Система генетических детерминант продуктивной кустистости мягкой озимой пшеницы

Автор: Мухордова Мария Евгеньевна
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 3 (23), 2016 года.
Бесплатный доступ
На основе диаллельных скрещиваний изучалась изменчивость и система генетических параметров в детерминации продуктивной кустистости у гибридов F1 мягкой озимой пшеницы. Из всех известных типов скрещивания общепринятым и наиболее информативным для генетического анализа количественных признаков растений является диаллельный. При подобных скрещиваниях исследователь располагает полным набором всех комбинаций генов, которыми обладают родительские сорта, а трудности, возникающие при их осуществлении, компенсируются богатством исходного материала для селекции. В полевых условиях 2013-2014 гг. на базе ФГБНУ «СибНИИСХ» г. Омска был заложен опыт. В статье изложен материал по изучению 5 сортов и 1 линии отечественной и зарубежной селекции, а также их диаллельных гибридов. Озимая пшеница - культура высокого плодородия. Реализовать свой биологический более высокий потенциал урожайности она может только при наличии генетической защиты от абиотических и биотических стрессоров и соответствующих условий выращивания. Генетический контроль рассматриваемого признака преимущественно определяется аддитивно-доминантной системой генов. Лабильность ее приводит к тому, что под влиянием условий среды может иметь место переопределение генетической формулы (набора генов) продуктивной кустистости. Вследствие нестабильности генетической системы контроля признака изменение его величины может управляться как доминантными, так и рецессивными генами. Отсюда следует, что отбор уникальных генотипов значительно затрудняется в связи со сменой сочетания метеофакторов. Несмотря на прогнозируемые трудности, отбор по признаку продуктивного стеблестоя следует начинать в ранних гибридных поколениях (с F2). В качестве доноров могут быть использованы сорта Минская и Заларинка в условиях переувлажнения, а при засухе - сорт Сплав.
Диаллельные гибриды, изменчивость, наследуемость, комбинационная способность, продуктивная кустистость, мягкая озимая пшеница
Короткий адрес: https://sciup.org/142199218
IDR: 142199218 | УДК: 631.524:633.11163249
Tilling capacity system of genetic determinants for soft winter wheat
Variability and genetic system determinants the tilling capacity from hybrids F1 of soft winter wheat in diallelic crossings was studied. The most informative for genetic of quantitative traits in plants is diallelic analysis all known types. The researcher has a complex of all genes combinations possessed by the parental forms at such way a crossings. The difficulties in their implementation are compensated by the diversity for hybrid material. The experiment was conducted in Siberian Scientific Research Institute of Agriculture (Omsk) in 2013-2014. The article presents the results of the studding 5 varieties and 1 line of native and foreign origin and their diallelic hybrids. Winter wheat is a crop which required high fertility soil. High biological potential yield it can be realized only when there is genetic protection against abiotic and biotic stressors and appropriate growing conditions. Genetic control of the tilling capacity trait is determined mainly by additive-dominant genes system. That is leads to the overriding genetic formulas (set of genes) of productive tilling capacity under the environmental conditions. The genetic instability of the control system of this trait operates by both dominant and recessive genes. So the selection of unique genotypes are greatly make difficulties, because of variability of meteorological factors. Despite of all difficulties, the selection on tilling capacity trait should begin in early hybrid generations (from F2). Varieties of winter wheat Minskaya and Zalarinka for wetting conditions, and variety Splav for drought conditions are offered as the donors.
Текст научной статьи Система генетических детерминант продуктивной кустистости мягкой озимой пшеницы
В настоящее время перед селекционерами стоит сложная задача – не только повысить урожайность пшеницы, но и сочетать ее с адаптивностью к различным почвенно-климатическим условиям региона, которые будут способствовать экологической безопасности, эффективно использовать природные и техногенные ресурсы, тем самым обеспечивать энергосбережение и рентабельность [1].
Озимая пшеница – культура высокого плодородия. Реализовать свой биологический более высокий потенциал урожайности она может только при наличии генетической защиты от абиотических и биотических стрессоров и соответствующих условий выращивания. При хорошей перезимовке в засушливые годы озимая пшеница является гарантированной культурой, благодаря способности использовать в экстремальных условиях влагу из более глубоких слоев почвы [2].
Количество стеблей, которое образует типичное растение у зерновых культур, – один из экологических признаков, значительно изменяющихся в связи с климатическими условиями (почвенная влага и температурный режим), применяемой агротехникой и наследственными особенностями генотипа. Продуктивная кустистость, как правило, в 1,5–2,0 раза меньше общей. Общая, и особенно продуктивная кустистость озимой пшеницы, во многом определяется биологическими особенностями сорта.
Устойчивость озимой пшеницы к отрицательным температурам во время перезимовки в значительной мере зависит от степени развитости растений, условий, сопровождающих закалку, влажности верхнего слоя почвы и других факторов. Наибольшую устойчивость к низким отрицательным температурам она приобретает в фазе кущения, когда имеется 2–4 побега [3, 4].
Сорта озимой пшеницы, отличающиеся большей кустистостью, обычно и более пластичны.
В опытах в условиях лесостепи Среднего Поволжья [5] выявлены корреляционные связи между коэффициентом кущения и зимостойкостью, глубиной залегания узла кущения и зимостойкостью, зимостойкостью и урожайностью. В данном эксперименте наблюдалась тесная связь между перезимовкой и коэффициентом кущения в 2006 г. ( r = 0,801), когда растения с осени хорошо раскустились, и слабая – в 2007 г. ( r = 0,231). В 2005 г. связь между перезимовкой и коэффициентом кущения не отмечена. Существенное влияние на перезимовку озимой пшеницы оказывала глубина залегания узла кущения.
Из всех известных типов скрещивания общепринятым и наиболее информативным для генетического анализа количественных признаков растений является диаллельный. При подобных скрещиваниях исследователь располагает полным набором всех комбинаций генов, которыми обладают родительские сорта, а трудности, возникающие при их осуществлении, компенсируются богатством исходного материала для селекции [6–18].
Цель работы – изучить изменчивость продуктивной кустистости мягкой озимой пшеницы и выявить системы генетического контроля в детерминации этого показателя.
Объекты и методы
Объект исследований – 5 сортов и 1 линия отечественной и зарубежной селекции (Жемчужина Поволжья, Юбилейная 180, Фантазия х (Донская остистая х Мутант 114) (далее Фантазия), Сплав, Минская, Заларинка), различающихся по ряду хозяйственно-ценных признаков; а также 30 диаллельных гибридов.
В 2013–2014 гг. в полевых условиях закладывались опыты. Высевались сорта и гибриды F 1 . Площадь питания растений 10 × 20 (см2). Повторность опыта трехкратная. Предшественник – кулисный пар.
Экспериментальный материал обработан методом дисперсионного анализа по Б.А. Дос-пехову [19], проведен генетический анализ по Акселю и Джонсу [20] в модификации Р.А. Цильке, Л.П. Присяжной [21].
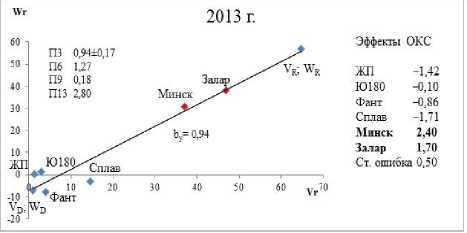
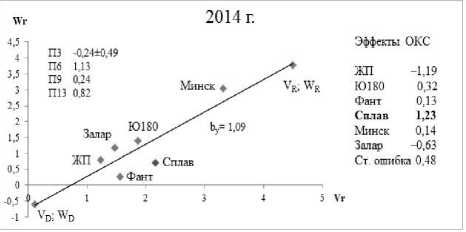
Генетику числа зерен в колосе озимой пшеницы изучали путем анализа графиков Хеймана [22] (зависимость W r от V r – коварианса и варианса) и генетических параметров: П3 – – (W r + V r ; X p ) – коэффициент корреляции между суммой W r + V r и с редним значением признака у родителей (мера направленности доминирования); П6 – √H1/D – мера средней степени доминирования внутри локусов в популяции; П9 – ¼ Н 2 /Н 1 – изме ряет среднее значение частот плюс и минус аллелей по всем локусам; П13 – √4DH1 + F/√4DH1 – F – характеризует отношение общего числа доминантных генов к общему числу рецессивных у родительских сортов. V D , W D ; V R , W R – координаты для полностью доминантного и рецессивного родителя.
На графике Хеймана связь между W r и V r выражается через коэффициент линейной регрессии b y . Эти параметры, которые дают относительно реальную ситуацию по организации количественного признака «число зерен в колосе», и использованы в наших исследованиях.
Комбинационную способность рассчитывали по В. Гриффингу [23, 24] модель I, метод I (в анализ включали данные о родителях, прямых и обратных гибридах).
По температурному режиму и количеству осадков условия вегетационного периода значительно различались по годам исследования.
Метеоусловия третьей декады августа 2012 г. (посев проведен 21.08.12) сложились благоприятно для получения дружных всходов и дальнейшего процесса кущения. Полевая всхожесть составила 78,2 %.
Характеризуя погодные условия зимнего периода 2013 г., можно сказать о том, что пониженные температуры декабря и двух декад января отрицательно сказались на перезимовке озимых культур, в частности мягкой озимой пшеницы. Количество осадков в декабрьский и февральский периоды также было пониженным.
При анализе гидротермического режима летних месяцев периода вегетации погода 2013 г. отмечена как прохладная и влажная (исключение – июнь).
Метеоусловия третьей декады августа 2013 г. (посев проведен 18.08.13) сложились благоприятно для получения дружных всходов и дальнейшего процесса кущения. Полевая всхожесть – 76,4 %.
В зимний период пониженные температуры были отмечены в третьей декаде января и первой февраля. Но они не повлияли отрицательным образом на перезимовку озимых культур, в частности мягкой озимой пшеницы, поскольку количество осадков в декабрьский и январский периоды было достаточным.
Гидротермический режим летних месяцев периода вегетации характеризует погоду 2014 г. как среднюю по температуре и засушливую.
Результаты исследований
В наших исследованиях продуктивная кустистость у сортов в среднем составила 15,4 шт. (табл. 1). В 2013 г. показатель продуктивного стеблестоя варьировал от 3,9 шт. (Жемчужина Поволжья) до 30,1 (Заларинка), а в 2014 г. – от 9,1 шт. (Заларинка) до 13,3 (Сплав).
У гибридов F 1 (табл. 1) продуктивная кустистость в среднем была ниже, чем у родительских форм. В 2013 г. она составила 14,9 стеблей: это меньше, чем у исходных форм на 5 стеблей. В 2014 г. эти показатели были равновеликими.
Таблица 1 Продуктивная кустистость, шт.
|
Сорт |
2013 г. |
2014 г. |
Среднее |
|||
|
P |
F 1 |
P |
F 1 |
P |
F 1 |
|
|
ЖП |
3,9 |
15,7 |
9,5 |
9,5 |
6,7 |
12,6 |
|
Ю180 |
17,5 |
14,1 |
12,3 |
9,8 |
14,9 |
11,9 |
|
Фантазия |
16,8 |
15,3 |
11,8 |
11,8 |
14,3 |
13,6 |
|
Сплав |
21,9 |
14,6 |
13,3 |
10,9 |
17,6 |
12,8 |
|
Минская |
29,3 |
15,1 |
9,5 |
11,9 |
19,4 |
13,5 |
|
Заларинка |
30,1 |
14,6 |
9,1 |
10,9 |
19,6 |
12,8 |
|
Среднее |
19,9 |
14,9 |
10,9 |
10,8 |
15,4 |
12,9 |
|
НСР 05 |
3,7 |
3,8 |
2,58 |
|||
На продуктивную кустистость большее влияние оказывал генотип, как подтверждает дисперсионный анализ в табл. 2, его доля составила почти 42 %. Чуть меньший эффект отмечен по условиям года – 37 %; на взаимодействие этих факторов приходится 21 %.
Полученная информация позволяет отметить, что данный показатель оказался наиболее стабильным по проявлению генотипических особенностей сортов и гибридов.
Таблица 2
Влияние факторов на изменчивость продуктивной кустистости
|
Фактор |
mS |
F ф |
F 05 |
% |
|
Генотип |
46,71* |
8,97 |
1,50 |
41,78 |
|
Условия года |
41,47* |
7,96 |
3,92 |
37,09 |
|
Взаимодействие |
23,62* |
4,53 |
1,50 |
21,09 |
|
Ошибка |
5,21 |
– |
– |
– |
* Достоверно при P ≤ 0,05.
Результаты расчетов комбинационной способности сортов по их гибридам (табл. 3) показали достоверность всех исследуемых факторов, а именно аддитивных эффектов генов, неаддитивных, а также ядерно-плазменных взаимодействий по годам исследования.
В прохладных и влажных условиях 2013 г. около 40 % вариации признака приходится на долю ОКС, что на 7 % меньше, чем на СКС. В 2014 г. (среднему по температурному режиму и засушливому) доля аддитивных эффектов генов была больше, чем в 2013 г. (53 %). На долю СКС приходилось около 31 %. Реципрокный эффект по годам исследования был равновеликим (13 и 15 % соответственно).
Таблица 3
Комбинационная способность сортов озимой пшеницы по продуктивной кустистости
|
Источник изменчивости |
2013 г. |
2014 г. |
||
|
mS |
% |
mS |
% |
|
|
ОКС |
34,52* |
39,55 |
8,34* |
53,35 |
|
СКС |
41,31* |
47,34 |
4,88* |
31,24 |
|
Р.Э. |
11,44* |
13,11 |
2,41* |
15,41 |
* Достоверно при P ≤ 0,05.
Соотношение варианс ОКС свидетельствует: в 2013 г. доля неаддитивных генов выше, чем в 2014 г. Это объясняется тем, что лимитирующим фактором в условиях южной лесостепи Западной Сибири является дефицит тепла, поэтому в неблагоприятный период возрастает вклад межаллельного взаимодействия.
Положительные эффекты ОКС отмечены у сортов Минская и Заларинка (рисунок) в 2013 г., в 2014 г. сорт Владимирской селекции Сплав внес положительный вклад в данный признак.
Генетика признака «продуктивная кустистость» сортов:
ЖП – Жемчужина Поволжья; Ю180 – Юбилейная 180; Фант – Фантазия;
Сплав; Минск – Минская; Залар – Заларинка
Анализ генетических параметров и графиков Хеймана позволяет отметить, что продуктивную кустистость увеличивают рецессивные гены в 2013 г., а в 2014 г. – доминантные. Это можно понять исходя из параметра П3, у которого разные направленности соответственно.
Линия регрессии W r /V r пересекает ось ординат с отрицательной стороны. Это указывает на присутствие в детерминации признака внутрилокусного сверхдоминирования. Наличие данного взаимодействия генов подтверждает и показатель средней степени доминирования, величина которого больше единицы (П6 1,27 и 1,13 соответственно по годам исследования).
В локусах, проявляющих доминирование, произведение частот плюс и минус аллелей асимметрично, как в 2013 г. (П9 = 0,18), так и в 2014 г. (П9 = 0,24), на что указывает отношение Н 2 /4Н 1 , которое не равно 0,25. Отношение общего числа доминантных генов у родительских сортов к общему числу рецессивных генов свидетельствует о превалирование первых (П13 = 3,45) в 2013 г.; в 2014 г. (П13 = 2,50).
Анализ расположения точек сортов на графике (вдоль линии регрессии) показывает их перемещение. Сорта, участвующие в опыте, проявляют относительную стабильность по годам исследования, располагаясь в одной и той же зоне на графиках Хеймана. Исключение – сорт Заларинка, в 2014 г. он переместился в зону доминантности.
Закл ючение
Генетический контроль рассматриваемого признака преимущественно определяется аддитивно-доминантной системой генов. Лабильность ее приводит к тому, что под влиянием условий среды может иметь место переопределение генетической формулы (набора генов) продуктивной кустистости. Вследствие нестабильности генетической системы контроля признака изменение его величины может управляться как доминантными, так и рецессивными генами. Отсюда следует, что отбор уникальных генотипов значительно затрудняется в связи со сменой сочетания метеофакторов. Несмотря на прогнозируемые трудности, отбор по признаку продуктивного стеблестоя следует начинать в ранних гибридных поколениях (с F 2 ). В качестве доноров могут быть использованы сорта Минская и Заларинка в условиях переувлажнения, а при засухе – сорт Сплав.
Список литературы Система генетических детерминант продуктивной кустистости мягкой озимой пшеницы
- Охременко А.В. Оценка коллекционных образцов озимой мягкой пшеницы и выделение источников ценных признаков для селекции в Центральном Предкавказье: дис. … канд. с.-х. наук. Ставрополь, 2016.
- Сандухадзе Б.И., Рыбакова М.И., Морозова З.И. Научные основы селекции озимой пшеницы в Нечерноземной зоне России. М., 2003. 426 с.
- Агробиологические основы производства, хранения и переработки продукции растениеводства/под ред. В.И. Филатова. М., 2003. 724 с.
- Пруцков Ф.М. Озимая пшеница. М., 1976. 352 с.
- Кривобочек В.Г., Косенко С.В. Результаты оценки исходного материала на зимостойкость в условиях лесостепи Среднего Поволжья//Вестн. Саратовского аграр. ун-та. 2011. № 2. С. 15-18.
- Михальцова М.Е., Калашник Н.А. Изменчивость и генетический контроль озерненности колоса у растений пивоваренного ячменя//Сельскохозяйственная биология. 2004. № 3. С. 72-75.
- Калашник Н.А., Козлова Г.Я., Аниськов Н.И. Генетика продуктивности и качества зерна пивоваренного ячменя в условиях Среднего Прииртышья. Новосибирск, 2005. 132 с.
- Гамзикова О.И., Калашник Н.А. Генетика признаков пшеницы на фонах питания. Новосибирск, 1988. 129 с.
- Генетика признаков продуктивности яровых пшениц в Западной Сибири/Драгавцев В.А., Цильке Р.А., Рейтер Б.Г. и др. Новосибирск, 1984. 232 с.
- Meirmans P.G. and Van Tienderen P.H. The effects of inheritance in tetraploids on genetic diversity and population divergence. Heredity, 2013, 110: 131-137; DOI: 10.1038/hdy.2012.80
- Special issues on advances in quantitative genetics: introduction/B. Walsh. Heredity, 2014, 112: 1-3; DOI: 10.1038/hdy.2013.115
- Anderson J.T., Wagner M.R., Rushworth C.A., Prasad K.V.S.K. and Mitchell-Olds T. The evolution of quantitative traits in complex. Heredity, 2014, 112: 4-12; DOI: 10.1038/hdy.2013.33
- Shaw R.G. and Shaw F.H. Quantitative genetic study of the adaptive process. Heredity, 2014, 112: 13-20; DOI: 10.1038/hdy.2013.42
- Aguirre J.D., Hine E., McGuigan K. and Blows M.W. Comparing G: multivariate analysis of genetic variation in multiple populations. Heredity, 2014, 112: 21-29; DOI: 10.1038/hdy.2013.12
- Wallace J.G., Larsson S.J. and Buckler E.S. Entering the second century of maize quantitative genetics. Heredity, 2014, 112: 30-38; DOI: 10.1038/hdy.2013.6
- Bijma P. The quantitative genetics of indirect genetic effects: a selective review of modelling issues. Heredity, 2014, 112: 61-69; DOI: 10.1038/hdy.2013.15
- Gholizadeha A., Dehghania H., Dvorak J. Interrelationships between seed yield and related traits in bread wheat genotypes in nonsaline and saline environments. Scientific Journal of Crop Science, 2014, 3(4): 24-31 DOI: 10.14196/sjcs.v3i4.1306
- Nietlisbach P., Keller L.F. and Postma E. Genetic variance components and heritability of multiallelic heterozygosity under inbreeding. Heredity, 2016, 116: 1-11; DOI: 10.1038/hdy.2015.59
- Доспехов Б.А. Методика полевого опыта. М., 1979. Aksel R., Johnson L. Anallysis of diallel cross: a work example. Advancing Frontiers of Plant Sciences; V.2: Ed. Radhu vira. Inst. for Advance of Sciences and Culture. India, New Delhi, 1963, V. 16: 37-53.
- Цильке Р.А., Присяжная Л.П. Методика диаллельного анализа исходного материала по количественным признакам: метод. рекомендации. Новосибирск, 1979.
- Hayman B. The analysis of variance diallel tables. Biometrics, 1954, 10: 235-244.
- Griffing B. Analysis of quantitative gene action by constant parent regression and related techniques. Genetics, 1950, 35: 303-312.
- Griffing B. Concept of general and specific combining ability in relation to diallel crossing system. Austral. J. Biol. Sci, 1956, 9: 463-493.
