Синтаксономическое разнообразие и экотопы лесных сообществ поймы реки Лямин (Сургутское Полесье)
")
Автор: Кукуричкин Глеб Михайлович, Бабюк Сергей Иванович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В данной статье рассмотрено синтаксономическое разнообразие лесной растительности поймы реки Лямин (на уровне циклов и формаций). Показаны зависимость фиторазнообразия поймы от литологии бассейна реки и связь растительности и динамично изменяющихся аллювиальных почв.
Пойменные леса, аллювиальные почвы, эколого-фитоценотическая классификация
Короткий адрес: https://sciup.org/148200808
IDR: 148200808 | УДК: 581.9(571.122-282.256.149):581.55.051
Syntaxonomic diversity and forest habitats of the Liamin river floodplain (Surgut woodlands)
In the article the syntaxonomic diversity (up to cycles and formations) of forest vegetation of the Liamin River floodplain is described. The interrelations between the community diversity, the lithology of drainage basin and the soil dynamic are shown.
Текст научной статьи Синтаксономическое разнообразие и экотопы лесных сообществ поймы реки Лямин (Сургутское Полесье)
Река Лямин протекает с севера на юг по Сургутской низине: берёт начало на увале Нумто и впадает в Обь в 100 км к западу от Сургута. Общая площадь водосбора составляет 15,9 тыс. км2 [8].
Бассейн реки расположен преимущественно в пределах зандровой равнины тазовского оледенения [3], правый берег реки является западной границей Сургутского полесья [12]. Бассейн Лямина характеризуется наивысшими для региона показателями заболоченности (до 80 %) и заозеренности (до 20 %) [1].
К литологическим особенностям водосбора, обусловливающим особенности русловых деформаций, характер твердого стока и, в конце концов, специфику почвенно-растительного покрова поймы, в первую очередь можно отнести почти повсеместное распространение в бассейне песчаных отложений. Более тяжелые отложения характерны для правого коренного берега р. Лямин (так называемый Ляминский материк), но река не имеет никаких существенных притоков с этой стороны.
Гидрологическая система реки Лямин образована слиянием трех рек – Лямин 1-й, Лямин 2-й, Ля-мин-3-й, каждая из которых имеет длину около 200 км, и нескольких малых притоков; длина основного русла – 281 км. Питание реки смешанное, с преобладанием снегового. Половодье начинается в конце апреля и заканчивается в среднем течении к середине лета; причем на эту фазу приходится 50 % стока. Размах колебаний уровня воды – свыше 4,5 м, средний многолетний расход – более 90 м3/с [8].
Лямин – активно меандрирующая река. В ходе свободного меандрирования формируются основные зоны поймы – молодая, зрелая, старая, каждая из которых характеризуется определенным режимом ландшафтообразующих процессов [5 и др.].
Согласно ландшафтному районированию Ханты-Мансийского автономного округа, эта территория относится к средней тайге; а пойма р. Лямин – к интразональным лесным ландшафтам [1].
Согласно геоботаническому районированию Западной Сибири [4; 11], практически весь бассейн р. Лямин относится к северной тайге; преобладающим контуром здесь являются кустарничково-сфаг-
новые олиготрофные болотно-озерные комплексы. Долина реки относится к средней тайге – к серии ивово-березовых, березово-сосновых травяно-зеленомошных и лишайниковых лесов долинных грив и сосново-березовых осоково-сфагновых лесов меж-гривных понижений.
По нашим данным, леса занимают около 80 % площади современной поймы; остальное приходится на болотные, водные и луговые фитоценозы.
Экологические индексы большинства местообитаний изученных лесных сообществ по шкалам Л.Г. Раменского [10] соответствуют влажнолуговой серии по увлажнению и мезоолиготрофной серии – по богатству.
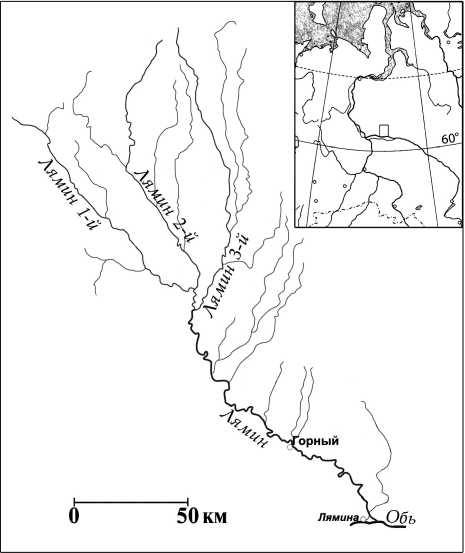
Рис . Район исследований
МАТЕРИАЛЫ И МЕТОДЫ
Изучение пойменных лесов осуществлялось в 2005-2007 гг. в рамках комплексной почвенногеоботанической экспедиции, проводившейся маршрутным методом – сплавом по реке Лямин и его притокам. Общая протяжённость обследованных участков речных пойм составляет около 300 км.
В связи с техническими особенностями водного маршрута в поле зрения исследователей попадали преимущественно доступные со стороны русла участки поймы. При планировании пеших маршрутов, а также для пространственной интерпретации выделенных синтаксонов и площадных оценок использовались материалы дистанционного зондирования.
Геоботанические описания выполнялись по традиционным методикам [9]. Относительное превышение экотопов над меженью определялось с использованием нивелира. Проводилась глазомерноинструментальная таксация древостоев и детальное исследование почв. Разнообразие аллювиальных почв р. Лямин рассмотрено в отдельной публикации [2].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Характеристика лесной растительности в настоящей работе приводится до уровня крупных синтаксонов эколого-фитоценотической классификации - циклов ассоциаций. Для каждого цикла можно выделить диагностические признаки - как во флористической композиции, так и в пространственной структуре сообществ. Общими видами почти для всех типов лесных сообществ поймы являются Calamagrostis purpurea s.l. и Vaccinium vitis-idaea . Активность первого максимальна в сообществах молодой поймы, второго - в зрелой пойме.
Почвы поймы р. Лямин типичны для большинства рек таежной зоны. К особенностям можно отнести однородность и крайнюю бедность почвообразующих пород.
Обсуждаемые циклы достаточно четко диагностируются, так как хорошо изучены на других реках региона [6 и др.], но в пойме Лямина охарактеризованы незначительным числом полных геоботанических описаний. Поэтому классы постоянства (к.п.) видов приводятся не для всех циклов.
Разнотравный цикл ( Varioherbosum ) приурочен к самым молодым аллювиальным поверхностям, намытым в последние 30-50 лет. Лесные ценозы формируются на месте первичных ястребинково-наземновейниковых лугов и других пионерных группировок растительности. Аллювиальные почвы характеризуются ослабленным дерновым процессом, в целом почвы примитивные, часто с захороненными задернованными горизонтами вследствие периодического пересыпания русловым аллювием. Сомкнутость древостоя 80-100 %. Подполо-говое пространство в этих лесных ценозах хорошо освещено даже при высокой сомкнутости - за счет того, что такие участки фрагментированы и по периферии окружены луговинами и водными пространствами. Сообщества цикла отличаются присутствием луговых видов, дающих несомкнутый травяной ярус (общее проективное покрытие (ОПП) не более 15 %, обычно менее 5 %). Обычны: Rumex thyrsiflorus (проективное покрытие (ПП) до 3 %), Sanguisorba officinalis (ПП до 4 %), Hieracium umbellatum (до 3 %), Solidago virgaurea (до 5 %),
Calamagrostis epigeios (до 1 %), C. purpurea (до 4 %). В моховом ярусе - представители рода Polytrichum , в разнообразных сочетаниях; в зависимости от условий аллювиальности ОПП яруса колеблется от 0 до 85 %. Цикл представлен молод-няками пионерных формаций - Pineta sylvestris и редко Betuleta pubescentis-pendulae . При высоте основного яруса в среднем 7-10 м здесь встречается низкорослый (до 1 м) разреженный подрост темнохвойных пород - кедра (до 2,5 тыс./га) и ели (до 0,4 тыс./га).
Сообщества мелкотравного цикла ( Parviherbo-sum ) встречаются преимущественно на повышенных краткопоемных участках молодой (прирусловой) поймы на аллювиальных слабодерновых маломощных почвах с менее выраженной современ -ной аллювиальностью. Для этого цикла характерны относительно хорошо развитый ярус подлеска (ПП 10-30 %) и фрагментированный моховой покров; в травяно-кустарничковом ярусе (ТКЯ) основную синузию составляет лесное мелкотравье. Основные виды (V к.п.): Padus avium (до 7 %), Sorbus sibirica (до 8 %), Maianthemum bifolium (до 25 %), Orthilia secunda (до 8 %), Trientalis europaea (ПП до 6 %), Pyrola minor (до 7%), Rubus arcticus (ПП до 20 %). Им обычно (IV к.п.) сопутствуют фоновые для поймы виды: Calamagrostis purpurea (ПП до 5 %), Vaccinium vitis-idaea (ПП до 10 %). Изредка (III к.п.) встречаются: Rosa acicularis (ПП до 10 %), Swida alba (ПП до 5 %). В моховом покрове (ОПП 5-50 %) выраженных доминантов нет; с различным постоянством и обилием встречаются: Polytrichum commune , Pleurozium schreberi , Hylocomium splendens . Цикл представлен двумя формациями: Pineta sylvestris и Betuleta pubescentis . В подросте доминирует кедр (до 6 тыс./га).
Княжениково-вейниковый цикл ( Ruboso arctici-calamagrostidosum purpurea ) приурочен к пониженным участкам молодой поймы, где откладывается более мелкодисперсный аллювий, а грунтовые воды в межень стоят близко к поверхности. Почвы аллювиальные дерновые слабо оглеенные маломощные. В травяном ярусе доминируют Calamagrostis purpurea (ПП 20-50 %) и Rubus arcticus (ПП 15-30 %). Им сопутствуют: Maianthemum bifolium (ПП до 10 %), Carex acuta (ПП в среднем 2 %), Naumburgia thyrsiflora (ПП в среднем около 1 %). Реже и с незначительным покрытием встречаются: Rosa majalis , Padus avium , Salix viminalis , S. dasyclados , Trientalis europaea , Juncus fili-formis . Моховой ярус отсутствует или представлен небольшими куртинками Polytrichum commune . В молодой пойме цикл представлен формацией Betuleta pubescentis . Кроме того, на «омолаживающихся» поверхностях зрелой поймы при длительном (несколько десятилетий) нахождении участка в условиях наложенного прируслового режима формируются вторичные осиновые сообщества этого цикла - Populeta tremulae . В подросте чаще преобладают мелколиственные породы (до 9 тыс./га), из темнохвойных - кедр (до 9 тыс./га).
Сообщества бруснично-зеленомошного цикла ( Vaccinioso-hylocomiosum ) наиболее широко распространены в зрелой (центральной) пойме р. Лямин, где занимают не менее 60 % облесенной площади. Это наиболее краткопоемные экотопы. Почвы с признаками оподзоливания, с выраженным оторфовыванием подстилки, в профиле преобладают окислительные процессы. Абсолютным до-минантом ТКЯ является Vaccinium vitis-idaea (V к.п.) с ПП от 10 до 60 % и более (за исключением «омоложенных» участков), а также хорошо развитым моховым покровом, в основном представленным Pleurozium schreberi (V к.п., ПП до 90 %), Polytrichum commune (V к.п., ПП до 45 %), Hylocomium splendens (IV к.п., ПП до 30 %). В ТКЯ обычны (IV к.п.): Linnaea borealis (до 10 %), Maianthemum bifolium (до 10-12 %), Trientalis eu-ropaea (ПП до 3 %), реже встречаются (III к.п.): Orthilia secunda (до 4 %), Vaccinium myrtillus (до 5 %). В подлеске обычна Sorbus sibirica (V к.п., ПП до 15 %). Большинство остальных видов встречаются единично с ПП не более 1-2 %. Цикл представлен несколькими лесными формациями: Pineta sylvestris , Piceeta obovatae , Pineta sibirici , Lariceta sibirici , Betuleta pubescentis . Существенных различий в пространственной и видовой структуре сообществ между формациями нет. Однако в кедровниках - наиболее сукцессионно зрелых сообществах цикла - обычен Sphagnum girgensohnii (IV к.п., ПП до 5 %) и усиливается роль Vaccinium myrtillus (V к.п., ПП до 10 %). Древостои часто двухъярусные; основу первого яруса древостоя составляют лиственница или кедр; во втором ярусе доминирует ель, реже - кедр. Подрост обычно обилен (до 29 тыс./га): кедр, ель, но представлен преимущественно особями младших генераций; разновозрастные елово-кедровые насаждения (в отличие от пойм некоторых других рек региона) не формируются.
Кустарничково-зеленомошный цикл ( Fruticu-loso-hylocomiosum ) приурочен к практически непоемным поверхностям старой поймы высокого уровня. Почвенный профиль несёт в себе явные следы аллювиального происхождения (слоистость и следы переменного увлажнения), но уже интенсивно идут процессы формирования оторфованной подстилки (мощность до 15 см), оподзоливания и оглеения. Цикл характеризуется абсолютным доминированием кустарничков: Vaccinium vitis-idaea (ПП до 50 %), V. myrtillus (ПП до 20 %), V. uligino-sum (ПП в среднем 3 %); с различным постоянством и обилием встречаются Ledum palustre , Em-petrum nigrum . Почва на 95 % покрыта зелеными мхами: Pleurozium schreberi (до 50 %), Hylocomium splendens (до 25 %), Dicranum polysetum (до 15 %); по микропонижениям - Sphagnum girgensohnii (до 20 %), по микроповышениям - Cladina stellaris (до 20 %). Подлесок не развит. Цикл представлен только сосновой формацией - Pineta sylvestris . Древостой разрежен (сомкнутость в среднем около 50 %).
Подрост немногочисленный (до 1,5 тыс./га - кедр, ель, сосна, береза).
Кроме рассмотренных выше довольно распространенных типов лесных сообществ, изредка встречаются аллювиофобные циклы: кустарничково-сфагновый цикл - Fruticuloso-sphagnosum , вейниковый цикл на естественно осушенных пойменных торфяниках - Calamagrostidoso purpurea turfo-sum (представленный только Betuleta pubescentis ) и др. Особый интерес представляет нетипичный для пойм лишайниковый цикл ( Cladinosum ) - на высоких гривах ляминской поймы. Эти лишайниковые сосняки, по мнению И.С. Ильиной, «ничем не отличаются от аналогичных водораздельных сообществ» [7]. Однако следует отметить, что по структуре почвенного профиля, производительности древостоя и по некоторым элементам флористической композиции пойменные и водораздельные сосняки-беломошники различаются.
Бедность и однообразие аллювиальных почв поймы р. Лямин является причиной крайне низкого флористического и синтаксономического разнообразия, по сравнению с поймами других рек региона и даже с водораздельными территориями. Индикатором этой бедности является отсутствие сообществ кислично-зеленомошного цикла и пихтовой формации ( Abieteta sibirici ). За три сезона полевых исследований в пойме Лямина лишь единственный раз был отмечен самосев пихты ( Abies sibirica ), но ни разу не встретились ни Oxalis acetosella , ни типичные ее региональные спутники - Aconitum sep-tentrionale , Actaea erythrocarpa , Atragene sibirica , Goodyera repens , Stellaria bungeana , исключительно редки и неактивны Gymnocarpium dryopteris , Rhytidiadelphus triquetrus . В то же время все эти виды достаточно часто встречаются на правом коренном берегу р. Лямин, а пихта местами является содоминантом древесного яруса внепойменных сообществ.
Всего в пойме р. Лямин выявлено 16 лесных ассоциаций, которые относятся к шести формациям. Первичные березняки характерны для молодой поймы, но встречаются и вторичные - на гарях зрелой поймы и на спущенных (естественно осушенных) торфяниках старой поймы. Осинники занимают крайне незначительные площади, обычно встречаясь на участках длительного интенсивного «омоложения» наносов. Лиственничники в пойме встречаются спорадически (по-видимому, все они пирогенного происхождения); ельники - очень редки. Можно предположить, что лимитирующим фактором развития темнохвойных лесов являются не только крайне бедные песчаные почвы, но и частые пожары, способствующие поддержанию длительнопроизводных сосновых лесов. Основной облик поймы р. Лямин формирует сосна - в отличие от рек Вахского полесья, где роль сосны в серийных сообществах пойм значительно меньше, чем березы, лиственницы, кедра и ели [7; 11 и др.]. Кедрачи занимают меньше 15% площади поймен- ных лесов. Но именно кедрачи-зеленомошники являются эдафическим субклимаксом в пойме р. Лямин – как и в поймах других рек Сургутского и Вах-ского полесий.
Список литературы Синтаксономическое разнообразие и экотопы лесных сообществ поймы реки Лямин (Сургутское Полесье)
- Атлас Ханты-Мансийского автономного округа -Югры. Т. II. Природа. Экология. Ханты-Мансийск-М., 2004. -152 с.
- Бабюк С.И., Кукуричкин Г.М. К характеристике почв поймы реки Лямин//Биологические ресурсы и природопользование: Сб. науч. тр. Вып. 11. Сургут: Дефис, 2008. С. 17-30.
- Земцов АА. Геоморфология Западно-Сибирской равнины (северная и центральная части). Томск, 1976. 344 с.
- Карта растительности Западно-Сибирской равнины. М. 1:1500000. М., 1976. 4 л.
- Кукуричкин Г.М. Морфология ландшафта поймы реки Глубокий Сабун//Экологические исследования восточной части Сибирских Увалов: Сб. науч. тр. Вып. 2. Нижневартовск: Приобье, 2003. С. 43-59.
- Кукуричкин Г.М. Циклы ассоциаций лесной растительности поймы реки Сабун//Экологический вестник Югории. 2004. Т. 1. № 3-4. С. 11-16.
- Кукуричкин Г.М. Темнохвойные леса в пойме реки Сабун//Актуальные проблемы геоботаники. Ч. I: материалы III Всерос. школы-конф. Петрозаводск: Кар НЦ РАН, 2007. С. 268-272.
- Лёзин В.А. Реки Ханты-Мансийского автономного округа: Справ. пособ. Тюмень: Вектор-Бук, 1999. 160 с.
- Полевая геоботаника. В 3 Т. М.-Л.: Наука, 1959-1976.
- Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. Экологическая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз, 1956. 472 с.
- Растительный покров Западно-Сибирской равнины. Новосибирск, 1985. 251 с.
- Шубаев Л.П. Сургутское Полесье Западно-Сибирской низменности//Изв. ВГО ВГО СССР. 1956. Т. 88, вып. 2. С. 167-169.
