в пределах 45–70 % сырой массы лишайников. Температура кристаллизации воды в талломах изменялась в зависимости от сезона и составляла минус 10–11о С в наиболее холодный период. При снижении температуры фазовому переходу «вода – лед» подвергалось от 25 до 45 % всей содержащейся в талломах воды. Высказано предположение о существовании связи уровня дегидринов с оводненностью и температурой замерзания воды, и об их участии в формировании фракции сильно связанной воды.
Lobaria pulmonaria, таллом, белки-дегидрины, содержание воды, температура замерзания воды
Короткий адрес: https://sciup.org/149149451
IDR: 149149451 | УДК: 582.29:581.1:574.24 | DOI: 10.19110/1994-5655-2025-7-100-108
Seasonal dynamics of dehydrins and water freezing temperatures in the thalli of the lichen Lobaria pulmonaria in the middle taiga zone
Seasonal changes in the content of dehydrin proteins, water content, and freezing point of water in the lichen Lobaria pulmonaria have been studied. Two dehydrins with mol. weight of 40 and 43 kDa have been identified for the first time. They are available in the thalli throughout the year, which indicates the constitutive expression of these proteins. The lowest content of these dehydrins has been identified for the spring period, which is the most favorable time for the lichen growth. Desiccation of the thalli in warm summer days increases the content of the proteins. The change for the cold year period also provokes an increase in their concentration. The relative water content ranges from 45 % to 70 % fresh weight of the thalli. The water crystallisation temperature varies depending on the season and is minus 10–11 °C for the coldest period. As the temperature decreases, 25–45 % of the total water content in the thalli undergoes a phase transition from water to ice. It has been suggested that the level of dehydrins correlates with the water content and freezing point of the water, and that dehydrins contribute to the formation of a fraction of the strongly bound water.
Текст научной статьи Сезонная динамика дегидринов и температуры замерзания воды в талломах лишайника Lobaria pulmonaria в условиях средней тайги
Лишайники – уникальные фототрофные организмы. Они представляют собой ассоциацию гетеротрофного микобионта и фотосинтезирующего фотобионта. Согласно современным исследованиям [1], метагеном лишайников, помимо геномов микобионта (грибы) и фотобионта (зеленые водоросли и/или цианобактерии), содержит геномы множества видов бактерий. Лишайники проявляют высо- кую устойчивость к неблагоприятным воздействиям среды, выживают после обезвоживания, гипо- и гипертермии, в условиях повышенного ионизирующего излучения и других стресс-факторов, губительных для большинства живых организмов [2, 3]. Они доминируют в крайних местообитаниях (Арктика, Антарктида, пустыни, высокогорья), но наиболее благоприятны для обитания лишайни- ков леса, о чем свидетельствует высокое биологическое разнообразие лихенобиоты в бореальной зоне [4].
Лишайники – многолетние, медленнорастущие пой-килогидрические организмы. Благоприятными для их роста являются весенний и осенний периоды с умеренной температурой и высокой влажностью. Летом талломы лишайников часто подсыхают и становятся физиологически неактивными, но быстро восстанавливают свои функции при увлажнении. Так, например, у крупнолистоватого лишайника Lobaria pulmonaria в теплый и сухой период днем превалировало дыхание, а поглощение СО2 отмечали только в ранние утренние часы после гидратации талломов парами атмосферной влаги [5]. Проведенные нами ранее исследования показали, что отобранные зимой талломы после увлажнения и 2-3-часовой акклимации в комнатных условиях были способны ассимилировать СО2 со скоростью, сопоставимой с измеренной в благоприятный для роста период [6].
В данной работе представлены результаты изучения сезонной динамики уровня дегидринов, содержания и температуры замерзания воды в талломах лишайника L. pulmonaria в годичном цикле.
Дегидрины представляют собой высокогидрофильные белки, характеризующиеся наличием высококонсервативных Y-, S- и K-сегментов, богатых глицином участков и высокой термоустойчивостью [7, 8]. Они накапливаются в разных клеточных компартментах и тканях растений в ответ на осмотический стресс, вызванный дефицитом воды, низкой температурой, высокой засоленностью, и участвуют в стабилизации важных биологических макромолекул и мембран [9]. Имеются данные о роли этих белков в защите клеток при холодовой адаптации и формировании морозоустойчивости древесных растений [1013]. Количество дегидринов в растениях сильно варьирует в зависимости от вида и условий среды, а их мол. массы находятся в пределах 15-150 кДа [14]. Сведения о дегидринах лишайников весьма ограничены и касаются в основном фотобионта. По данным работ [15, 16], в лишайниковой водоросли Trebouxia erici постоянно присутствовали пять дегидринов с мол. массой 15, 18, 34, 40 и 120 кДа.
Материалы и методы
Lobaria pulmonaria (L.) Hoffm. (лобария легочная) – эпифитный крупнолистоватый лишайник. Вид распространен в бореальных, умеренных, горных и океанических районах северного полушария и тропических лесах Восточной и Южной Африки [17]. Охраняется в большинстве стран северной и центральной Европы.
Таллом L. pulmonaria гетеромерного типа с четко очерченным альгальным слоем под плотно прилегающими друг к другу гифами верхней коры. Под слоем клеток зеленой водоросли рыхло расположены гифы гриба, формирующие сердцевину. Нижний коровый слой имеет выросты (ризины), с помощью которых слоевище прикрепляется к субстрату. Толщина хорошо развитого таллома в зоне резин может достигать 300 мм, вне ее составляет около 200 мм [5].
Талломы (слоевища) лишайника отбирали в старовозрастном осиннике с примесью ели и пихты вблизи г. Сыктывкара (61о 34’ с. ш., 50о 33’ ‘в. д.). Сбор образцов осуществляли со стволов деревьев на высоте 1-3 м от земли в первой половине дня (10–11 ч) в разные сезоны года. Район исследований характеризуется умеренно-континентальным климатом, со среднегодовой температурой воздуха около +1о С. Среднесуточная температура самого теплого месяца (июля) – около +17о С, самого холодного (января) – минус 16о С. Годовое количество осадков составляет 600–700 мм. Наибольшее их количество выпадает летом и осенью, 60–80 мм за месяц, зимой и весной месячная норма осадков в 1,5-2 раза меньше. Переход средней суточной температуры через 0оС весной происходит во второй декаде апреля, осенью – в начале октября. Длительность безморозного периода составляет 180–190 дней, а продолжительность периода со среднесуточной температурой >5о С – около 160 дней [18].
Свежеотобранные талломы фиксировали в жидком азоте и высушивали лиофильно с помощью FreeZone 2.5 L (Labconso, США). Растворимый белок выделяли из ли-офильно высушенного материала. Для этого образцы (0,1 г), охлажденные в жидком азоте, тщательно растирали с кварцевым песком в присутствии метанола (1 : 4, v/v). Осадок, получившийся после центрифугирования гомогената (9000 g, 1 мин, 4о С), последовательно смешивали с 25%-ным хлороформом и метанолом, каждый раз отбрасывая жидкую фазу после центрифугирования (9000 g, 1 мин, 4о С). Общий белок, содержащийся в осадке, растворяли на водяной бане (3 мин.) в буфере для образца, содержащем 0,125 M Трис-HCl (pH 6,8), 10 % додецилсульфата натрия, 10 мМ β-меркаптоэтанола, 1 мМ ЭДТА, 20 % глицерина. Содержание белка в образце определяли с помощью флуориметра QubitTM (“Invitrogen”, США). Электрофорез в ПААГ и иммуноблоттинг проводили в системе miniProtean III (“Bio–Rad”, США) по прилагаемой инструкции. Для иммуноблоттинга использовали первичные антитела против специфической К-последовательности дегидринов (Agrisera, AS07 206, Швеция) и вторичные антитела, коньюгированные с щелочной фосфатазой (Sigma, США) [19]. Молекулярную массу выявленных дегидринов определяли с использованием белковых маркеров PageRuler Unstained Protein Ladder (Thermo Scientific, США) с помощью программного обеспечения Image Laboratory (версия 5.2) (Bio Rad, США). Содержание дегидринов оценивали с помощью программы Gel Analysis после перевода изображения мембраны в цифровой формат в условных единицах как отношение количества окрашенных пикселей данного белка к общей сумме окрашенных пикселей на мембране [20].
Температуру замерзания воды определяли калориметрически на калориметре DSC – 60 Shimadzu (Япония) [21]. Образцы (кусочки свежесобранных талломов массой до 50 мг) взвешивали, помещали в алюминиевый контейнер объемом 90 мм3 и охлаждали со скоростью 1 ºС/мин от +5º С до -30º С, непрерывно регистрируя интенсивность тепловыделения. Температуру кристаллизации воды определяли по началу пика фазового перехода. После проведения калориметрических измерений образцы высушивали при +105º С до постоянного веса. Содержание воды определяли по разнице сырой и сухой массы образцов. Количество воды, претерпевшей фазовый переход, – по формуле: m замер. воды = 335×Q, где Q – тепловой эффект при нуклеации воды (Дж), 335 – удельная теплота льдообразования в мДж/мг (для пресной воды).
Статистическую обработку данных проводили с помощью программы Statistica 10 (StatSoft Inc., США). В таблицах и рисунках приведены средние арифметические значения и их стандартные ошибки. Значимость различий содержания дегидринов определяли с помощью непараметрического критерия Манна-Уитни (р<0,05). Определения дегидринов выполнены в трех биологических повторностях. Измерения тепловыделения – с использованием четырех-пяти независимых образцов.
Результаты и их обсуждение
Анализ белков L. pulmonaria с применением электрофореза и иммуноблоттинга показал наличие в талломах двух четко выраженных белков-дегидринов с мол. массой 40 и 43 кДа. Эти среднемолекулярные дегидрины присутствовали во всех образцах, отобранных в разные сезоны года (рис. 1). Анализ сезонных изменений показал, что содержание дегидринов не оставалось постоянным и изменялось примерно в пять раз (от 0,04 до 0,15 усл. ед. интенсивности окрашивания) (рис. 2). Низкое содержание дегидринов выявлено в январе и третьей декаде апреля, когда в лесу еще лежал снег, но дневные температуры уже достигали +10о С. В сентябре содержание дегидринов в талломах было существенно выше. Высокие количества этих белков отмечены в конце июня. Этот период характеризовался сравнительно сухой и теплой погодой, когда температура воздуха в полуденные часы поднималась до +28... +30о С, при этом относительная влажность воздуха снижалась до 40 %. Анализ образцов, отобранных в разное время суток, показал максимум содержания дегидринов в полуденные часы (рис. 2), далее содержание обоих белков снижалось, к утру оно было в два раза ниже величин, характерных для полудня.
Содержание воды в лишайнике варьировало в пределах от 45 до 70 % сырой массы талломов (рис. 3). Наибольшая оводненность талломов была отмечена в октябре. В остальное вре- мя этот показатель составлял 50–60 % сырой массы образцов. Калориметрические определения показали, что температура фазового перехода «вода-лед» в холодный период (ноябрь–январь) составляла -9… -11о С, в теплое время была на 2–3о С выше. Заметное снижение температуры замерзания воды отмечали уже в сентябре. В зависимости от сезона и оводненности, фазовому переходу подвергалось от 25 до 45 % содержавшейся в талломах воды.
Нами выявлены сезонные изменения уровня бел-ков-дегидринов, оводненности и температуры замерзания воды в лишайнике L. pulmonaria . В талломах идентифицировано два дегидрина с мол. массой 40 и 43 кДа. По имеющимся в литературе сведениям, растительные дегидрины представлены белками с мол. массой от 15 до 150 кДа [14] и могут отличаться высоким внутривидовым
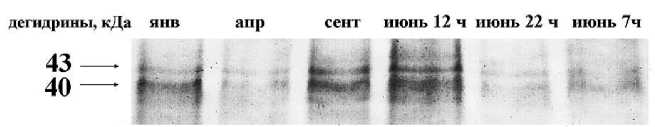
Рисунок 1. Сезонные и суточные изменения содержания дегидринов в талломах Lobaria pulmo-naria по результатам Western Blot. Приведено изображение типичной мембраны.
Условные обозначения. янв – январь; апр – апрель; сент – сентябрь. Слева указаны молекулярные
массы дегидринов.
Figure 1. Seasonal and daily changes in the content of dehydrins in the thalli of Lobaria pulmonaria according to the Western Blot. The figure illustrates the typical membrane.
Keys. янв –January; апр – April; сент– September. The molecular weights of the dehydrins are given on the left.
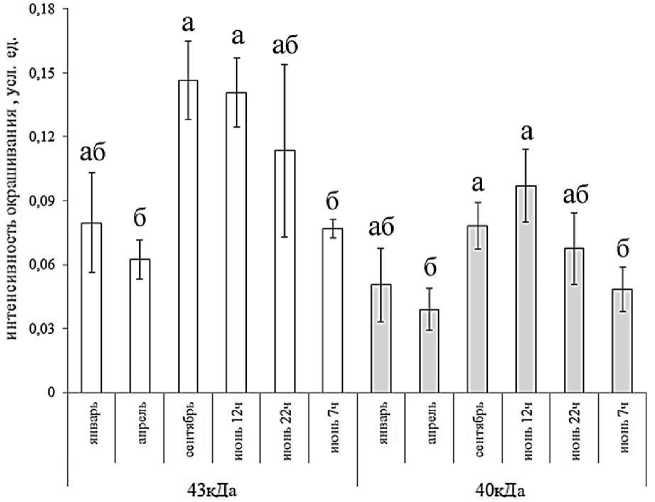
Рисунок 2. Сезонные и суточные изменения содержания дегидринов в талломах Lobaria pulmo-naria по результатам денситометрического измерения.
Примечание. Приведены средние значения и стандартные ошибки (n = 3). Разные буквы над столбцами обозначают статистически значимые различия в сезонной и суточной динамике. Оценку значимости различий проводили с помощью критерия Манна-Уитни в программе Statistica при p<0,05 отдельно для каждого из дегидринов.
Figure 2. Seasonal and daily changes in the content of dehydrins in the thalli of Lobaria pulmonaria according to the results of densitometric measurement.
Note. The mean values and standard errors (n = 3) are given. The different letters above the columns indicate significant differences between month and time of day. The significance of the differences was assessed using the Mann-Whitney U-test in the Statistica program at p<0.05 separately for each of the dehydrins.
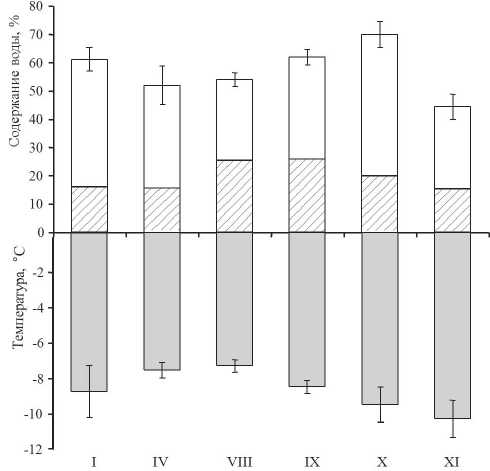
Рисунок 3. Сезонные изменения температуры фазового перехода «вода-лед», содержание воды и доля замерзшей воды (показано штриховкой) в талломах Lobaria pulmonaria .
Примечание. Римскими цифрами обозначены сроки (месяцы) отбора образцов.
Figure 3. Seasonal changes in the "water-ice" phase transition temperature, water content, and the fraction of frozen water (shown by hatching) in the thalli of Lobaria pulmonaria .
Note. Roman numerals indicate the sampling dates (months).
полиморфизмом [12, 22, 23].
Имеющиеся в литературе сведения о сезонной динамике дегидринов, в основном, касаются почек древесных и листьев хвойных растений [12, 13, 22, 24]. Характер сезонной динамики дегидринов в почках березы плосколистной и хвое сосны обыкновенной позволил авторам сделать заключение об участии этих белков в процессах, ассоциированных с формированием низкотемпературной устойчивости древесных растений [22]. Стабильно высокий уровень дегидринов сохранялся в течение всего периода с низкими отрицательными температурами, причем фракция низкомолекулярных дегидринов увеличивалась зимой [12, 13].
В талломах L. pulmonaria постоянно присутствовали два белка – дегидрина с мол. массой 43 и 40 кДа (рис. 1, 2), что свидетельствует об их конститутивном биосинтезе. L. pulmonaria является трехкомпонентным лишайником, представляющим собой симбиоз между грибом-аскоми-цетом (микобионтом), зеленой водорослью (фикобионтом) и цианобактерией (цианобионтом). Следует отметить, что дегидрин с мол. массой 40 кДа был обнаружен в лишайниковой водоросли Trebouxia erici [16]. В клетках фико-бионта L. pulmonaria водоросли Symbiochloris reticulate [25] были выявлены белки с типичными для дегидринов аминокислотными последовательностями [26]. В клетках линии цианобионта L. pulmonaria цианобактерии Nostoc дегидрин с мол. массой 40 кДа накапливался в ответ на осмотический стресс [27]. У грибов Alternaria brassicicola и Tuber borchii из отдела аскомицетов экспрессия генов нескольких дегидринов усиливалась при действии температурного, окислительного или осмотического стрессов
[28, 29]. Таким образом, весьма вероятно, что продуцентом дегидринов может быть любой из компонентов L. pulmo-naria , при этом продуцентом дегидрина с массой 40 кДа может быть цианобионт. Условия среды могут по-разному влиять на способность компонентов лишайника к биосинтезу дегидринов. Выявление вклада каждого из компонентов лишайника в накопление этих белков при разных условиях требует отдельного исследования.
Анализ сезонной динамики дегидринов в талломах лобарии выявил их пониженное содержание весной (конец апреля). Мы связываем это с благоприятными для активного метаболизма и роста лишайников условиями (достаточная влажность и умеренная температура). Согласно ранее опубликованным данным [6], интенсивность тепловыделения (показателя, отражающего общую метаболическую активность) весной была в 2,5 раза выше, чем летом.
Анализ образцов талломов, отобранных в сухой и теплый период (конец июня), выявил значительные суточные изменения уровня дегидринов. Наибольшее их содержание было отмечено в полуденные часы, когда талломы практически полностью теряли воду и высыхали. Учитывая способность дегидринов связывать воду, наши результаты могут свидетельствовать об участии этих белков в защите клеточных структур при дегидратации лишайников в теплый и сухой период.
Сведения о температуре замерзания воды в лишайниках неоднозначны. В работе [30] показано, что при охлаждении талломов Umbilicaria aprina до -20о С клеточная вода диффундировала из грибных и водорослевых клеток и кристаллизовалась на поверхности гиф в межклетниках медуллярного и альгального слоев. При нагревании таллома вода оттаивала и поглощалась клетками, которые снова становились тургесцентными. В талломах лишайников было обнаружено несколько фракций воды (слабо, сильно и очень сильно связанная), которые замерзали при разной температуре [31]. По нашим данным, полученным методом дифференциальной калориметрии, температура перехода «вода-лед» в талломах L. pulmonaria варьировала в пределах -7…-11о С, причем в теплый период года (весна – лето) она была выше, чем в холодный (поздняя осень – зима).
Большой интерес представляет показатель, характеризующий количество воды в талломе, которое может участвовать в кристаллизации. Эту величину можно оценить по площади, ограничиваемой кривой фазового перехода, «вода–лед». Расчеты показали, что замерзало от одной четверти до половины всей содержащейся в талломе воды. По-видимому, замерзанию подвергалась свободная вода, находящаяся в межклетниках, вакуолях, клеточной оболочке и, возможно, слабо связанная вода, гидратирующая коллоиды цитоплазмы. Вероятно, остальная часть воды была представлена фракциями сильно и очень сильно связанной воды. Такая вода может быть связана с гидрофильными радикалами макромолекул (белков, липидов) клеточных мембран, белками-ферментами и т. п. Согласно данным работы [32], при температурах -20° С и ниже всю способную к кристаллизации воду в мери- стемах почек хвойных древесных растений удерживают низкомолекулярные водорастворимые вещества, включающие в основном водорастворимые белки цитоплазмы, свободные аминокислоты и водорастворимые углеводы. Вероятно, что в формировании фракции сильно связанной воды участвуют также обладающие высокой гидрофильностью дегидрины. По данным [22], сезонные вариации дегидринов коррелировали с изменением содержания воды в хвое сосны обыкновенной.
Заключение
Итак, в лишайнике L. pulmonaria впервые идентифицировано два белка-дегидрина с мол. массой 40 и 43 кДа. Они присутствовали в талломах в течение всего года, но их содержание варьировало в зависимости от сезона. Наименьшее количество данных белков было обнаружено в наиболее благоприятный для роста талломов весенний период.
Показано увеличение содержания дегидринов летом в полуденные часы при десикации талломов. Уровень дегидринов сохранялся сравнительно высоким в осенне-зимний период. Температура кристаллизации воды в осенне-зимний период составляла в среднем –10о С и была на 2–3о С ниже, чем в более теплый весенне-летний период. Относительное содержание воды варьировало в пределах от 45 до 70 % сырой массы талломов. При снижении температуры фазовому переходу «вода – лед» подвергалось от 25 до 45 % всей содержащейся в талломах воды. Высказано предположение, что дегидрины могут участвовать в формировании фракции сильно связанной воды. По-видимому, существует связь между уровнем дегидринов, оводненностью и температурой замерзания воды в годичном цикле у талломов лишайника L. pulmo-naria . Однако требуется больше экспериментальных данных для подтверждения этого предположения.
