Сезонная динамика массовых родов фитопланктона в озере Байкал

Автор: Изместьева Л.Р., Мур М.В., Хэмптон С.Э., Зилов Е.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1 т.8, 2006 года.
Бесплатный доступ
Проанализированы изменения наступления годового пика численности 4-х наиболее массовых родов фитопланктона озера Байкал с 1951 по 1999 г. на основе долговременного ряда наблюдений. Несмотря на повышение температуры воздуха и более раннее освобождение ото льда южного бассейна в последнем столетии, сроки массового развития Aulacoseira, Cyclotella, Synedra и Ankistrodesmus (=Monoraphidium) не изменились. Сигналом к массовому развитию этих водорослей по всей вероятности, а большей степени служат условия освещенности, а не температура. За проанализированный 48-летний период средняя годовая численность Aulacoseira существенно снизилась, а численность водорослей других родов практически не изменилась. Снижение численности Aulacoseira может быть связано с изменением количества зимних осадков или ледовой обстановки, но эти вопросы требуют дальнейшей проработки.
Короткий адрес: https://sciup.org/148197799
IDR: 148197799 | УДК: 574.5(285.2)
Seasonal dynamics of common phytoplankton in lake Baikal
Changes in the seasonal dynamics of phytoplankton are potentially important, because they can generate a mismatch between the timing of peak productivity and that of consumers, thereby interrupting energy flow between trophic levels. Here, we use a long term data set for the phytoplankton community in the southern basin of Lake Baikal, Siberia to analyse for changes in the seasonal timing of peak abundance of the 4 most common genera during the period 1951-1999. Despite the development of warmer air temperatures and an earlier date of ice-off in the southern basin during the last century, the timing of peak abundance of Aulacoseira, Cyclotella, Synedra, and Ankistrodesmus (=Monoraphidium) did not change. These phytoplankton, especially the diatoms, may rely on light rather than temperature as a seasonal cue for initiating rapid population growth. During the 48-year period of analysis, mean annual abundance of Aulacoseira declined significantly, but the abundance of the other genera remained unchanged. The decline in Aulacoseira may be related to changes in winter precipitation and ice characteristics, but this remains to be examined.
Текст научной статьи Сезонная динамика массовых родов фитопланктона в озере Байкал
В последнее время все чаще появляются сообщения об изменении сезонной динами-ки и сроков массового развития фитопланк-тона, особенно в высоких широтах (см . на-пример [4, 21]). При этом указывается на кор-реляцию изменения сроков массового разви-тия водорослей с повышением температуры воды [7, 10] или ранним вскрытием водоемов, вызванных крупномасштабными климати-ческими аномалиями, такими , например, как Северо-Атлантическая Осцилляция [8, 19]. Изменения фенологии фитопланктонных ви-дов важны и в том плане, что они могут выз-вать разрыв между временем развития фито-планктона и консументов , прерывая или уменьшая, таким образом, поток энергии меж-ду трофическими уровнями [6]. Например, когда массовое развитие фитопланктона не совпало по времени с сезонным появлением личинок трески, поскольку фитопланктон и личинки трески реагируют на повышение температуры воды различно , восполнение за-пасов трески в Северном море существенно пострадало [7].
В озере Байкал многие виды фитопланк-тона характеризуются выраженной сезонно-стью пиков развития, приуроченных к опре-деленным коротким интервалам годового цикла [13]. Изменений фенологии фитоплан-ктона оз. Байкал можно ожидать, основыва-ясь на недавних сообщениях как о потепле-нии в его регионе [18], так и о тенденции бо-лее раннего вскрытия озера ото льда [15]. в настоящей работе мы использовали часть материалов базы данных о состоянии планк-тонного сообщества Байкала за 60 лет [1] для проверки изменений сроков массового раз-ВИТИЯ 4-х родов фитопланктона и проанали-зировали тренды среднегодовой численное-ти этих родов с 1951 по 1999 .
Методы
Пробы фитопланктона отбирались ежеме-сячно (обычно - чаще)с 1945 г. -ной пелагической станции в Южном Байка-ле, расположенной против пос. Большие Коты на расстоянии 2,7 км от берега над глу-биной 800 м (51°52 ’ 48 ” с.ш., 105°05 ’ 02 ” в.д.). Пробы фиксировались раствором Утермёлля, осаждались, просчет водорослей выполнял-ся под световым микроскопом. В данной ра-боте мы рассматриваем динамику 4-хнаибо-
: диатомовых Aulacoseira , Cyclotella и Synedra, зеленой Ankistrodesmus (= Monoraphidium ) в столбе воды 0-50 м.
Наличие изменений среднегодовой чис-ленности и времени массового развития мы определяли с помощью линейной регрессии с 1951 по 1999 г.
даты, на которые каждый год приходилось массовое развитие. Из анализа были исклю-чены 3 года (1992, 1993, 1995), поскольку тог- да отборы проб производились в течение менее чем 10 месяцев. Для каждой регрессии мы применяли тест Дурбина-Уотсона для проверки возможности влияния временной автокорреляции на получаемые результаты. Во всех случаях тест Дурбина-Уотсона пока-зал, что автокорреляция была несуществен-на (P > 0,18).
Результаты
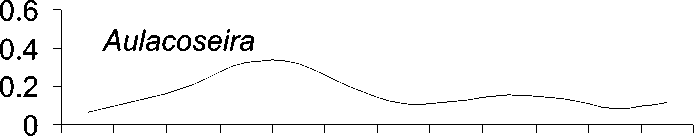
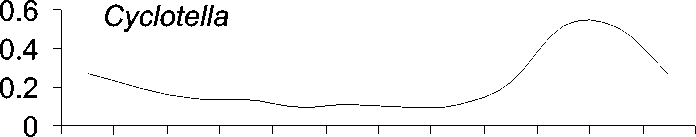
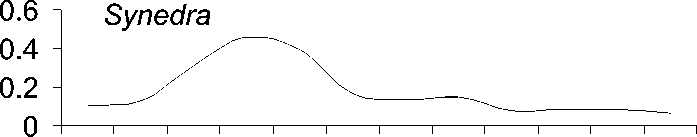
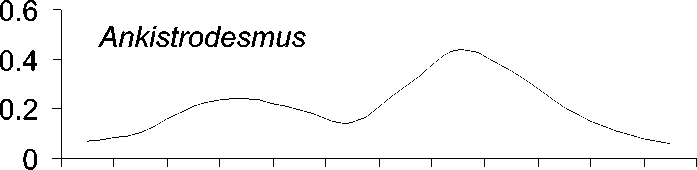
Исследованные роды фитопланктона ха-рактеризуются различным сезонным распре-делением (рис. 1).
1 2 3 4 5 6 7 8 9 10 11 12
мес:яцы
Рис. 1. Среднемноголетнее (с 1951 по 1999 г.) распределение численности водорослей (в% от годового максимума)в 0-50 м столбе воды по месяцам
Пики диатомей Aulacoseira и Synedra при-ходятся на подледный сезон, тогда как Cyclotella массово развивается поздней осе-нью или зимой перед покрытием Байкала льдом . Ankistrodesmus же , как правило , наи-более многочислен летом или ранней осе-НЬЮ .
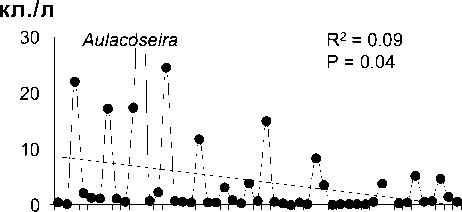
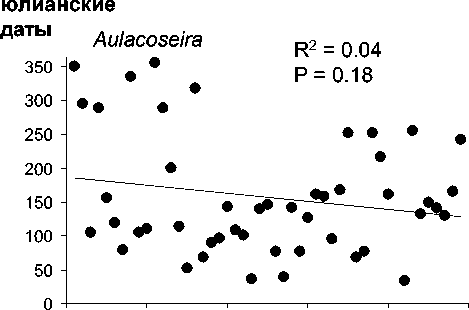
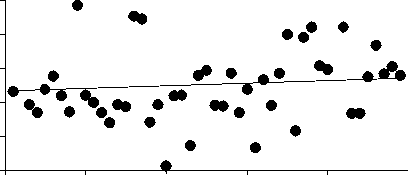
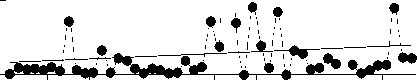
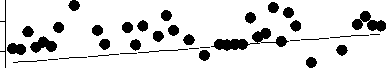
Среднегодовая численность Aulacoseira существенно (более чем на 70%) снизилась за рассматриваемый период, тогда как сред-негодовая численность 3-х других родов зна-чительно не изменилась (рис. 2). Снижение численности Aulacoseira никак не было свя-зано со сроками развития, сроки развития Aulacoseira, Cyclotella и Synedra устойчиво выдерживались все 48 лет (рис. 3).
Ankistrodesmus , напротив, проявляет неко-торую тенденцию к более позднему массо-вому развитию, хотя это изменение и не очень существенно (рис. 3).
Обсуждение
Сезонная динамика трех родов диатомо-вых водорослей (рис. 1) хорошо соответству-ет описанной в литературе . Виды родов Aulacoseira и Synedra обычно составляют ос-нову весеннего цветения подо льдом в мар-те -апреле [17], тогда как Cyclotella обычно массово развивается во время осенней гомо-термин в октябре-ноябре [16]. Осенние вспышки развития Cyclotella, вероятно, огра-ничиваются из-за ветрового перемешивания и выедания водорослей копеподой Epischura baicalensis Sars [17]. Пики численности Ankistrodesmus в августе и сентябре, возмож-но , обеспечивают пищей копеподиты и взрослую эпишуру до массового появления Cyclotella в октябре .
Значительное снижение численности Aulacoseira за проанализированный период (рис . 2) вызывает интерес, особенно в связи с тем , что эти водоросли часто доминируют в фитопланктоне Байкала по биомассе и вно-сят существенный вклад в первичную про-дукцию [14]. Более того, сообщаемое нами снижение численности водорослей этого рода, возможно, недооценивается. Надо сра-зу отметить, что годы , не включенные нами в анализ (1992, 1993 и 1995), также не были годами массового развития Aulacoseira («ме-лозирными годами») [5, 9]. Aulacoseira baicalensis (K.Meyer) Simonsen, эндемичный вид рода Aulacoseira, развивается подо льдом, но его рост угнетается, если глубина снега превышает 10 см . При таких условиях ослаб-ление света ограничивает как рост диатомей, так и процессы перемешивания воды, удер-живающие водоросли в толще воды [11, 16]. Количество снегопадов и толщина снегово-го покрова возросли в последние десятиле-тия на севере Евразии , возможно, в ответ на повышение зимних температур [12, 20]. Со-вместный анализ долговременных трендов толщины снегового покрова и льда с данны-ми долговременных наблюдений численно-сти A. baicalensis способен прояснить вопрос о влиянии изменения климата зимой на фун-кционирование байкальского пелагического сообщества.
Удивляет отсутствие сдвига пиков числен-ности 4-х родов водорослей в сторону более раннего развития их за 48 лет. -вало бы ожидать исходя из существенного повышения температуры воздуха в XX сто-летии [18] и удлинения сезона открытой воды на 16,1 дней за 137 лет [15]. В Северной Ев-ропе раннее вскрытие озер ото льда, связан-ное с Севере-Атлантической Осцилляцией, вызывает ранние вспышки цветения фито-планктона, в частности диатомового [2, 21]. В отличие от озера Байкал весеннее цвете-ние фитопланктона в Северной Европе про-исходит после схода льда, когда масса озер хо-рошо перемешивается, поддерживая фито-планктон в толще воды. В Байкале, однако, диатомовые водоросли родов Aulacoseira и Synedra развиваются главным образом подо льдом (в апреле-мае) и прекращают цветение по схождении льда в период ветрового пере-мешивания в июне (рис. 1). Следовательно, в оз . Байкал более раннее вскрытие льда мо-жет скорее прекращать весеннее цветение диатомей, а не сдвигать его на более ранние сроки . Автотрофный пикопланктон, однако, возмещает весеннее развитие диатомей в оз. Байкал в ходе короткого периода прямой тем-пературной стратификации в июле- августе [4], и эти водоросли могут отвечать на более раннее вскрытие озера сдвигом пика своего обилия, но этот вопрос требует дальнейшего изучения.
Представляется возможным, что для некоторых родов фитопланктона сигналом, инициирующим быстрый рост популяции, является , скорее, изменение освещенности, а не температуры.
Cyclotella
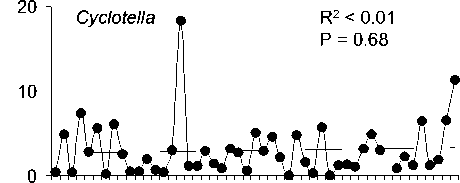
R2 < 0.01
P = 0.71
Synedra
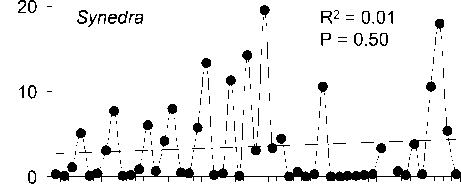
Ankistrodesmus
R2 < 0.01
P = 0.64
R2 = 0.04
P = 0.42
1951 1956 1961 1966 1971 1976 1981 1986 1991 1996
. 2.
4- .
0-50 1951 1999.
получены с помощью линейной регрессии
Ankistrodesmus
R2 = 0.07
P = 0.08
1950 1960 1970 1980 1990 2000
. 3.
0-50 1951 1999.
получены с помощью линейной регрессии (см. методы)
В Северной Атлантике сроки весенних вспышек развития диатомей остаются отно-сительно постоянными в течение последней полусотни лет, ччсмотря на существенное по вышение температуры воды [7]. Данные океанологических исследований подтверждают, что сохранение сроков весеннего развития диатомей связано именно с тем, что они кон- тролируются фотопериодизмом в большей степени, чем температурой [7]. Интересно, что 3 рода, рассматриваемых в настоящей статье, для которых не показано сдвигов сро-ков весеннего развития - диатомовые водо-росли. Единственный род, для которого по-казано некоторое изменение сроков макси-мального развития (p = 0,08), Ankistrodes-mus, зеленая водоросль.
Важно, что расхождение в сроках между развитием последовательных трофических уровней может произойти, если сроки раз-вития фитопланктона останутся прежними, а развитие зоопланктона сдвинется во вре-мени в ответ на повышение температуры воды [7]. В Северной Атлантике сроки цве-тения диатомей в течение последних 50 лет не изменились, тогда как многие группы зоо-планктона стали развиваться раньше, что и создало разрыв трофической цепи, вполне могущий быть ответственным за уменьшение трескового стада. Дальнейший анализ данных долговременных наблюдений планктона оз. Байкал необходим для проверки возможное-ти такого несоответствия в сроках развития
Список литературы Сезонная динамика массовых родов фитопланктона в озере Байкал
- База состояния планктона озера Байкал (База данных "ПЛАНКТОН", № 2005620028 Россия. Свидетельство/Л.Р Изместьева, Е.В. Пешкова; ГОУВПО Иркутский Государственный университет -№ 2004620262; заявл. 30.11.2004; опубл. 21.01.2005.
- Adrian R., Walz N., Hintze T., Hoeg S., Rusche R. Effects of ice duration on the plankton succession during spring in a shallow polymictic lake//Freshwater Biology. 1999. V. 41.
- Beaugrand G., Brander K.M., Lindley J.A., Souissi S., Reid P.C. Plankton effect on cod recruitment in the North Sea//Nature. 2003. V. 426.
- Belykh O.I., Sorokovikova E.G. Autotrophic picoplankton in Lake Baikal: Abundance, dynamics, and distribution//Aquatic Ecosystem Health and Management. 2003. Vol. 6.
- Bondarenko N.A. Floral shift in the phytoplankton of Lake Baikal, Siberia: Recent dominance of Nitzschia acicularis//Plankton Biology and Ecology. 1999. Vol. 46.
- Cushing D.H. Plankton production and year-class strength infish populations -an update of the matchmismatch hypothesis//Adv. Mar. Biol. 1990. Vol. 26.
- Edwards M., Richardson A.J. Impact of climate change on marine pelagic phenology and trophic mismatch//Nature. 2004. № 430.
- Gerten D., Adrian R. Climate-driven changes in spring plankton dynamics and the sensitivity of shallow polymictic lakes to the North Atlantic Oscillation//Limnology and Oceanography. 2000. Vol. 45.
- Granin N.G., Jewson D.H., Gnatovsky R.Yu. et al. Turbulent mixing under ice and the growth of diatoms in Lake Baikal//Verh. Intern. Verein. Limnol. 2000. Vol. 27.
- Hampton S.E. Increased niche differentiation between two Conochilus species over 33 years of climate change and food web alteration//Limnology and Oceanogr. 2005. Vol. 50.
- Jewson D.H., Granin N.G. How can present day studies of diatoms help in understanding past climatic change in Baikal?//Terra Nostra. 2000. Vol. 9.
- Kitaev L., Kislov A., Krenke A. et al. The snow cover characteristics of northern Eurasia and their relationship to climatic parameters//Boreal Environment Res. 2002. Vol. 7.
- Kozhova O.M. Phytoplankton of Lake Baikal: structural and functional characteristics//Arch. Hydrobiol. Beih. Ergebn. Limnol. 1987. Vol. 25.
- Kozhova O.M., Izmest'eva L.R. (Eds). Lake Baikal: Evolution and Biodiversity. 2nd edition. Leiden: Backhuys Publishers, 1998.
- Livingstone D.M. Ice break-up on southern Lake Baikal and its relationship to local and regional air temperatures in Siberia and to the North Atlantic Oscillation//Limnol. and Oceanogr. 1999. Vol. 44.
- Mackay A.W., Ryves D.B., Battarbee R.W. et al. 1000 years of climate variability in central Asia: assessing the evidence using Lake Baikal (Russia) diatom assemblages and the application of a diatom-inferred model of snow cover on the lake//Global and Planetary Change. 2005. Vol. 46.
- Popovskaya G.I. Ecological monitoring of phytoplankton in Lаke Baikal//Aquatic Ecosystem Health and Management. 2000. Vol. 3.
- Shimaraev M.N., Kuimova L.N., Sinyukovich V.N., Tsekhanovskii V.V. Manifestation of global climatic changes in Lake Baikal during the 20th century//Dokl. Earth Sci. 2002. Vol. 383A.
- Straile D., Adrian R. The North Atlantic Oscillation and plankton dynamics in two European lakestwo variations on a general theme//Global Change Biology. 2000. Vol. 6.
- Ye H. Decadal variability of Russian winter snow accumulation and its associations with Atlantic sea surface temperature anomalies//Intern. J. of Climatology. 2000. Vol. 20.
- Weyhenmeyer G.A., Blenckner T., Pettersson K. Changes of the plankton spring outburst related to the North Atlantic Oscillation//Limnology and Oceanogr. 1999. Vol. 44.
