Сезонные изменения зоопланктона в высокотрофных малых водоемах

Автор: Фефилова Е.Б., Кононова О.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные ресурсы
Статья в выпуске: 1-4 т.12, 2010 года.
Бесплатный доступ
Выявлены особенности сезонного развития зоопланктона в высокотрофных водоемах лесной зоны Северо-восточной части европейской России. Показано, что в гипертрофных прудах один раз за вегетационный сезон происходит смена доминирующего комплекса зоопланктона, так что динамика численности всего зоопланктлона имеет два пика. В них наблюдается более простая по сравнению с эвтрофными озерами трофическая структура планктонных животных. В разнотипных водоемах отмечали различные по направлению изменения видового разнообразия зоопланктона в течение вегетационного сезона.
Биологические сезоны, кривая динамики, трофность, водные экосистемы
Короткий адрес: https://sciup.org/148199121
IDR: 148199121 | УДК: 574.34:591.524.12
Zooplankton seasonal changes in hightrophic small reservoirs
Features of zooplankton seasonal development in hightrophic reservoirs of woody zone of the Northeast part of European Russia are revealed. It is shown, that in hypertrophic ponds once for a vegetative season there is change of zooplankton dominating complex so, that dynamics of numerosity of all zooplankton has two peaks. In them is observed more simple in comparison with eutrophic lakes trophic structure of planktonic animals. In polytypic reservoirs various in direction of change the zooplankton species diversity during a vegetative season was marked.
Текст научной статьи Сезонные изменения зоопланктона в высокотрофных малых водоемах
Материал и методы исследования.
Характеристика обследованных водоемов. Регион исследований расположен на водосборе р. Северная Двина, в бассейнах ее притоков первого (р. Вычегда) и второго (р. Сысо-ла) порядков. Изучение зоопланктона проводились на 2 прудах (1 и 2) и 4 озерах: Пезмог-ты, Важъэжва, Еля-ты, Куа-ты. Пруды имеют округлую форму, сходны по площади и глубине, их дно илисто-песчаное (табл. 1). Пойменные озера удлиненной формы, имеют постоянную связь с рекой (Вычегдой или Сысолой) посредством проток. В них преобладают илистые или илисто-песчаные донные субстраты. Летом прибрежье озер и прудов зарастает макрофитами.
Методы сбора и анализа данных. В прудах пробы зоопланктона отбирали с ноября 2005 по ноябрь 2006 гг.: в зимние месяцы (исключая декабрь), в марте, апреле – один раз в месяц, в остальные месяцы – подекадно. В периоды открытой воды из прибрежья и середины водоемов за раз отбирали четыре субпробы, из которых получали одну интегральную пробу. В зимнее время пробы отбирали из середины прудов. При этом получали за раз 3 интегральные пробы, объединяя для каждой из них субпробы с разных глубин. Полевые сборы материала из озер для исследования зоопланктона приходились только на вегетационные сезоны (с июня по сентябрь), из оз. Еля-ты – 3 лет (2003-2005 гг.), из остальных озер – 1-2 лет. В каждом из озер пробы отбирались ежемесячно с 9 станций, расположенных в прибрежье и на середине. Единожды в сезон из прудов и раз в месяц из озер получали пробы воды для ее химического анализа. Пробы зоопланктона отбирали принятыми в гидробиологии методами [8]. Результаты химического анализа водных проб были предоставлены нам экоаналитической лабораторией Института биологии Коми НЦ. Трофический тип водоемов определялся по коэффициенту трофии – Е [7]. Экологические группы зоопланктеров выделялись по способам их передвижения и питания по данным литературы [12] .
Таблица 1. Основные морфометрические и некоторые гидрохимические характеристики обследованных водоемов в вегетационный период
|
Водоем |
Площадь, км 2 |
Наибольшая глубина, м |
Общий азот, мг/л |
Общий фосфор, мг/л |
|
Пруд 1 |
~ 0,001 |
1,6 |
1,1 ± 1,1 |
0,10 ± 0,01 |
|
Пруд 2 |
~ 0,001 |
1,5 |
1,3 ± 1,0 |
нет данных |
|
Оз. Пезмог-ты |
1,18 |
3,5 |
1,0 ± 0,4 |
0,14 ± 0,05 |
|
Оз. Важъэжва |
0,6 |
4,0 |
0,6 ± 0,1 |
0,15 ± 0,04 |
|
Оз. Еля-ты |
0,28 |
6,0 |
1,6 ± 0,9 |
0,05 ± 0,01 |
|
Оз. Куа-ты |
0,12 |
3,5 |
1,1 ± 0,2 |
0,12 ± 0,05 |
Результаты исследования.
Условия обитания гидробионтов. Ход температуры воды в обследованных прудах в 2006 г. сопровождался максимумами в конце июня (25-26°С). Полное освобождение этих водоемов ото льда пришлось на 26-28 апреля, а образование устойчивого ледяного покрова – на 3 октября. Наименьшая концентрация растворенного кислорода в прудах (2 мг/л) была зарегистрирована в марте, когда толщина льда была наибольшей (68 см). В дальнейшем динамика этого показателя была различной для прудов 1 и 2. В обследованных озерах наибольший за вегетационный сезон прогрев воды (25-28°С) приходился на июль.
По уровню жесткости и рН воды прудов и большинства озер относятся к мягким и нейтрально-щелочным. По ионному составу они однотипны, относятся к гидрокарбонатно-кальциевым. Содержание биогенных элементов (табл. 1) колебалось в течение сезона в пределах, соответствующих для пруда 2 уровню слабой эвтрофикации (содержание общего азота), для оз. Еля-ты – средней эвтрофикации и для остальных водоемов – высокому уровню загрязнения биогенами [18].
Сезонные изменения зоопланктона. Зоопланктон обследованных высокотрофных водоемов за весь период наблюдений был представлен 240 видами и формами. Планктонная фауна озер была богаче по числу таксонов. В прудах за весь год было зарегистрировано 68 (в пруду 2) и 70 (в пруду 1) видов. В озерах число видов, встреченных в течение 1-3 вегетационных сезонов, варьировало от 100 (в оз. Важъэж-ва) до 120 (в оз. Еля-ты). Очевидно, что видовое богатство планктонной фауны в обследованных водоемах зависело от их размера и типа. По числу видов в прудах преобладали коловратки (39-40 видов), в озерах – ракообразные (52-63). Коэффициенты Съеренсена, рассчитанные для определения сходства видового состава зоопланктона в водоемах в летние месяцы, составили: для прудов – 0,64, для озер – 0,70-0,73, для прудов и озер – 0,43-0,51. Типологически сходные водоемы обладали высоким сходством видового состава сообществ, что позволило анализировать их ниже совместно.
Первоначально для прудов по развитию животного планктона были выделены основные этапы их сезонной сукцессии – биологические сезоны, которые по срокам не совпадали с календарными. Основными критериями, по которым эти сезоны были выделены, являлись сроки появления в планктоне тех или иных видов, достижение ими наибольшей числен-ности, а также сроки их размножения. В зимний период в планктоне присутствовало небольшое количество круглогодичных форм: Lecane luna (Müller), Bdelloida n.det., Macrocyclops albidus (Jurine), Eucyclops macruroides (Lilljeborg), Cyclops strenuus Fischer, C. kolensis Lilljeborg, C. vicinus Uljan, M. leuckarti (Claus). Ракообразные не размножались (самки с яйцами не встречались), все теплолюбивые, летние виды, равно как весенние, находились в состоянии угнетения и если и встречались в планктоне, то в состоянии «активной» диапаузы: M. leuckarti – на стадии копеподита, остальные виды циклопид (холодолюбивые) – на взрослой стадии. В течение биологической весны в водоемах размножались холодолюбивые планктонные циклопиды, эти же формы достигали своего наибольшего разнообразия. К началу биологического лета цикл их размножения заканчивался, и они либо выпадали из состава зоопланктона, либо существенно снижали численность. Их повторное за сезон появление в планктоне и размножение наблюдали с наступлением сезона биологической осени (рис. 1). К «весеннее-осенним» прудовым видам были отнесены холодолюбивые: коловратка Euchlanis incisa Carlin и веслоногие раки – C. strenuus , C. kolensis , C. insignis Claus, Megacyclops gigas (Claus), Diacyclops bicuspidatus (Claus), D. bisetosus (Rehberg). Оптимальные для этих видов температуры составляли 3,6-13,6°С. В июне в планктоне начинали появляться теплолюбивые виды коловраток и ракообразных. Только в конце мая и в июне в прудах постоянно присутствовала коловратка Keratella testudo (Ehrenberg). Diaphanosoma brachyurum (Lievin), Daphnia longispina O.F.Müller появились в водоемах в начале лета. В этом же сезоне они образовали пик численности и вошли в состав доми-нантов в зоопланктоне пруда 1. В пруду 2 в этот период по численности преобладала K. quadrata (Müller).
Наступление второй части биологического лета было связано с развитием макрофитов и сопровождалось сменой доминирующих в зоопланктоне таксонов, появлением теплолюбивых, фитофильных форм, не встречающихся до сих пор и ставших обычными в сообществах к концу лета. В июле-сентябре в планктоне прудов массово развивались и доминировали молодь циклопид родов Thermocyclops, Mesocyclops, а также коловраток рода Polyarthra и ветвистоусых семейства Chydoridae (рис. 1). Последние преобладали в пруду 2, зеркало которого почти полностью покрывали заросли макрофитов. Только во второй половине лета в зоопланктоне прудов присутствовали половозрелые Paracyclops affinis (Sars) и Ectocyclops phaleratus (Koch).
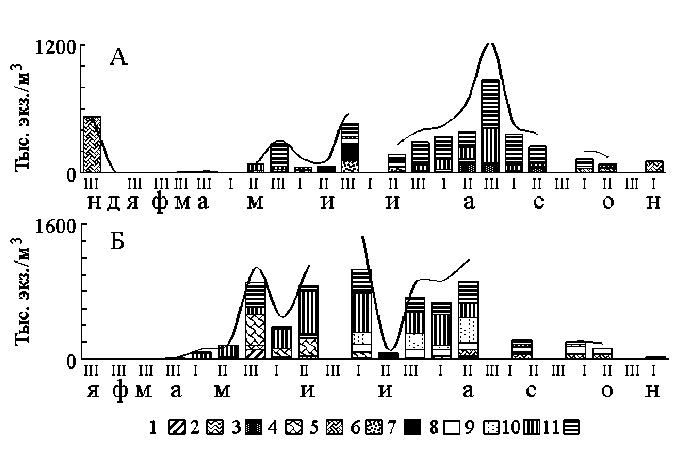
Рис. 1. Сезонная динамика численности зоопланктона в обследованных прудах 1 (А) и 2 (Б), ноябрь 2005 г. – ноябрь 2006 г.
1 – Polyarthra longiremis , 2 – P. vulgaris , 3 – P. sp ., 4 – Keratella quadrata , 5 – Bdelloida n.det., 6 – Diaphanosoma brachiu-rum , 7 – Daphnia longispina , 8 – Chydorus sphaericus , 9 – Alonella excisa , 10 – Nauplius, 11 – Copepodit. По оси X: декады (арабскими цифрами) и месяцы, сплошной линией показана динамика общей численности зоопланктона
Наивысшее видовое разнообразие животного планктона в обследованных прудах наблюдали в период наибольшего прогрева воды, начиная со второй части лета показатели разнообразия зоопланктона снижались (табл. 2). Трофическая структура доминирующих планктонных комплексов в гипертрофных водоемах изменялась в течение года через кратковременное увеличение роли первичных фильтраторов в июне к преобладанию вторичных фильтраторов к концу вегетационного сезона. Смешанная группа ювенильных форм Cyclopoida была многочисленна во все сезоны. Роль хищников в планктонных сообществах была незначительна (табл. 2).
В озерах в течение лета также происходила смена доминирующих комплексов. Холодолюбивые Polyarthra dolichoptera Idelson, C. kolensis единично присутствовали в планктоне в июне и сентябре и не встречались в остальные месяцы. Ко второй половине лета в прибрежных биотопах озер разнообразие зоопланктона повышалось за счет фитофильных или придонных коловраток и рачков (рода Trichocerca, Pleuroxus, Graptoleberis, Camptocercus, Alona). Доминирующие в сообществах рачок Polyphemus pediculus (Linne) (в прибрежье оз. Еля-ты) и коловратка A. priodonta (в середине озер Еля-ты, Куа-ты и Важъэжва), сезонная динамика популяций которых была изучена в обследованных озерах [4], характеризовались одним полным за сезон циклом развития. P. pediculus имел один пик численности в июне, в августе развитие популяции вида завершалось (рис. 2). A. priodonta достигала высокой численности популяции дважды: в июне и августе-сентябре (рис. 2), но только осенний пик завершался половым размножением этой коловратки.
Экологическая (трофическая) структура планктонных сообществ в пойменных озерах была хорошо развита и в целом сохранялась в течение всего периода исследований. Биомасса зоопланктона от начала к середине-концу вегетационного сезона снижалась в связи с повышением в сообществе роли мелких кладоцер относительно крупных. В том же временном направлении число видов зоопланктеров увеличивалось, а видовое разнообразие по индексу Симпсона, наоборот, уменьшалось (табл. 2).
Таблица 2. Основные характеристики зоопланктона обследованных водоемов в разные сезоны
S в bi
В в в в в в в
§ к 9
в
W в в к к
В
к В к с
захвату
В в пруды
|
весна |
IV, V |
10,5 |
54 |
126±46 |
1,6±0,5 |
0,51±0,06 |
1,09±0,11 |
Cop |
Cop |
⊗ ⟡ |
|
начало лета |
VI |
6,0 |
56 |
580±179 |
5,4±3,3 |
0,86±0,02 |
2,30±0,13 |
R, Cop, Cl |
Cop, Cl |
⊗ ⊕ ⟡ |
|
лето |
VII-IX |
6,2 |
92 |
665±118 |
4,2±0,5 |
0,77±0,03 |
2,02±0,09 |
Cop |
Cop |
⟡ |
|
осень |
X, XI |
7,0 |
51 |
154±24 |
2,4±0,4 |
0,72±0,06 |
1,70±0,23 |
R, Cop, Cl |
Cop, Cl |
∇ ⟡ |
|
зима |
XII-III |
– |
8 |
2±1 |
0,1±0,0 |
0,25±0,14 |
0,41±0,24 |
R, Cop |
Cop |
– |
|
озера |
||||||||||
|
начало лета |
VI |
2,0 |
114 |
81±12 |
4,5±1,6 |
0,31±0,01 |
2,44±0,06 |
R, Cop, Cl |
R, Cop, Cl |
► ⊗ ⟡ |
|
лето |
VII-IX |
2,0 |
157 |
111±27 |
1,1±0,2 |
0,23±0,01 |
2,93±0,04 |
R, Cop, Cl |
R, Cop, Cl |
► ⊗ ⊕ ⟡ |
Примечание : «R» - Rotatoria, «Cop» - Copepoda, «Cl» - Cladocera, « ⊗ » - плавающие вертикаторы, « ⊕ » - плавающие фильтра-торы, « ∇ » - плавающие, ползающие вторичные фильтраторы, «►» - плавающие хищники, «⟡» смешанные
Обсуждение результатов. Сезонная динамика зоопланктона в водных экосистемах умеренного пояса связана с размером водоема, степенью его зарастаемости и трофическим статусом [16, 20, 23]. Концентрации общего азота и фосфора в обследованных водоемах изменялись по сезонам и варьировали в диапазоне, соответствующем высокой трофности. Состав планктонных животных-индикаторов (отраженный в индексе Е, табл. 2) в течение всего года или вегетационного сезона соответствовал гипер-трофному уровню состояния экосистем прудов и эвтрофному – озер, что характеризует его как надежный показатель трофического статуса малых водоемов, первоначально для которых он и применялся [7]. Обследованные пруды оказались более продуктивными и менее экологически благополучными (по гидрохимическим показателям, по разнообразию зоопланктона и т.д.) по сравнению с озерами, что соответствует закономерности увеличения трофического статуса водоемов умеренной зоны от более глубоководных к более мелким [1, 15, 23]. Небольшое количество видов планктонных беспозвоночных в обследованных прудах и чрезвычайно низкое количественное развитие зоопланктона в них в холодный период обусловливались малыми размерами водоемов. В крупных покрытых льдом озерах зимние сообщества богаче и представлены большим числом форм, в том числе кладоцерами [9, 10], которые отсутствовали в прудах.
По имеющимся обобщенным сведениям [1], только в высокотрофных водоемах наблюдается несколько пиков численности за вегетационный сезон, которые можно описать одновершинной интегральной кривой с максимумом в летний период. В этих водоемах обычно преобладают мелкие формы планктонных животных (ветвистоусые родов Bosmina, Chydorus и коловратки), чья жизненная стратегия и тип размножения определяют такую динамику общей численности. Вместе с тем, для некоторых эвтрофных водоемов описан один пик численности зоопланктона, сложенного, в основном, веслоногими раками [17, 21]. В исследованных гипертрофных прудах описать динамику численности кривой с одной за вегетационный сезон вершиной представляется нам затруднительным. Два пика численности зоопланктона, по одному на каждый из летних биологических сезонов - «начало лета» и собственно «лето» -прослеживаются в прудах, где некоторые коловратки или кладоцеры повышали численность в оба летних сезона на фоне количественного превалирования молоди циклопид. Сходная сезонная динамика зоопланктона наблюдалась в водных экосистемах на юге Канады [14].
В малых гипертрофных водоемах смена состава доминирующего комплекса может быть обусловлена снижением в их водах концентрации растворенного кислорода, что характерно для водных объектов такого типа. Известно [3], что рачки рода Daphnia проявляют большую чувствительность к понижению этого показателя, которая проявляется в снижении продуктивности этих животных за счет уменьшения скорости ювенильного роста и снижении их способности к конкуренции. Возможно, подобные реакции на изменение условий среды в летнее время в высокотрофных водоемах присущи и другим планктонным гидробионтам, чья экология менее изучена.
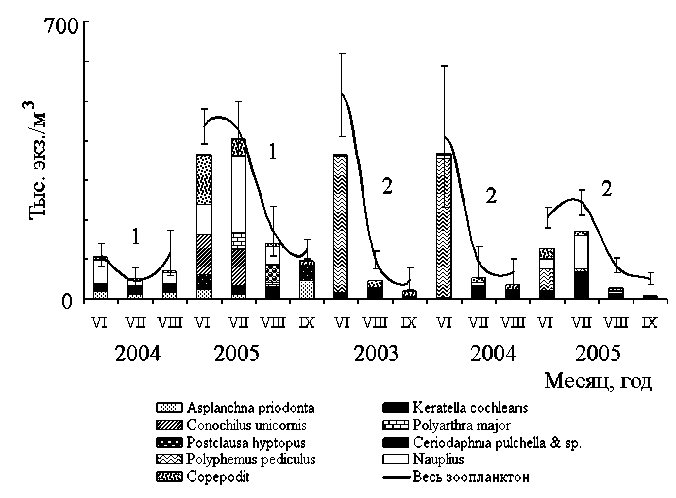
Рис. 2. Динамика численности зоопланктона и доминирующих видов и форм в оз. Еля-ты: 1 – в середине, 2 – в прибрежье
Трофическая структура планктонных сообществ обследованных прудов оказалась более простой по сравнению с таковой в озерах. В последних летний состав доминирующих экологических групп обнаруживал сходство с их составом в мезотрофных условиях [14]. Преобладание форм со смешанным типом питания и способных на разных стадиях онтогенеза потреблять весь спектр наличных пищевых ресурсов, и небольшая роль хищников в прудах также идентифицирует высокий трофический статус этих экосистем [2, 15]. Повышение в планктоне исследованных гипертрофных водоемов доли организмов, добывающих пищу с поверхности субстрата, только осенью связано с накоплением в них органического вещества [2, 15] и согласуется с данными о сезонном повышении трофности. Присутствие в планктоне теплолюбивых форм лишь в период с июня по сентябрь, моноциклия доминирующих видов, которые в более южных широтах имеют 2 и более цикла развития [4], можно отнести к региональным особенностям сезонной динамики зоопланктона в обследованных водоемах. Они определяют 2 пика интегральной кривой, наблюдаемых и в других северных регионах лесной зоны [14], сближенных во времени благодаря небольшой продолжительности тепловодного периода, а также быстрому прогреванию водных масс после его наступления в условиях мелководности.
Выводы:
-
1. В результате проведенных исследований показано, что сезонная динамика зоопланктона в мелких лесных водоемах Северо-Востока европейской России зависела от их трофического статуса и климатических особенностей региона. В течение всего года в прудах и, по крайней ме-
- ре, вегетационного сезона – в озерах сохранялись стабильно высокие значения коэффициента трофности. В обследованных прудах и пойменных озерах в течение вегетационного сезона наблюдалось 2 пика развития планктонных сообществ, различающихся по таксономическому и экологическому составу доминирующих комплексов. Для гипертрофных прудов такой ход сезонной динамики животного планктона, по-видимому, был специфичен. В гипертрофных водоемах наблюдалась более простая, чем в эвтрофных, структура планктонных сообществ, меньшее видовое богатство и разнообразие.
-
2. Показано, что наибольшее число видов присутствовало в планктоне прудов во второй половине биологического лета, максимальные биомассы зоопланктона, видовое и экологическое разнообразие приходились на начало биологического лета. В эвтрофных озерах зоопланктон был более разнообразен как в видовом отношении, так и по числу доминирующих экологических групп во второй половине летнего сезона.
Работа выполнена при поддержке гранта РФФИ № 07-04-00006-A и проекта, выполняемого совместно организациями УрО, СО и ДВО РАН - № 09-С-4-1017.
Список литературы Сезонные изменения зоопланктона в высокотрофных малых водоемах
- Андроникова, И.Н. Структурно-функциональная организация зоопланктона озерных экосистем. -СПб: Наука, 1996. -189 с.
- Деревенская,О.Ю. Структура пищевых сетей в сообществах зоопланктона разнотипных озер Волжско-Камского государственного природного биосферного заповедника/О.Ю. Деревенская, Е.Н. Унковская//Биология внутренних вод. -2007. -№2. -С. 55-61.
- Дубовская, О.П. Не связанная с хищниками смертность планктонных ракообразных, ее возможные причины (обзор литературы)//Журнал общей биологии. -2009. -Т.70, №2. -С. 168-192.
- Кононова, О.Н. Структура и динамика зоопланктона водоемов бассейна среднего течения реки Вычегда. Автореф. дисс. на соискание уч. степени канд. биол. наук. Сыктывкар, Институт биологии Коми научного центра Уральского отделения РАН, 2009. -19 с.
- Лазарева, В.И. Зоопланктон малых озер Южной Карелии при различном уровне pH и гумификации//Экология. -1996. -№1. -С. 33-39.
- Моисеенко, Т.И. Зональные особенности формирования химического состава вод малых озер на территории европейской части России/Т.И. Моисеенко, Н.А. Гашкина, Л.П. Кудрявцева и др.//Водные ресурсы. -2006. -Т. 33, № 2. -С. 163-180.
- Мяэметс, А.Х. Изменения зоопланктона//Антропогенное воздействие на малые озера. -Л.: Наука, 1980. -С. 54-64.
- Ривьер, И.К. Зоопланктон и нейстон//Методика изучения биогеоценозов внутренних водоемов. -М.: Наука, 1975. -С. 138-157.
- Ривьер, И.К. Состав и экология зимних зоопланктонных сообществ. -Л.: Наука, 1986. -160 c.
- Столбунова, В.Н. Зоопланктон озера Плещеево. -М.: Наука, 2006. -152 с.
- Сярки, М.Т. Сезонная динамика зоопланктона Онежского озера (влияние температурного и антропогенного факторов)//Экологическое состояние континентальных водоемов северных территорий. -СПб: Наука, ВВМ, 2005. -С. 215-220.
- Barnett, A.J. Functional diversity of crustacean zooplankton communities: towards a trait-based classification/A.J. Barnett, K. Finlay, B.E. Beisner//Freshwater Biology. -2007. -V. 52. -P. 796-813.
- Bledzki, L.A. Pelagic zooplankton of 11 lakes in Tuchola Forasts and 14 lakes in Brodnica lakeland/L.A. Bledzki, L. Bittel, D. Bronisz-Wolnomiejska//Acta Universitatis Nicolai Copernici. Limnological Papers. -1992. -№17. -P. 3-29.
- Hann, B.J. Littoral microcrustaceans (Cladocera, Copepoda) in a prairie coastal wetland: seasonal abundance and community structure/B.J. Hann, L. Zrum//Hydrobiologia. -1997. -V. 357. -P. 37-52.
- Hanson, P.C. Small lakes dominate a random sample of regional lake characteristics/P.C. Hanson, S.R. Carpenter, J.A. Cardille et al.//Freshwater Biology. -2007. -V. 52. -P. 814-822.
- Karabin, A. Eutrophication processes in a shallow, macrophyte-dominated lake -factors influencing zooplankton structure and density in Lake Łuknajno (Poland)/A. Karabin, J. Ejsmont-Karabin, R. Kornatowska//Hydrobiologia. -1997. -V. 342/343. -P. 401-409.
- Mayer, J. Seasonal succession and trophic relations between phytoplankton, zooplankton, ciliate and bacteria in a hypertrophic shallow lake in Vienna, Austria/J. Mayer, M. Dokulil, M. Salbrechter et al.//Hydrobiologia. -1997. -V. 342/343. -P. 165-174.
- Moss, B. The determination of ecological status in shallow lakes -a tested system (ECOFRAME) for implementation of the European Water Framework Directive/B. Moss, D. Stephen, C. Alvarez et al.//Aquatic Conserv: Mar. Freshw. Ecosyst. -2003. -V. 13. -P. 507-549.
- Nõges, P. Factors controlling hydrochemical and trophic state variables in 86 shallow lakes in Europe/P. Nõges, T. Nõges, L. Tuvikene et al.//Hydrobiologia. -2003. -V. 506-509. -P. 51-58.
- Nurminen, L.K.L. A diurnal study on the distribution of filter feeding zooplankton: Effect of emergent macrophytes, pH and lake trophy/L.K.L. Nurminen, J.A. Horppila//Aquat. Sci. -2002. -V. 64. -P. 198-206.
- Pociecha, A. Dynamics of phyto-and zooplankton in the submountane dam reservoirs with different trophic status/A. Pociecha, E. Wilk-Wozniak//Limnological Review. -2005. -V.5. -P. 215-221.
- Rautio, M. Benthic and pelagic food resources for zooplankton in shallow high-latitude lakes and ponds/M. Rautio, W.F. Vincent//Freshwater Biology. -2006. -V. 51. -P. 1038-1052.
- Søndergaard, M. Pond or lake: does it make any difference?/M. Søndergaard, E. Jeppesen, J.P. Jensen//Arch. Hydrobiol. -2005. -V. 162, № 2. -P. 143-165.
