Сезонный рост побегов представителей рода Аcer (Аceraceae) в условиях интродукции
 в условиях интродукции")
Автор: Кищенко Иван Тарасович, Потапова Марина Николаевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 2 (92), 2008 года.
Бесплатный доступ
Исследования проводились в подзоне средней тайги (Южная Карелия). Объектами исследований служили представители шести интродуцированных видов рода Acer: Acer ginnala Maxim., A. semenovii Regel. et Herd., A. negundo L., A. tataricum L., A. platanoides L. и A. pseudoplatanus L. В результате проведенных исследований установлено, что особенности роста побегов и листьев обусловлены биологическими свойствами вида.
Рост, побеги, виды клена, интродукция
Короткий адрес: https://sciup.org/14749400
IDR: 14749400
Текст научной статьи Сезонный рост побегов представителей рода Аcer (Аceraceae) в условиях интродукции
Усиливающееся загрязнение окружающей среды все настоятельнее требует увеличения объема озеленительных работ. Большинство аборигенных видов древесных растений таежной зоны России плохо переносят прогрессирующее загрязнение окружающей среды. Между тем многие виды лиственных древесных растений, в т. ч. и рода Acer (из других географических районов) устойчивы к загазованно сти и задымлению воздуха. По данным К. К. Калуцкого и Н. А. Болотова [1], хорошо подобранные к новым экологиче ским условиям виды древесных растений при введении в культуру часто значительно перегоняют по продуктивности местные виды. В связи с этим предполагается их интродукция и ее оценка.
Один из важнейших показателей интродукции – степень соответствия ритмики роста и развития растения динамике экологических факторов [2]. Именно сезонный ритм роста является интегральным показателем, характеризующим адаптацию растений к условиям среды и соответствие по следних биологии вида [3, 4].
В отечественной литературе выяснению особенностей сезонного роста вегетативных органов лиственных древесных растений уделено сравнительно мало внимания [5, 6, 7]. Цель нашей работы – изучение в этом аспекте интродуцентов рода Acer в таежной зоне России.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводили в 2001–2002 гг. в ботаническом саду Петрозаводского государственного университета, расположенного на северном берегу Петрозаводской губы Онежского озера (подзона средней тайги). Объектами исследований служили 6 видов рода Acer : клен приречный A. ginnala Maxim., клен Семенова A. semenovii Rеgеl. et Herd., клен ясенелистный A. negundo L., клен татарский A. tataricum L., клен остролистный A. platanoides L. и клен ложноплатановый A. pseudoplatanus L.* (табл. 1).
Таблица 1
Характеристика объектов исследований
|
Вид растения |
Место происхождения саженцев, город |
Возраст, лет |
|
Acer ginnala Maxim. |
Красное село |
46 |
|
A. negundo L. |
С-Петербург |
51 |
|
A. platanoides L. |
Красное село |
61 |
|
A. pseudoplatanus L. |
Красное село |
51 |
|
A. semenovii Regel. et Herd. |
Красное село |
46 |
|
A. tataricum L. |
Красное село |
61 |
*Примечание: по С. К. Черепанову [8].
С целью изучения роста побегов текущего года измеряли длину осевых стеблей второго порядка ветвления с южной части кроны на высоте 1–1.5 м с момента набухания почек до заложения зимующих почек, площадь листьев – с момента их обособления до полного прекращения роста через каждые 2–3 дня. Объем выборки по каждому объекту – 25 побегов и листьев. Величину суточного прироста определяли как разницу в длине изучаемых органов между последующим и предшествующим наблюдениями, деленную на число суток этого периода [9, 10].
Климатические данные были получены от Сулажгорской метеостанции (Карельская гидрометобсерватория), расположенной в 3 км юго-западнее Ботанического сада. Все выборки проверены на закон нормального распределения. Коэффициенты корреляции и различия между средними величинами оценены на достоверность. Из полученных элементарных статистик, в частности, следует, что показатель точности опыта довольно высок (4–5%), а коэффициент вариации невелик (13–17%).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Проведенные исследования позволили установить, что сроки начала роста побегов изучаемых видов Acer могут варьировать по годам в пределах двух недель и приходятся на первую половину июня (рис. 1). При этом различия между видами не превышают 3–4 сут (табл. 2). Изменчивость такого рода отмечал и Н. В. Шкутко [11].
Сроки окончания роста побегов варьируют по годам также в пределах двух недель. Время наступления этой фенофазы у разных видов Acer отличается не более чем на 3–7 сут и наблюдается во второй половине июля. По мнению Н. В. Шкутко [11], степень адаптации интроду-центов к новым климатическим условиям тем выше, чем менее изменчивы сроки начала и окончания их фенофаз. По нашим данным, наименьшая вариабельность сроков прекращения роста побегов (в пределах 3 сут) характерна для A. ginnala , A. negundo и A. platanoides.
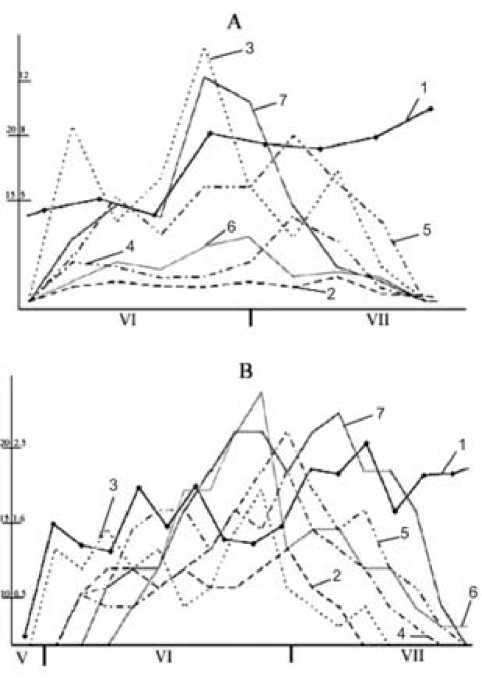
Рис. 1. Сезонная динамика температуры воздуха и суточного прироста побегов различных видов Acer
Годы: А – 2001, В – 2002. 1 - температура воздуха, 2 - Acer plata-noides; 3 - A. negundo; 4 - A. ginnala; 5 - A. pseudoplatanus;
6 - A. tataricum; 7 - A. semenovii. По вертикальным осям: слева - температура воздуха, ºС; справа – суточный прирост, мм
Значительные погодичные изменения в сроках начала и окончания роста побегов соответственно отражаются на продолжительности их формирования. В зависимости от вида растения она варьирует от 34 до 56 сут (табл. 3).
Установлено, что время кульминации прироста побегов Acer весьма существенно изменяется по годам. Раньше всех эта фаза наступает у A. negundo (20 VI–28 VI), а позднее – у A. se-menovii (25 VI–24 VII). Таким образом, различия между видами по этому показателю достигают почти месяца. Величина максимального суточного прироста побегов у изученных видов клена также значительно различается. Его наибольшая величина (до 11–13 мм/сут) обнаружена у A. ne-gundo и A. semenovii . У других изучаемых видов этот показатель меньше в 2–10 раз. Следует подчеркнуть, что погодичная изменчивость величины максимального прироста у побегов может достигать 50–400% (табл. 3).
Таблица 2
Температурный режим в период роста побегов (над чертой) и листьев (под чертой) у различных видов Acer
|
Вид |
ж к ч 2 ч ю cd К 3 ч |
Начало роста |
Кульминация прироста |
Окончание роста |
||||||
|
cd Й |
cd 3 ^ ® g о m и 1 |
я и я R н ° а с 2 Я V 2 * >1 я U Я |
cd Й |
cf 3 ® g о m и 1 |
л и s R н ° а с 2 U Я |
cd Й |
cd 3 ® g о m ||у и | |
я и Я R н ° а с 2 >1 я U Я |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
|
Acer ginnala |
2001 |
12 VI |
10.8 |
550 |
3–6 VII |
20.5 |
889 |
19 VII |
23.6 |
1150 |
|
01 VI |
8.0 |
363 |
05–08 VI |
14.1 |
453 |
16 VII |
23.8 |
1083 |
||
|
2002 |
01 VI |
20.2 |
300 |
28 VI–02 VII |
15.0 |
770 |
22 VII |
17.8 |
1158 |
|
|
24 V |
2.5 |
215 |
12–16 VI |
13.7 |
526 |
18 VII |
16.6 |
1081 |
||
|
A. negundo |
2001 |
12 VI |
10.8 |
550 |
25–28 VI |
12.3 |
725 |
16 VII |
23.8 |
1083 |
|
01 VI |
8.0 |
363 |
08–12 VI |
12.8 |
503 |
16 VII |
23.8 |
1083 |
||
|
2002 |
28 V |
7.5 |
239 |
24–28 VI |
14.9 |
711 |
14 VII |
18.5 |
1015 |
|
|
24 V |
2.5 |
363 |
12 –16 VI |
13.7 |
526 |
18 VII |
16.6 |
1081 |
||
|
A. platanoides |
2001 |
16 VI |
16.4 |
602 |
7–10 VII |
18.2 |
961 |
19 VII |
23.6 |
1150 |
|
01 VI |
8.0 |
363 |
03–07 VII |
19.6 |
905 |
16 VII |
23.8 |
1083 |
||
|
2002 |
01 VI |
20.2 |
300 |
28 VI–02 VII |
15.0 |
770 |
14 VII |
18.5 |
1015 |
|
|
24 V |
2.5 |
215 |
16–20 VI |
19.0 |
600 |
10 VII |
21.1 |
928 |
||
|
A. pseudoplatanus |
2001 |
12 VI |
10.8 |
550 |
3–6 VII |
20.5 |
889 |
16 VII |
23.8 |
1083 |
|
01 VI |
8.0 |
363 |
01–05 VI |
10.9 |
409 |
16 VII |
23.8 |
1083 |
||
|
2002 |
01 VI |
20.2 |
300 |
28 VI–02 VII |
15.0 |
770 |
26 VII |
19.6 |
1236 |
|
|
28 V |
7.5 |
239 |
28 VI–02 VII |
15.0 |
771 |
18 VII |
16.6 |
1081 |
||
|
A. semenovii |
2001 |
12 VI |
10.8 |
550 |
25–28 VI |
12.3 |
725 |
16 VII |
23.8 |
1083 |
|
01 VI |
8 |
215 |
01–05 VI |
10.9 |
409 |
16 VII |
23.8 |
1083 |
||
|
2002 |
01 VI |
20.2 |
300 |
20–24 VII |
13.4 |
656 |
26 VII |
19.6 |
1236 |
|
|
28 V |
7.5 |
239 |
12–16 VI |
13.7 |
526 |
22 VII |
17.8 |
1158 |
||
|
A. tataricum |
2001 |
12 VI |
10.8 |
550 |
29 VI–2 VII |
20.4 |
807 |
16 VII |
23.8 |
1083 |
|
01 VI |
8 |
363 |
29 VI–03 VII |
20.2 |
826 |
16 VII |
23.8 |
1083 |
||
|
2002 |
04 VI |
9.1 |
341 |
24–28 VI |
14.9 |
711 |
30 VII |
21.5 |
1316 |
|
|
01 VI |
20.2 |
300 |
20–24 VI |
13.4 |
656 |
22 VII |
17.8 |
1158 |
||
Обнаруженная изменчивость в продолжительности и интенсивности роста побегов обусловливает и соответствующее различие в величине их годичного прироста. При этом погодичная вариабельность длины побегов у всех изучаемых видов Acer весьма значительна, достигая 30– 400 %. Из данных табл. 3 следует, что в 2001 г. самые длинные побеги сформировались у A. negundo (215 мм). На следующий год самые короткие побеги были характерны для этого вида, а также для A. platanoides (29–50 мм). Вероятно, сочетание погодных условий конкретного года специфически сказывается на интенсивности деятельности апикальных меристем того или иного вида. Анализ результатов исследований свидетельствует о том, что величина годичного прироста побегов обусловлена соответствующими различиями в интенсивности роста, а не в его продолжительности. Так, длина стебля у A. semenovii в 1,5–2 раза боль- ше, чем у A. tataricum, а продолжительность роста у них одинакова.
Физиологические реакции растений, в том числе и ростовые, определяются состоянием среды и диапазоном толерантности вида к экологическим факторам (закон Шелфорда). Следовательно, установив значение факторов среды в ключевые периоды роста, а также форму и силу связи между динамикой прироста и изменчивостью этих факторов, можно судить о степени их соответствия требованиям организма.
Результаты исследований показали, рост побегов у изучаемых видов Acer может начаться при повышении среднесуточной температуры воздуха до 9-11 ° С (табл. 2). Кроме того, начало этой фенофазы зависит и от температуры воздуха предшествующего периода. К этому времени сумма положительных температур достигает 239-241 ° С.
Таблица 3
Таблица 4
Некоторые характеристики прироста побегов (над чертой, мм) и листьев (под чертой, мм 2 ) у различных видов Acer
|
Вид |
I >5 Л S К К CD Л 1=1 1—1 to |
Максимальный суточный прирост |
Годичный прирост |
Продолжительность роста, сут |
|
2001 |
4.2 |
72 |
37 |
|
|
Acer ginnala |
123 |
3680 |
46 |
|
|
2002 |
2.2 |
58 |
52 |
|
|
118 |
4020 |
55 |
||
|
2001 |
12.7 |
215 |
34 |
|
|
A. negundo |
150 |
4950 |
46 |
|
|
2002 |
2.0 |
50 |
52 |
|
|
112 |
5010 |
55 |
||
|
2001 |
1.2 |
29 |
34 |
|
|
A. platanoides |
302 |
10670 |
46 |
|
|
2002 |
1.2 |
38 |
44 |
|
|
305 |
10460 |
47 |
||
|
2001 |
8.2 |
179 |
34 |
|
|
A. pseudoplatanus |
628 |
18540 |
46 |
|
|
2002 |
2.7 |
86 |
56 |
|
|
542 |
18020 |
51 |
||
|
2001 |
11.2 |
174 |
34 |
|
|
A. semenovii |
110 |
3720 |
46 |
|
|
2002 |
2.7 |
109 |
56 |
|
|
102 |
4080 |
55 |
||
|
2001 |
3.2 |
65 |
34 |
|
|
A. tataricum |
112 |
4270 |
46 |
|
|
2002 |
3.2 |
77 |
56 |
|
|
145 |
5130 |
52 |
Во время прекращения роста побегов среднесуточная температура воздуха и сумма положительных температур варьируют в довольно широких пределах -17,8-23,8 ° С и 1015-1316 ° С соответственно. Эти данные свидетельствуют о том, что сроки прекращения этой фенофазы у представителей рода Acer не связаны с температурным режимом, а, скорее всего, обусловлены генотипом вида. Следовательно, тепла вполне достаточно для завершения годичного цикла развития вегетативных почек для всех изучаемых интроду-центов, что хорошо согласуется с выводами Л. А. Фроловой [12] и И. Т. Кищенко [13].
Требовательность растения к температуре воздуха в период максимального прироста побегов у различных видов Acer почти не различается. Кульминация приро ста побегов может наступить уже при повышении среднесуточной температуры воздуха до 13-15 ° С. Между тем, сумма положительных температур в этот период варьирует в широких пределах (656-965 ° С), что указывает на отсутствие заметного влияния данного параметра на интенсивность прироста побегов изучаемых видов.
Коэффициент корреляции между экологическими факторами и приростом вегетативных органов различных видов Acer
|
Вид |
2 ч to s S s и и CD Л 1=1 4 О |
cd 5 £ g I 2 и H |
^ о m |
# § о о S CD |
1=1 5 и а |
|
побеги |
|||||
|
Acer ginnala |
2001 |
0.12 |
0.62 |
0.10 |
0.58 |
|
2002 |
0.10 |
0.16 |
0.50 |
0.14 |
|
|
A. negundo |
2001 |
0.72 |
0.19 |
0.22 |
0.12 |
|
2002 |
0.62 |
0.37 |
0.15 |
0.10 |
|
|
A. platanoides |
2001 |
0.29 |
0.45 |
0.12 |
0.52 |
|
2002 |
0.27 |
0.20 |
0.25 |
0.15 |
|
|
A. pseudoplatanus |
2001 |
0.03 |
0.89 |
0.36 |
0.81 |
|
2002 |
0.11 |
0.12 |
0.48 |
0.52 |
|
|
A. semenovii |
2001 |
0.34 |
0.68 |
0.45 |
0.54 |
|
2002 |
0.29 |
0.34 |
0.44 |
0.18 |
|
|
A. tataricum |
2001 |
0.26 |
0.69 |
0.40 |
0.55 |
|
2002 |
0.14 |
0.20 |
0.33 |
0.20 |
|
|
листья |
|||||
|
Acer ginnala |
2001 |
0.58 |
0.21 |
0.20 |
0.12 |
|
2002 |
0.31 |
0.42 |
0.37 |
0.37 |
|
|
A. negundo |
2001 |
0.46 |
0.30 |
0.17 |
0.19 |
|
2002 |
0.52 |
0.60 |
0.41 |
0.20 |
|
|
A. platanoides |
2001 |
0.30 |
0.17 |
0.08 |
0.66 |
|
2002 |
0.16 |
0.15 |
0.04 |
0.44 |
|
|
A. pseudoplatanus |
2001 |
0.66 |
0.38 |
0.43 |
0.20 |
|
2002 |
0.44 |
0.17 |
0.41 |
0.09 |
|
|
A. semenovii |
2001 |
0.73 |
0.49 |
0.19 |
0.25 |
|
2002 |
0.54 |
0.50 |
0.25 |
0.37 |
|
|
A. tataricum |
2001 |
0.23 |
0.16 |
0.10 |
0.12 |
|
2002 |
0.33 |
0.35 |
0.27 |
0.12 |
|
Данные табл. 4 свидетельствуют о том, что между интенсивностью роста побегов и температурой воздуха, как правило, прослеживается достоверная положительная и довольно слабая корреляция ( r =0,3). Наиболее сильная подобная связь характерна для A. negundo ( r =0,6…0,7).
Между динамикой суточного прироста побегов и изменчивостью относительной влажности воздуха также обычно прослеживается положительная корреляция слабой и средней силы ( r =0,2…0,5). Примерно такой же характер корреляции выявлен и между динамикой прироста побегов изучаемых видов Acer и выпадением атмосферных осадков.
Зависимость интенсивности роста побегов от солнечной радиации выражена еще менее заметно (r=0,2…0,3).
Проведенные исследования позволили установить, что сроки начала роста листьев (фенофаза «обособление листьев на побегах») изучаемых видов Acer могут варьировать в пределах
3–7 сут (рис. 2). Наступление этой фенофазы за годы исследований наблюдалось 24 V–1 VI. Погодичная изменчивость в сроках прекращения роста листьев составляет не более 3–6 сут. Наблюдается эта фенофаза 10–22 VII, причем у A. platanoides на несколько сут раньше, чем у других видов. Незначительные погодичные различия в сроках начала и окончания роста листьев обусловливают и небольшие изменения в продолжительности их формирования у тех или иных видов Acer . В зависимости от года исследования продолжительность роста листьев может изменяться от 46 до 55 сут (табл. 3).
Установлено, что время интенсивного прироста листьев довольно сильно связано с видовыми особенностями и из года в год может существенно варьировать. В первую половину июня эта фаза уже наступает у A. ginnala, A. semenovii и A. negundo, а у других видов – во вторую половину июня и начале июля. Величина максимального прироста листьев у разных видов Acer существенно различается. Его наибольшее значение (в среднем 570 мм/сут) обна-
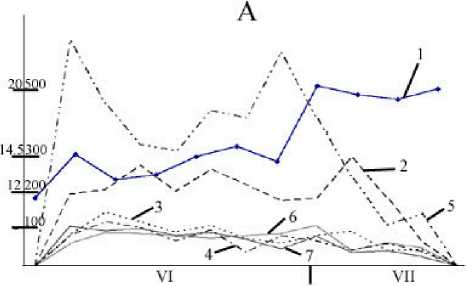
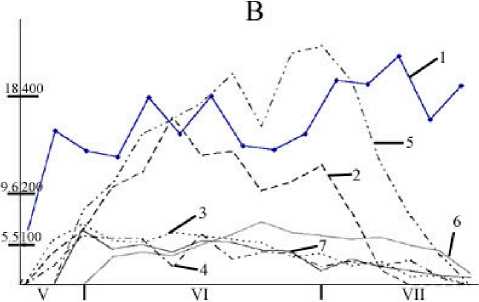
Рис. 2. Сезонная динамика температуры воздуха и суточного прироста листьев различных видов Acer
Годы: А - 2001, В - 2002. 1 — температура воздуха, 2 — Acer plata-noides; 3 — A. negundo; 4 — A. ginnala; 5 — A. pseudoplatanus;
6 — A. tataricum; 7 — A. semenovii. По вертикальным осям: слева — температура воздуха, ºС; справа – суточный прирост, мм2
ружено у A. ginnalа; у других видов этот показатель в 1.5–5 раз меньше. Следует отметить, что величина максимального прироста листьев у изучаемых видов Acer за годы исследований оставалась постоянной (табл. 3) .
Значительная изменчивость интенсивности роста листьев обусловливает соответствующие различия в величине их годичного прироста. Площадь листовой пластинки у A. ginnala, A. semenovii и A. tataricum , сформированной в разные вегетационные периоды, различалась не более чем на 8–16%, а у других видов оставалась практически без изменений. Из данных табл. 3 следует, что наибольшая площадь листа (в среднем 1.8 дм 2 ) формируется у A. pseudoplatanus , у A. platanoides она почти в 2 раза меньше, а у остальных видов она достигает всего 0.4–0.5 дм 2 . Различия в величине данного показателя обусловливаются интенсивностью, а не продолжительностью роста листьев. Так, площадь листа у A. pseudoplatanus в 4.5 раза больше, чем у A. semenovii, а продолжительность роста примерно такая же.
По данным трехлетних наблюдений, начало роста листьев при самой низкой среднесуточной температуре воздуха (2,5 ° С) отмечается у A. ginnala и A. negundo (табл. 2). Между тем начало данной фенофазы у других видов может отмечаться только при повышении температуры до +8 ° С. Рост листьев по площади заканчивает -ся в самый теплый за вегетацию период (17– 24 ° С), когда сумма положительных температур достигает 928-1158 ° С.
Требовательность растений к температуре воздуха в период максимального прироста листьев в значительной степени определяется биологией вида. Так, его наступление у A. plata-noides отмечено при температуре воздуха около 19 ° С, а для других видов - всего при 13-14 ° С. Между тем сумма положительных температур в этот период варьирует в широких пределах (409-905 ° С), что указывает на отсутствие явного влияния данного параметра на интенсивности прироста у всех изучаемых видов.
Проведение корреляционного анализа позволило установить, что так же, как и в отношении стеблей, между интенсивностью роста листьев, с одной стороны, и динамикой температуры и влажности воздуха, атмосферных осадков и солнечной радиации, с другой стороны, обычно прослеживается положительная и довольно заметная зависимость (табл. 4).
ВЫВОДЫ
-
1. Рост листьев у изученных видов Acer начинается в конце мая, а побегов – в первой половине июня. При этом различия между видами не превышают 3–4 суток.
-
2. Время прекращения роста побегов и листьев у разных видов Acer отличается не более чем на 3–7 суток и наблюдается во второй половине июля.
-
3. Благодаря максимальной интенсивности ростовых процессов наибольшая площадь листовой пластинки формируется у A. pseudoplatanus и у A. platanoides. Величина годичного прироста побегов у изученных видов претерпевает значительные изменения.
-
4. Сроки начала и кульминации прироста побегов и листьев определяются в основном температурой воздуха, в связи с чем они могут
-
5. Между интенсивностью прироста побегов и листьев, с одной стороны, и динамикой температуры и влажности воздуха, атмосферных осадков и солнечной радиации, с другой стороны, обычно прослеживается положительная и довольно заметная зависимость.
испытывать погодичную изменчивость в пределах 1–3 недель.
Список литературы Сезонный рост побегов представителей рода Аcer (Аceraceae) в условиях интродукции
- Калуцкий К. К., Болотов Н. А. Биоэкологические особенности лесной интродукции//Лесная интродукция. Воронеж, 1983. С. 4-14.
- Лапин П. И. Сезонный ритм развития древесных растений и его значение для интродукции//Бюл. ГБС АН СССР. 1967. Вып. 65. С. 12-18.
- Логинов В. Б. К методике построения частных теорий интродукции//Теории и методы интродукции растений и зеленого строительства: Материалы. Респ. конф. Киев, 1980. С. 58-60.
- Шестопалова В. В. Итоги интродукции сосновых (Pinaceae Lindl.) на Среднерусской возвышенности и перспективы их использования: Автореф. дисс... канд. биол. наук. Киев, 1982. 22 с.
- Мисник Г. Е. Сроки и характер цветения деревьев и кустарников. Киев: Наука, 1976. 215 с.
- Колесниченко А. Н. Сезонные ритмы развития древесных интродуцентов//Охрана, изучение и обогащение растительного мира. Киев, 1985. С. 21-32.
- Шутилов В. А. Интродукция видов рода клен в Камышинском дендрарии//Бюллетень ВНИИ агролесомелиорации. 1990. С. 32-44.
- Черепанов С. К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья. 1995. 992 с.
- Самцов А. С. Динамика роста листьев дуба черешчатого//Ботаника, исследования. 1966. Вып. 8. С. 52-61.
- Молчанов А. А., Смирнов В. В. Методика изучения прироста древесных растений. М.: Наука, 1967. 95 с.
- Шкутко Н. В. Хвойные Белоруссии. М.: Наука, 1991. 263 с.
- Фролова Л. А. Влияние температуры воздуха на сезонное развитие сосен в Ботаническом саду МГУ на Ленинских горах//Термический фактор в развитии растений различных географических зон: Тезисы докл. Всесоюз. конф. М., 1979. С. 37-39.
- Кищенко И. Т. Рост и развитие аборигенных и интродуцированных видов семейства Pinaceae Lindl. в условиях Карелии. Петрозаводск: Изд-во ПетрГУ, 2000. 211 с.
