Симбиотическая эффективность арктических штаммов ризобий на горошке (Vicia L.), копеечнике (Hedysarum L.), астрагале (Astragalus L.) и остролодочнике (Oxytropis DC.) в вегетационном опыте
, копеечнике (Hedysarum L.), астрагале (Astragalus L.) и остролодочнике (Oxytropis DC.) в вегетационном опыте")
Автор: Кузнецова И.Г., Гуро П.В., Сазанова А.Л., Сексте Э.А., Юзихин О.С., Тихомирова Н.Ю., Белимов А.А., Сафронова В.И., Карлов Д.С.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Бобово-ризобиальный симбиоз
Статья в выпуске: 5 т.60, 2025 года.
Бесплатный доступ
Растительно-микробные взаимоотношения, в частности бобово-ризобиальный симбиоз, играют одну из ключевых ролей в развитии сельского хозяйства. Особую значимость эта роль приобретает при создании устойчивого земледелия на арктических территориях России в экстремальных почвенно-климатических условиях. Изучение таксономического разнообразия арктических ризобий и их симбиотической активности в отношении различных видов бобовых растений позволяет определить хозяйскую специфичность штаммов и выявить наиболее эффективные азотфиксаторы, адаптированные к северным условиям. В настоящей работе впервые в условиях вегетационных опытов показана симбиотическая эффективность арктических штаммов Rhizobium beringeri P8/5-2, Rhizobium sp. 20-1/1, Mesorhizobium norvegicum 20/1-4 и Mesorhizobium sp. 9-4/1 и 25-2/1 на клубеньковых кормовых растениях Vicia sativa и V. cracca , а также на дикорастущих бобовых Hedysarum arcticum , Astragalus frigidus и Oxytropis adamsiana . Целью работы было изучение способности ранее селектированных арктических ризобиальных штаммов Rhizobium и Mesorhizobium из порядка Hyphomicrobiales формировать азотфиксирующий симбиоз с кормовыми викой посевной ( Vicia sativa L.) и викой мышиной ( V. cracca L.) и дикорастущими арктическими бобовыми копеечником арктическим ( Hedysarum arcticum B. Fedtsch) , астрагалом холодным ( Astragalus frigidus (L.) A. Gray) и остролодочником Адамса ( Oxytropis adamsiana (Trautv.) Jurtzev) в условиях вегетационного опыта по кросс-нодуляции. Работу проводили в лаборатории ФГБНУ ВНИИСХМ (г. Санкт-Петербург) в 2024 году. Семена дикорастущих популяций бобовых растений V. cracca , H. arcticum , O. adamsiana и A. frigidus были собраны на о. Самойловский и о. Тит-Ары в 2021 году. Семена кормовой раннеспелой вики посевной (яровой) сорта Приобская 25 были приобретены у агрофирмы «Аэлита» (Россия). Штаммы Rhizobium beringeri P8/5-2 (RCAM06326), Rhizobium sp. 20-1/1 (RCAM05664), Mesorhizobium norvegicum 20/1-4 (RCAM05519), Meso-rhizobium sp. 9-4/1 (RCAM06350) и 25-2/1 (RCAM06353) были предоставлены Сетевой биоресурсной коллекцией в области генетических технологий для сельского хозяйства (ФГБНУ ВНИИСХМ, г. Санкт-Петербург). Они были выделены из клубеньков арктических бобовых Lat-hyrus pratensis , V. cracca , H. arcticum , A. frigidus и O. taimyrensis . Растения V. sativa и V. cracca культивировали в пластиковых контейнерах объемом 2 л, содержащих 2 кг стерильного песка. Растения H. arcticum, O. adamsiana и A. frigidus выращивали в пластиковых стаканах объемом 200 мл, содержащих 300 г стерильного песка. Контейнеры, содержащие по 7-8 проростков, и стаканы с 3 проростками, инокулировали суспензиями индивидуальных штаммов в количестве 106 клеток на сосуд. Положительным контролем для Vicia служил коммерческий штамм R. leguminosarum bv. viciae RCAM0626. Неинокулированные растения были использованы в качестве отрицательного контроля. Растения выращивали в климатической камере MLR-352H («PHCbi», Япония). После окончания культивирования сырую биомассу побегов и корней взвешивали и считали число образовавшихся клубеньков. Азотфиксирующую активность клубеньков определяли ацетиленовым методом с помощью газового хроматографа GC-2014 («Shimadzu», Япония). Показана способность штамма R. beringeri P8/5-2 образовывать эффективный симбиоз с кормовыми бобовыми V. sativa и V. cracca , а после инокуляции дикорастущих арктических видов A. frigidus , O. adamsiana и H. arcticum , несмотря на отсутствие клубеньков, этот штамм приводил к достоверно более высоким значениям общей сырой массы растений в сравнении с отрицательным контролем. Штамм M. norvegicum 20/1-4 образовал эффективный симбиоз с H. arcticum , O. adamsiana и A. frigidus , однако после инокуляции этих видов бобовых штаммом Mesorhizobium sp. 9-4/1 симбиоз был наиболее эффективным. Так, прибавка общей массы растений A. frigidus составила от 13 до 37 %, O. adamsiana - от 32 до 174 %, тогда как H. arcticum - от 48 до 72 % в сравнении с остальными вариантами. При инокуляции растения-хозяина V. cracca штаммом Rhizobium sp. 20-1/1 симбиоз был эффективным, показав более высокие значения общей биомассы (прибавка составила 39 %), числа клубеньков и азотфиксирующей активности по сравнению с коммерческим штаммом R. leguminosarum bv. viciae RCAM0626. Таким образом, арктические штаммы Rhizobium sp. 20-1/1 и Mesorhizobium sp. 9-4/1 перспективны для дальнейших испытаний в условиях полевых опытов с целью создания на их основе высокоэффективных микробных препаратов для использования в северных регионах России при возделывании кормового бобового растения V. cracca и дикорастущих пастбищных бобовых, представителей родов Hedysarum , Оxytropis и Astragalus , как компонентов высокопродуктивных многолетних пастбищных и сенокосных сеяных лугов.
Арктический регион, бобово-ризобиальный симбиоз, азотфиксирующие клубеньковые бактерии
Короткий адрес: https://sciup.org/142247705
IDR: 142247705 | УДК: 579.64:579.262:582.736 | DOI: 10.15389/agrobiology.2025.5.898rus
Symbiotic efficiency of Arctic rhizobia strains on Vicia L., Hedysarum L., Astragalus L. and Oxytropis DC. in a vegetation experiment
Plant-microbe relationships, and in particular the legume-rhizobia symbiosis, play a key role in the development of agriculture. This role is of particular importance for the development of sustainable agriculture in the Arctic regions of Russia under extreme soil and climatic conditions. The study of the taxonomic diversity of Arctic rhizobia and their symbiotic activity with different legume species allows us to determine the host specificity of strains and to identify the most effective nitrogen fixers adapted to northern conditions. In this work, for the first time, under the conditions of pot experiments, the symbiotic efficiency of the arctic strains Rhizobium beringeri P8/5-2, Rhizobium sp. 20-1/1, Mesorhizobium norvegicum 20/1-4 and Mesorhizobium sp. 9-4/1 and 25-2/1 on the nodule forage plants Vicia sativa and V. cracca , as well as on the wild legumes Hedysarum arcticum , Astragalus frigidus and Oxytropis adamsiana is demonstrated. The aim of the work was to investigate the ability of arctic rhizobial strains Rhizobium and Mesorhizobium of the order Hyphomicrobiales to form nitrogen-fixing symbioses with forage plants ( V. sativa and V. cracca ) and wild Arctic legumes H. arcticum , A. frigidus and O. adamsiana under the conditions of a pot experiment on cross-nodulation. The work was carried out in FSBSI ARRIAM in 2024. Seeds of wild populations of the legumes V. cracca , H. arcticum , O. adamsiana and A. frigidus were collected during a Russian-German expedition to the Lena River Delta (Arctic zone of the Republic of Sakha (Yakutia)) in 2021. Seeds of fodder V. sativa Priobskaya 25 were purchased from Aelita agricultural company (RF). Arctic strains R. beringeri strains P8/5-2 (RCAM06326), R hizobium sp. 20-1/1 (RCAM05664), M. norvegicum 20/1-4 (RCAM05519), Mesorhizobium sp. 9-4/1 (RCAM06350) and 25-2/1 (RCAM06353) were provided by the Network Bioresource Collection in the Field of Genetic Technologies for Agriculture (FSBSI ARRIAM, St. Petersburg). They were isolated from nodules of the arctic legumes Lathyrus pratensis , V. cracca , H. arcticum , A. frigidus , and O. taimyrensis . V. sativa and V. cracca plants were grown in 2-liter plastic containers containing 2 kg of sterile sand. H. arcticum , O. adamsiana , and A. frigidus plants were grown in 200-ml plastic beakers containing 300 g of sterile sand. Containers with 7-8 seedlings and beakers with 3 seedlings were inoculated with suspensions of individual strains at a rate of 106 cells per vessel. The commercial strain R. leguminosarum bv. viciae RCAM0626 served as a positive control for Vicia . Uninoculated plants were used as a negative control. Plants were grown in a climate chamber MLR-352H (PHCbi, Japan). After cultivation, the fresh biomass of the shoots and roots was weighed, and the number of formed nodules was counted. The nitrogen-fixing activity of the nodules was determined by the acetylene method using a GC-2014 gas chromatograph (Shimadzu, Japan). The ability of the R. beringeri P8/5-2 strain to form an effective symbiosis with the forage legumes V. sativa and V. cracca was demonstrated. After inoculation with the wild arctic species A. frigidus , O. adamsiana , and H. arcticum , despite the absence of nodules, this strain resulted in significantly higher total fresh mass values of plants compared to the negative control. The M. norvegicum strain 20/1-4 formed an effective symbiosis with H. arcticum , O. adamsiana , and A. frigidus , but after inoculation of these legume species with the strain Mesorhizobium sp., the symbiosis was most effective. Thus, the increase in the total plant weight of A. frigidus ranged from 13 to 37 %, O. adamsiana from 32 to 174 %, while H. arcticum from 48 to 72 % compared to the other variants. When the host plant V. cracca was inoculated with the Rhizobium sp. strain, the symbiosis was effective, showing higher values of total biomass (the increase was 39 %), the number of nodules, and nitrogen-fixing activity compared to the commercial strain R. leguminosarum bv. viciae RCAM0626. Thus, the Arctic strains Rhizobium sp. 20-1/1 and Mesorhizobium sp. 9-4/1 are promising for further testing under field conditions to create highly effective microbial preparations for use in the northern regions of Russia in the cultivation of the forage legume V. cracca and wild pasture legumes of the genera Hedysarum , Oxytropis and Astragalus as components of highly productive perennial pasture and hay sown meadows.
Текст научной статьи Симбиотическая эффективность арктических штаммов ризобий на горошке (Vicia L.), копеечнике (Hedysarum L.), астрагале (Astragalus L.) и остролодочнике (Oxytropis DC.) в вегетационном опыте
Обеспечение устойчивого развития сельского хозяйства в Арктической зоне России представляет собой нетривиальную задачу ввиду очень непростых климатических и экологических условий в этом регионе. Однако возможные изменения климата могут привести к большей доступности арктических территорий, что открывает значительные экономические перспективы (1, 2). Существенный интерес представляют механизмы и закономерности адаптации организмов к экстремальным высокоширотным условиям Севера. В таких исследованиях арктические виды растений служат модельными объектами (3, 4).
Роль бобовых в экосистемах Арктики во многом определяется их биоценотическими и симбиотическими связями. Способность бобовых к симбиозу с азотфиксирующими бактериями позволяет им осваивать обедненные почвы и щебнистые осыпи, которые широко распространены на Севере (1, 3). Хозяйственная ценность бобовых определяется высокой урожайностью, хорошими кормовыми достоинствами, поедаемостью зеленой массы, сена и соломы, а также положительным эффектом при севооборотах.
Вика посевная ( Vicia sativa L.) представляет собой ценное кормовое растение, поскольку отвечает всем предъявляемым требованиям. Кормовые достоинства зеленой массы вики заметно превосходят гороховую, она содержит меньше клетчатки, долго не грубеет и поедается всеми видами животных (5). Недавно был обоснован агроклиматический потенциал возделывания вики посевной в условиях Центральной Якутии (6).
К бобовым растениям, перспективным для внедрения в арктические многолетние фитоценозы относится и вика мышиная ( Vicia cracca L.). Этот вид обладает широкой биологической пластичностью, имеет обширный па-леоарктический ареал и по химическому составу не уступает лучшим кормовым травам (7). Сорта V. cracca входят в Государственный реестр селекционных достижений, допущенных к использованию, и активно культивируются в качестве ценной кормовой травы в пастбищных и сенокосных фитоценозах (8).
В умеренных и арктических зонах России широко представлены растения из родов Копеечник (Hedysarum L.), Остролодочник (Îxytropis DC.) и Астрагал (Astragalus L.), которые входят в рацион питания различных животных и птиц (9, 10). Так, холодоустойчивое бобовое Astragalus frigidus (L.) A.Gray широко распространено за полярным кругом, эту высокобелковую кормовую культуру поедают северные олени (9). Вид Oxytropis adamsiana (Trautv.) Jurtzev распространен в арктической тундре, редколесьях, в высокогорьях на каменистых склонах, осыпях, и по берегам горных рек. Растение хорошо поедается травоядными животными (10). Вид включен в состав коллекции Якутского ботанического сада Института биологических проблем криолитозоны СО РАН (г. Якутск, Республика Саха) и изучается в качестве перспективного для дальнейшей интродукции в условиях резко континентального климата Якутии (11). Копеечник арктический (Hedys-arum arcticum B. Fedtsch.) представляет интерес для введения в состав бобово-злаковых агрофитоценозов в северных регионах России. Была показана высокая биологическая продуктивность естественных растительных сообществ с преобладанием H. arcticum по сравнению с фитоценозами, основанными на других местных видах бобовых в условиях Большеземель-ской тундры (12).
Известно, что для представителей рода Vicia наиболее характерны симбиотические отношения с различными штаммами вида Rhizobium leguminosarum bv. viciae . Однако описаны и другие микросимбионты, относящиеся к родам Rhizobium , Mesorhizobium , Phyllobacterium , Bosea и Tardiphaga , которых можно обнаружить в клубеньках вики (13-17). В частности, из клубеньков V. cracca , произрастающей в арктическом регионе России, были выделены штаммы R. herbae и B. psychrotolerans (17). Показано, что представители Astragalus и Oxytropis обладают широким разнообразием микросимбионтов, относящихся к родам Rhizobium , Sinorhizobium , Bradyrhizobium , Meso-rhizobium , Bosea и Tardiphaga (17-22). Так, для Astragalus frigidus из Арктической зоны Якутии описаны представители родов Mesorhizobium и Bosea (17), тогда как для растений O. adamsiana , собранных в дельте реки Лены и на плато Путорана, выделенные штаммы были отнесены к Pararhizobium , Neo-rhizobium , Phyllobacterium , Bradyrhizobium , Bosea и Tardiphaga (данные не опубликованы). Судя по данным литературы, растения рода Hedysarum но-дулируются видами, родственными Rhizobium и Mesorhizobium (23, 24). В то же время в качестве микросимбионтов копеечника щетинистого ( Hedysarum gmelinii Ledeb. subsp. setigerum ), произрастающего в Байкальском регионе, были описаны представители Phyllobacterium и Bosea , а из бобового растения H. arcticum , произрастающего в арктической Якутии, были выделены микросимбионты R. giardinii и M. norvegicum (25).
Эффективность возделывания бобовых растений зависит не только от параметров почвенно-климатической зоны, но и правильного подбора микросимбионта. Так, формирование растительно-микробной системы, включающей виды (сорта) растений и комплементарные им бактериальные штаммы, позволяет повысить адаптивность макросимбионта к условиям возделывания и прибавку урожайности. Для увеличения продуктивности бобово-ризобиального взаимодействия подбирают наиболее эффективные пары из уже существующих сортов и штаммов или проводят сопряженную селекцию, которая заключается в одновременном создании исходного сорта растения и генетически подходящего ему комплементарного штамма полезных микроорганизмов (26). Арктические виды бобовых растений и их микросимбионты уже адаптированы к почвенно-климатическим условиям Крайнего Севера, поэтому изучение хозяйской специфичности и симбиотической эффективности арктических ризобий в отношении растений-хозяев важно для развития сельского хозяйства в условиях Крайнего Севера.
В настоящей работе впервые в условиях вегетационных опытов показана симбиотическая эффективность арктических штаммов Rhizobium beringeri P8/5-2, Rhizobium sp. 20-1/1, Mesorhizobium norvegicum 20/1-4 и Mesorhizobium sp. 9-4/1 и 25-2/1 на клубеньковых кормовых растениях Vicia sativa и V. cracca , а также на дикорастущих бобовых Hedysarum arcticum , Astragalus frigidus и Oxytropis adamsiana .
Целью работы было изучение способности ранее селектированных арктических ризобиальных штаммов Rhizobium и Mesorhizobium из порядка Hyphomicrobiales формировать азотфиксирующий симбиоз с кормовыми викой посевной ( Vicia sativa L.) и викой мышиной ( V. cracca L.) и дикорастущими арктическими бобовыми копеечником арктическим ( Hedysarum arcticum B. Fedtsch) , астрагалом холодным ( Astragalus frigidus (L.) A. Gray) и остролодочником Адамса ( Oxytropis adamsiana (Trautv.) Jurtzev) в условиях вегетационного опыта по кросс-нодуляции.
Ìåòîäèêà. Для постановки вегетационного опыта по кросс-нодуля- ции (2024 год) использовали семена дикорастущих арктических (V. cracca, H. arcticum, O. adamsiana и A. frigidus) и культурных (V. sativa) бобовых растений, а также пять арктических ризобиальных штаммов из рода Rhizobium и Mesorhizobium.
Семена дикорастущих популяций бобовых растений V. cracca , H. arcticum , O. adamsiana и A. frigidus были собраны на о. Самойловский (72 ° 22 ‘ 00 ‘‘N 126 ° 30 ‘ 0 ‘‘E) и о. Тит-Ары (71 ° 57 ‘ 32 ‘‘ N 127 ° 05 ‘ 53 ‘‘ Е) в ходе российско-немецкой экспедиции в дельту р. Лены (арктическая зона Республики Саха — Якутия) в 2021 году. Семена кормовой раннеспелой вики посевной (яровой) сорта Приобская 25 приобретены у агрофирмы «Аэлита» (Россия).
Штаммы Rhizobium beringeri P8/5-2 (RCAM06326), Rhizobium sp. 201/1 (RCAM05664), Mesorhizobium norvegicum 20/1-4 (RCAM05519), Mesorhiz-obium sp. 9-4/1 (RCAM06350) и 25-2/1 (RCAM06353) были предоставлены Сетевой биоресурсной коллекцией в области генетических технологий для сельского хозяйства Всероссийского НИИ сельскохозяйственной микробиологии (ФГБНУ ВНИИСХМ, г. Санкт-Петербург) . Ранее эти штаммы были выделены из клубеньков арктических бобовых Lathyrus pratensis, V. cracca, H. arcticum, A. frigidus и O. taimyr-ensis (17, 25, 27, 28).
Семена растений скарифицировали и обрабатывали 98 % раствором серной кислоты (H 2 SO 4 ) в течение 5 мин (семена копеечника, астрагала и остролодочника) и 10 мин (семена вики), после чего их дважды промывали стерильной водопроводной водой в течение 15 мин. Эффективность обработки контролировали, высевая часть семян на чашки Петри с мясопептонным агаром (МПА) и бобовым агаром с последующей инкубацией при 28 ° С в течение 72 ч. Отсутствие роста микроорганизмов подтверждало стерильность семян. Проращивали семена на фильтровальной бумаге в чашках Петри при 25 ° С в темноте в течение 3-5 сут (в зависимости от вида растения).
Растения V. sativa и V. cracca культивировали в пластиковых контейнерах объемом 2 л, содержащих 2 кг стерильного песка. Растения H. articum, O. adamsiana и A. frigidus выращивали в пластиковых стаканах объемом 200 мл, содержащих 300 г стерильного песка. В каждый контейнер добавляли 400 мл, в стакан — 60 мл среды Красильникова-Кореняко (K 2 HPO 4 — 1,0, MgSO 4 7H 2 O — 1,0, Ca 3 (PO 4 ) 2 — 0,2, FeSO 4 – 0,02 г/л), а также смесь микроэлементов по М.В. Федорову (H 3 ВО 3 — 0,05, (NH 4 ) 2 MoO 4 — 0,05, KCl — 0,005, NaBr — 0,005, ZnSO 4 7H 2 O — 0,003, MnSO 4 — 0,002 г/л) объемом 40 мл на контейнер и 10 мл на стакан.
Контейнеры с 7-8 проростками в каждом и стаканы с 3 проростками инокулировали суспензиями индивидуальных штаммов в количестве 106 клеток на сосуд. Каждый вариант инокуляции V. sativa и V. cracca был представлен 14 повторностями, тогда как варианты H. arcticum, O. adamsiana и A. frigidus — по 6 повторностей. Положительным контролем для Vicia служил коммерческий штамм R. leguminosarum bv. viciae RCAM0626 из Сетевой био-ресурсной коллекции в области генетических технологий для сельского хозяйства (ФГБНУ ВНИИСХМ, г. Санкт-Петербург). Неинокулированные растения использовали в качестве отрицательного контроля.
Растения выращивали в климатической камере MLR-352H («PHCbi», Япония) при 18-22 ° С в течение 30 сут при относительной влажности 50 % и четырехуровневом режиме освещения/температуры: ночь (18 ° С, 8 ч), утро (200 мкмоль квантов • м - 2 • с - 1, 20 ° С, 2 ч), день (400 мкмоль квантов • м - 2 • с - 1, 23 ° С, 12 ч), вечер (200 мкмоль квантов • м - 2 • с - 1, 20 ° С, 2 ч). Освещение осуществлялось лампами L36W/77 FLUORA («Osram Licht AG»,
Германия). После окончания культивирования сырую биомассу побегов и корней взвешивали и подсчитывали образовавшиеся клубеньки. Азотфик-сирующую активность клубеньков определяли ацетиленовым методом с помощью газового хроматографа GC-2014 («Shimadzu», Япония).
Реизоляты выделяли общепринятыми микробиологическими методами (28). Принадлежность реизолятов к интродуцированным штаммам проверяли методом секвенирования последовательностей гена 16S rРНК (1400 п.н.).
Геномную ДНК из чистых культур выделяли с помощью наборов DNeasy Blood&Tissue kit («QIAGEN N.V.», Германия). Для амплификации использовали пару праймеров fD1 5´-AGAGTTTGATCCTGGCTCAG-3´ и rD1 5´-AAGGAGGTGATCCAGCC-3´ (29). Состав реакционной смеси для ПЦР: 38 мкл milli-Q H 2 O (ОАО «Евроген», Россия), 5 мкл буфера («Хели-кон», Россия), 5 мкл набора dNTP (ОАО «Евроген», Россия), по 0,5 мкл праймеров fD1и rD1 (ОАО «Евроген», Россия), 0,5 мкл Taq-полимеразы («Хеликон», Россия) и 1 мкл (50-100 нг) мДНК. Амплификацию проводили на приборе T100 Thermal Cycler («Bio-Rad», США). Условия ПЦР были следующими: 3 мин 30 с при 95 ° С (первичная денатурация); 1 мин 10 с при 94 ° С (денатурация), 40 с при 56 ° С (отжиг праймеров), 2 мин 10 с при 72 ° С (элонгация) (35 циклов); 6 мин 10 с при 72 ° С (финальная элонгация). Размер и количество амплифицированных фрагментов ДНК оценивали визуально с помощью электрофореза в 1,0 % агарозном геле в 0,5½ TАE-буфере с использованием маркера молекулярных масс MassRuler («Fermentas», Литва). Очистку ПЦР-продукта из агарозного геля осуществляли с помощью набора Cleanup S-Cap (ОАО «Евроген», Россия).
Секвенирование по Сэнгеру проводили на генетическом анализаторе ABI PRISM 3500xl («Life Technologies», США) на базе Центра коллективного пользования «Геномные технологии, протеомика и клеточная биология» Всероссийского НИИ сельскохозяйственной микробиологии. Полученные последовательности анализировали с помощью программы ChromasLite 2.6.4 . Последовательности сравнивали с нуклеотидной базой данных GenBank , используя программу BLAST.
Данные вегетационного опыта обрабатывали стандартным методом однофакторного дисперсионного анализа (ANOVA) в среде R (v. 4.2.3; R Core Team, 2023). Определяли средние значения ( M ) и их стандартные ошибки (±SEM) для каждого из измеряемых симбиотических параметров в каждом варианте инокуляции. При статистически значимых результатах ANOVA (p < 0,05) проводили тест Дункана для множественного сравнения средних с помощью пакета agricolae.
Ðåçóëüòàòû. Семена и клубеньки дикорастущих бобовых растений, из которых выделены ризобиальные штаммы, были собраны в г. Норильске, на о. Самойловский и о. Тит-Ары (рис. 1.).
На растениях V. sativa эффективные клубеньки образовались после инокуляции штаммом R. beringeri P8/5-2, однако число клубеньков, азот-фиксирующая активность и общая масса растений были достоверно ниже (p < 0,05) в сравнении с коммерческим штаммом R. leguminosarum bv. viciae RCAM0626. После инокуляции Rhizobium sp. 20-1/1 формировались неэффективные клубеньки, однако общая сырая биомасса была ниже в сравнении с вариантом без инокуляции (табл. 1).
На растениях V. cracca азотфиксирующие клубеньки были обнаружены в вариантах с инокуляцией R. beringeri P8/5-2 и Rhizobium sp. 20-1/1, при этом оба варианта обработки характеризовались достоверно более высо- кими значениями (p < 0,05) симбиотических параметров (числа клубеньков, общей сырой биомассы и азотфиксирующей активности) в сравнении со штаммом R. leguminosarum bv. viciae RCAM0626. Отметим, что после инокуляции Rhizobium sp. 20-1/1 наблюдались достоверно более высокие значения сырой биомассы растений и азотфиксирующей активности в сравнении с R. beringeri P8/5-2 (см. табл. 1).
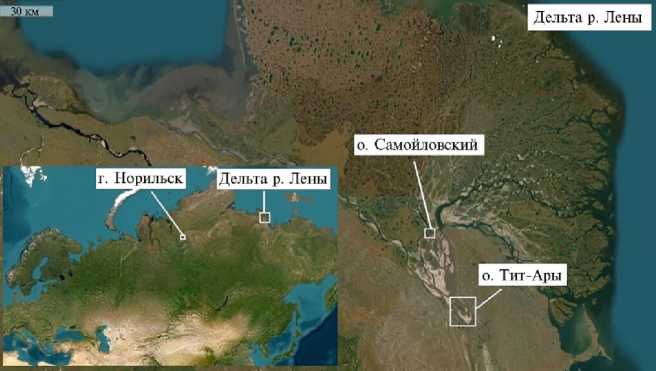
Места сбора семян и клубеньков дикорастущих бобовых растений в г. Норильске (Красноярский край), на о. Самойловский и о. Тит-Ары (дельта реки Лены, Северная Якутия). Карта с сайта (ESRI Sputnik), 2025.
-
1. Эффект инокуляции бобовых растений Vicia sativa и V. cracca арктическими изолятами из рода Rhizobium и коммерческим штаммом R. leguminosarum bv. viciae RCAM0626 в условиях вегетационного опыта ( n = 14, Ì ±SЕМ; вегетационный опыт, 2024 год)
-
2. Эффект инокуляции бобовых растений Hedysarum arcticum , Astragalus frigidus и Oxytropis adamsiana арктическими изолятами из родов Rhizobium и Meso-rhizobium ( n = 6, Ì ±SЕМ; вегетационный опыт, 2024 год)
|
Вариант инокуляции |
Число клубеньков на растение |
Сырая биомасса, мг/растение |
Ацетилен-редуктаз-ная активность, мкмоль C 2 H 4 растение - 1 сут - 1 |
||
|
побег |
корень |
общая масса |
|||
|
Без инокуляции |
0 d |
V. sativa 81,6±18,69b 118,9±19,54b |
200,4±19,47c |
0c |
|
|
R. leguminosarum bv. viciae RCAM0626 |
128,1±20,49b |
161,6±33,52a |
139,4±24,75a |
333,5±71,56a |
10,6±0,42a |
|
Rhizobium sp. 20-1/1 |
153,4±33,75a |
86,3±21,55b |
110,5±20,76b |
196,8±39,90c |
0c |
|
Rhizobium beringeri P8/52 |
76,2±12,70c |
155,6±30,69a |
122,1±23,42ab |
277,7±51,89b |
6,0±1,13b |
|
Без инокуляции |
0c |
V. cr 15,0±1,63b |
acca 10,6±1,51b |
23,9±1,68b |
0d |
|
R. leguminosarum bv. viciae RCAM0626 |
18,3±2,27b |
10,6±1,15c |
10,6±1,55b |
21,3±2,33c |
0,2±0,03c |
|
Rhizobium sp. 20-1/1 |
24,2±3,56a |
16,9±1,98a |
12,7±1,55a |
29,6±2,75a |
8,8±0,20a |
|
Rhizobium beringeri P8/5-2 |
22,1±2,74a |
10,2±1,26c |
7,9±1,91c |
18,1±2,64d |
3,3±0,05b |
|
Примечание. Штамм Rhizobium |
beringeri P8/5- |
2 был идентифицирован на уровне вида в работе |
|||
I.G. Kuznetsova с соавт. (16).
a, b и c Разными латинскими буквами отмечены варианты, различия между которыми статистически зна- чимы (критерий Дункана, p < 0,05).
На растениях H. arcticum азотфиксирующие клубеньки образовались в вариантах со штаммами M. norvegicum 20/1-4 и Mesorhizobium sp. 25-2/1, неэффективные клубеньки — с Mesorhizobium sp. 9-4/1, тогда как в варианте с R. beringeri P8/5-2 клубеньков обнаружено не было. После инокуляции M. norvegicum 20/1-4, Mesorhizobium sp. 9-4/1 и R. beringeri P8/5-2 наблюдались достоверно более высокие значения (p < 0,05) для общей биомассы растений в сравнении с отрицательным контролем, тогда как обработка штаммом Mesorhizobium sp. 25-2/1 не приводила к статистически значимому увеличе-нию общей биомассы растений (табл. 2).
|
Вариант инокуляции |
Число клубеньков на растение |
Сырая биомасса, мг/растение |
Ацетилен-редуктаз-ная активность, мкмоль C 2 H 4 растение - 1 сут - 1 |
||
|
побег |
корень |
общая масса |
|||
|
Без инокуляции |
0c |
H. arcticum 34,0±4,83d 38,1±5,43c |
72,1±7,78b |
0 b |
|
|
Mesorhizobium norvegicum 20/1-4 |
4,2±0,84a |
54,0±11,40ab |
58,0±13,04b |
112,0±22,80a |
0,3±0,02a |
|
Mesorhizobium sp. 25-2/1 |
3,4±0,70b |
42,0±4,83cd |
42,0±4,83c |
84,0±8,10b |
0,3±0,03a |
|
Mesorhizobium sp. 9-4/1 |
4,5±1,01a |
63,3±7,91a |
61,1±6,51ab |
124,4±12,86a |
0b |
|
Rhizobium beringeri 8/5-2 |
0c |
47,9±13,39bc |
68,3±11,36a |
116,2±21,37a |
0b |
|
Без инокуляции |
0 d |
A. frigidus 23,3±5,16с 31,7±7,53 |
55,0±10,49с |
0 d |
|
|
Mesorhizobium norvegicum 20/1-4 |
2,5±0,53c |
28,8±4,43b |
36,3±5,18 |
65,0±8,45b |
0,5±0,05b |
|
Mesorhizobium sp. 25-2/1 |
5,0±1,10a |
30,8±3,546b |
35,5±3,02 |
66,3±2,07b |
0,2±0,04c |
|
Mesorhizobium sp. 9-4/1 |
4,1±0,69b |
35,7±5,35a |
40,0±5,77 |
75,7±8,86a |
1,2±0,10a |
|
Rhizobium beringeri P8/5-2 |
0d |
26,9±2,67bc |
39,9±5,15 |
66,7±5,38b |
0d |
|
Без инокуляции |
0 c |
O. adamsiana 22,0±3,39c 32,2±4,09b |
54,2±7,40d |
0 d |
|
|
Mesorhizobium norvegicum 20/1-4 |
4,1±0,78b |
40,0±6,12b |
37,8±8,33b |
77,8±9,72c |
2,1±0,08c |
|
Mesorhizobium sp 25-2/1 |
4,9±0,60b |
38,9±6,51b |
40,0±7,07b |
78,9±9,93c |
2,6±0,30b |
|
Mesorhizobium sp 9-4/1 |
6,8±1,36a |
86,3±18,47a |
62,5±9,64a |
148,8±23,87a |
6,4±0,82a |
|
Rhizobium beringeri P8/5-2 |
0c |
42,0±5,48b |
70,6±7,86a |
112,6±11,92b |
0d |
Примечание. Штамм Mesorhizobium norvegicum 20/1-4 был идентифицирован на уровне вида в работе D.S. Karlov с соавт. (25).
a, b, c и d Разными латинскими буквами отмечены варианты, различия между которыми статистически значимы (критерий Дункана, p < 0,05). В вариантах без буквенного ранжирования статистически значимых групповых различий (р > 0,05) выявлено не было и тест Дункана не проводился.
На A. frigidus активные клубеньки обнаружили после инокуляции штаммами M. norvegicum 20/1-4, Mesorhizobium sp. 25-2/1 и 9-4/1, тогда как после инокуляции R. beringeri P8/5-2 клубеньков выявлено не было. Все варианты показали достоверно более высокие значения (p < 0,05) сырой биомассы растений по сравнению с отрицательным контролем. При этом штамм Mesorhizobium sp. 9-4/1 продемонстрировал наиболее высокие, статистически достоверные значения (p < 0,05) сырой биомассы и азотфикси-рующей активности в сравнении с остальными вариантами инокуляции.
На O. adamsiana эффективные клубеньки образовались после инокуляции M. norvegicum 20/1-4, Mesorhizobium sp. 25-2/1 и 9-4/1. В варианте с инокуляцией Mesorhizobium sp. 9-4/1 отмечали достоверно более высокие значения (p < 0,05) для всех исследуемых симбиотических параметров в сравнении с остальными вариантами. Обработка растений штаммами M. Norveg-icum 20/1-4 и Mesorhizobium sp. 25-2/1, а также не формировавшим клубеньков штаммом R. beringeri P8/5-2 привела к достоверно более высоким показателям (p < 0,05) массы побега и общей массы растений в сравнении с отрицательным контролем.
Все штаммы были повторно реизолированы из клубеньков, а их принадлежность к соответствующему инокуляту подтверждена с помощью секвенирования и анализа сходства маркерного гена 16S rRNA. У всех реизо-лированных штаммов сходство между собой по гену 16S rRNA варьировало от 93,60 до 99,86 %, тогда как сходство каждого реизолята с соответствующим штаммом-инокулятом составило 100 %.
Настоящее исследование представляет собой продолжение ранее проведенной работы по выделению, идентификации и отбору наиболее перспективных арктических клубеньковых бактерий в условиях стерильных микровегетационных опытов с кормовыми ( V. cracca , V. sativa , L. sativus , L. pratensis , T. repens и M. sativa ) и дикорастущими ( H. arcticum, A. frigidus и O. adamsiana ) бобовыми растениями (17, 25, 27, 28).
Штаммы Rhizobium sp. 20-1/1 и R. beringeri P8/5-2 были выделены нами из клубеньков бобовых, произрастающих в арктических регионах России, — соответственно V. cracca и L. pratensis (17, 27). Способность штамма Rhizobium sp. 20-1/1 формировать эффективный симбиоз с культурными V. cracca , V. sativa , T. repens L. и Medicago sativa L., а также дикорастущими бобовыми O. adamsiana и A. frigidus была изучена ранее в условиях стерильных микровегетационных опытов (МВО) (27, 28) . В частности, была показана способность Rhizobium sp. 20-1/1 формировать эффективный симбиоз только после инокуляции с V. cracca . При этом наблюдалась значительно более высокая азотфиксирующая активность по сравнению с коммерческим штаммом R. leguminosarum bv . viciae RCAM0626, что согласуется с результатами вегетационного опыта на песке в настоящей работе (см. табл. 1).
Однако отметим малую эффективность штамма R. leguminosarum bv . viciae RCAM0626 на растениях V. cracca , что, вероятно, связано с узкой специфичностью RCAM0626 к растению-хозяину V. sativa. В настоящее время коммерческие штаммы ризобий для V. cracca не описаны, поэтому высокоактивный штамм Rhizobium sp. 20-1/1 можно рекомендовать в качестве положительного контроля при постановках микровегетационных и вегетационных опытов.
После инокуляции Rhizobium sp. 20-1/1 культурных V. sativa , Trifolium repens L. и M. sativa L. клубеньки были неэффективными, тогда как на корнях дикорастущих бобовых O. adamsiana и A. frigidus клубеньков выявлено не было (27, 28) . Следовательно, штамм Rhizobium sp. 20-1/1 показал себя как высокоэффективный микросимбионт только во взаимодействии с V. cracca , что свидетельствует о его узкой специфичности в отношении растения-хозяина. Известно, что сужение хозяйской специфичности характерно для поздних этапов эволюции микросимбионтов и тесно связано с повышением их N 2 -фиксирующей активности (30), что хорошо согласуется с результатами настоящей работы.
В предыдущем исследовании у штамма R. beringeri P8/5-2 мы обнаружили основные sym -гены клубенькообразования ( nodA и nodC ) (17), а также выявили способность формировать клубеньки на корнях V. cracca, L. pratensis и T. repens в условиях микровегетационного опыта (данные не опубликованы), однако способность к азотфиксации не изучалась. В настоящем исследовании в условиях вегетационного опыта R. beringeri P8/5-2 формировал эффективный симбиоз с кормовыми бобовыми V. sativa и V. cracca , тогда как на дикорастущих арктических видах A. frigidus , O. adamsiana и H. arcticum клубеньков выявлено не было. Большая часть сформировавшихся клубеньков отличалась продолговатой формой и розоватым оттенком, свидетельствующим об их азотфиксирующей активности.
После инокуляции культурных бобовых V. sativa и V. cracca штаммом R. beringeri P8/5-2 наблюдались достоверно более низкие (p < 0,05) значения большинства симбиотических параметров (число клубеньков, азотфиксиру-ющая активность и общая биомасса) по сравнению соответственно с R. leguminosarum bv. viciae RCAM0626 и Rhizobium sp. 20-1/1. В то же время, несмотря на отсутствие способности R. beringeri P8/5-2 формировать клубеньки на дикорастущих растениях A. frigidus, H. arcticum и O. adamsiana, показатели массы побегов, корней и общей сырой биомассы были достоверно выше (p < 0,05) в сравнении с отрицательным контролем, что, вероятно, связано со способностью штамма продуцировать фитогормоны, влияющие на рост и развитие растений. Известно, что клубеньковые бактерии, в частности представители рода Rhizobium, могут выступать в качестве ассоциативных микросимбионтов для многих экономически ценных небобовых культур, выделяя ростостимулирующие и фунгистатические вещества, что приводит к улучшению продуктивности сельскохозяйственных культур (31, 32).
Штамм M. norvegicum 20/1-4 был ранее выделен нами из растений H. arcticum , произрастающих в дельте р. Лены в арктической зоне Якутии (25) . В настоящей работе M. norvegicum 20/1-4 формировал эффективный симбиоз с H. arcticum , O. adamsiana и A. frigidus в условиях вегетационного опыта, тогда как ранее была показана его способность формировать активные клубеньки на O. adamsiana и неэффективные клубеньки на A. frigidus в условиях МВО (28). Присутствие азотфиксирующей активности после инокуляции A. frigidus штаммом M. norvegicum 20/1-4 в условиях открытого опыта на песке в сравнении с результатом МВО на вермикулите может отражать наличие в первом случае более благоприятных условий для формирования эффективного бобово-ризобиального симбиоза арктических растений с ризобиями.
Штаммы Mesorhizobium sp. 9-4/1 и Mesorhizobium sp. 25-2/1 были выделены из клубеньков O. taimyrensis и A. frigidus , соответственно, произрастающих в арктической зоне Якутии (28). Показано, что штаммы формировали эффективный симбиоз с арктическими бобовыми O. adamsiana и A. frigidus и были не способны образовывать клубеньки на кормовом бобовом T. repens в условиях стерильного МВО (28).
В настоящей работе штамм Mesorhizobium sp. 9-4/1 формировал азот-фиксирующий симбиоз с A. frigidus и O. adamsiana , что в большинстве случаев приводило к достоверно более высоким (p < 0,05) значениям симбиотических параметров в сравнении с остальными вариантами инокуляции (см. табл. 2). Так, прибавка общей массы растений A. frigidus составила 37 %, тогда как O. adamsiana — 174 % относительно контроля без инокуляции. В случае с H. arcticum на корнях растений образовывались неэффективные клубеньки, что, однако, приводило к приросту общей сырой биомассы на 72 % в сравнении с отрицательным контролем.
Штамм Mesorhizobium sp. 25-2/1 формировал эффективный симбиоз с H. arcticum , O. adamsiana и A. frigidus в условиях вегетационного опыта, однако только в двух последних вариантах наблюдались достоверно более высокие (p < 0,05) значения общей сырой массы растений по сравнению с отрицательным контролем.
В работах других авторов была показана способность арктических штаммов, выделенных из клубеньков бобовых Astragalus и Oxytropis , эффективно нодулировать кормовое бобовое эспарцет виколистный ( Onobrychis viciifolia Scop.). Выявлено, что арктические ризобии эффективнее коммерческих штаммов в условиях низкотемпературных лабораторных и полевых опытов (33, 34). Решающую роль в инициировании симбиоза при низких температурах может играть АТФаза, транспортирующая β -глюкан, который участвует в прикреплении бактериальных клеток к корням растений. Способность адаптироваться к экстремально низким температурам также может быть связана с повышенным уровнем синтеза белков холодового шока у арктических ризобий (35, 36).
Таким образом, в вегетационном опыте по изучению кросс-нодуля- ции с участием кормовых бобовых Vicia sativa и V. cracca клубеньки формировались в обоих вариантах инокуляции штаммами Rhizobium sp. 20-1/1 и R. beringeri P8/5-2. Однако на V. sativa штамм Rhizobium sp. 20-1/1 формировал неэффективные клубеньки, тогда как в варианте инокуляции растения-хозяина V. cracca симбиоз был эффективным, показав более высокие значения общей биомассы (прибавка составила 39 %), числа клубеньков и азот-фиксирующей активности по сравнению с коммерческим штаммом R. leguminosarum bv. viciae RCAM0626. В отношении местных арктических видов Hedysarum arcticum, Oxytropis adamsiana и Astragalus frigidus более активными были штаммы, выделенные из тех же местных видов бобовых. Наибольший положительный эффект наблюдался после инокуляции штаммом Mesorhizobium sp. 9-4/1, который формировал активные клубеньки на A. frigidus и O. adamsiana. Так, прибавка общей массы растений A. frigidus составила от 13 до 37 %, O. adamsiana — от 32 до 174 %, H. arcticum — от 48 до 72 % в сравнении с остальными вариантами инокуляции. Следовательно, арктические штаммы Rhizobium sp. 20-1/1 и Mesorhizobium sp. 9-4/1 перспективны для дальнейших испытаний в условиях полевых опытов с целью создания на их основе высокоэффективных микробных препаратов для использования в северных регионах России при возделывании кормового бобового растения V. cracca и дикорастущих пастбищных бобовых. Штамм R. beringeri P8/5-2 не формировал клубеньков на H. arcticum, A. frigidus и O. adamsiana, однако оказывал положительный эффект на массу побегов и корней, не приводя к статистически значимому увеличению общей массы растений в сравнении с большинством вариантов инокуляции. Для изучения механизма такого эффекта необходимо проведение полногеномного секвенирования R. beringeri P8/5-2 с последующим поиском и изучением функциональных особенностей рост-стимулирующих генов, отвечающих за синтез фитогормонов (ауксинов, гиббереллинов, цитокининов). Последующее изучение симбиотических взаимоотношений арктических ризобий с кормовыми сельскохозяйственными и дикорастущими бобовыми в условиях вегетационных и микровегетационных опытов будет способствовать поиску наиболее перспективных растительно-ризобиальных систем, адаптированных к агроэкологическим условиям Севера. Генетический материал описанных ризобий послужит ценным ресурсом при создании эффективных микробных препаратов, перспективных для развития экологически устойчивого сельского хозяйства на бедных мерзлотных почвах Арктики.
Быражаем благодарность руководству и координаторам экспедиции «Лены 2021» за организацию и проведение экспедиции в район дельты р. Лены. Искренне благодарим Сергея Александровича Иравкина (ААНИИ) за помощь в сборе и транспортировке семян бобовых культур. Быражаем благодарность коллективу научно-исследовательской станции «Остров Самойловский» и лично Федору Биссанионовичу Селляхову за предоставленный транспорт.
ФГБНУ Бсероссийский НИИ сельскохозяйственной микробиологии,
Иоступила в редакцию 25 февраля 2025 года Принята к публикации 24 апреля 2025 года
Sel’skokhozyaistvennaya biologiya [ Agricultural Biology ], 2025, V. 60, ¹ 5, pp. 898-910
SYMBIOTIC EFFICIENCY OF ARCTIC RHIZOBIA STRAINS ON Vicia L., Hedysarum L., Astragalus L. AND Oxytropis DC. IN A VEGETATION EXPERIMENT
I.G. Kuznetsova 8 , P.V. Guro, A.L. Sazanova, E.A. Sekste, O.S. Yuzikhin, N.Yu. Tikhomirova, À.À. Belimov, V.I. Safronova, D.S. Karlov
The authors declare no conflict of interests
Acknowledgements:
We express our gratitude to the leadership and coordinators of the expedition « Lena 2021» for organizing and conducting the expedition to the Lena River Delta. We also sincerely thank Sergei Aleksandrovich Pravkin (AARI) for his assistance in collecting and transporting legume seeds. We also thank the staff of Research Station Samoilovsky Island and Fyodor Vissanionovich Selyakhov for providing transportation.
This work was conducted using equipment from the Genomic Technologies, Proteomics, and Cell Biology" Center for Collective Use at the All-Russian Research Institute of Agricultural Microbiology.
Supported financially by the Russian Science Foundation (RSF Project No. 20-76-10042-P)
