Скрининг гена альфа-субъединицы бензоат диоксигеназы в бактериальных ассоциациях, полученных в результате селекции на (хлор)ароматических соединениях
ароматических соединениях")
Автор: Егорова Дарья Олеговна, Пьянкова Анна Александровна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Генетика
Статья в выпуске: 4, 2019 года.
Бесплатный доступ
Показано, что в тотальной ДНК шести бактериальных ассоциаций, полученных в результате накопительного культивирования в присутствии бифенила/хлорированных бифенилов, обнаружены фрагменты гена benA, кодирующего альфа-субъединицу бензоат диоксигеназы. В результате филогенетического анализа нуклеотидных последовательностей амплифицированных фрагментов гена benA установлено, что большинство изученных фрагментов формируют кластеры с гомологичными генами представителей отдела/филума Proteobacteria, уровень сходства при этом составил 100%. Уровень сходства с benA-генами известных грамотрицательных штаммов-деструкторов составил 79.5-89.7%. В ассоциации CHN4 выявлен ген benA, формирующий единый кластер с геном benA штаммов рода Rhodococcus, и на 96% схожий с гомологичным геном штамма-деструктора R. jostii RHA1.
Полихлорированные бифенилы, бактериальные ассоциации, бензоат диоксигеназа, ген
Короткий адрес: https://sciup.org/147227113
IDR: 147227113 | УДК: 579.222 | DOI: 10.17072/1994-9952-2019-4-464-470
Alpha-subunit benzoate dioxygenase gene screening in bacterial associations obtained by selection on (chlorine) aromatic compounds
It was shown that fragments of the benA gene encoding the alpha subunit of benzoate dioxygenase were found in the total DNA of six bacterial associations obtained as a result of enrichment cultivation in the presence of biphenyl / chlorinated biphenyls. As a result of phylogenetic analysis of the nucleotide sequences of amplified benA gene fragments, it was found that most of the studied fragments form clusters with homologous genes of the department / phylum Proteobacteria, the similarity level was 100%. The level of similarity with benA genes of known gram-negative strains-destructors was 79.5 - 89.7%. The benA gene, which was found in the DNA of the CHN4 association, forms a single cluster with the benA gene of strains of the genus Rhodococcus, and was 96% similar to the homologous gene of the straindestructor R. jostii RHA1.
Текст научной статьи Скрининг гена альфа-субъединицы бензоат диоксигеназы в бактериальных ассоциациях, полученных в результате селекции на (хлор)ароматических соединениях
Бензойная кислота по химической структуре принадлежит к классу ароматических соединений, имеющих в своем составе одно ароматическое кольцо. В окружающей среде широко встречаются как замещенные бензойные кислоты, содержащие в молекуле в качестве заместителей гидрокси-, метокси- группы, атомы хлора, так и незамещенная бензойная кислота. Данные соединения являются компонентами метаболических путей растений и бактерий, в результате чего они попадают в почву, воду и донные отложения [Abd El-Mawla, Beerhues, 2002; Field, Sierra-Alvarez, 2008a; Soly-anikova et al., 2015].
Значительное количество бензойной кислоты, выявленной в объектах окружающей среды, имеет антропогенное происхождение. Бензойная кислота используется в качестве консерванта при производстве широкого круга продуктов питания, а также в составе препаратов противомикробного и фунгицидного действия. Кроме этого, бензойная кислота входит в состав лекарственных средств, направленных против кожных заболеваний грибкового и клещевого происхождения. В промышленности бензойная кислота используется как исходное сырье для синтеза широкого спектра химических соединений []. Стоит отметить, что при биотрансформации соединений группы стойких органических загрязнителей, та-
ких как полихлорированные бифенилы (ПХБ), также возможно образование бензойной/хлор-бензойной кислоты [Pieper, 2005; Field, Sierra-Alvarez, 2008b].
Основными биологическими агентами, осуществляющими разложение бензойной кислоты в природе, являются аэробные бактерии. Способность к трансформации замещенных и незамещенных бензойных кислот описана для представителей филумов Proteobacteria, Actinobacteria, Fermicutes [Pieper, 2005; Field, Sierra-Alvarez, 2008a, b]. Биоразложение бензойной кислоты начинается с окисления молекулы под действием фермента бензоат 1,2-диоксигеназы (БДО) [Parales, Resnick, 2006]. БДО (КФ 1.14.12.10) состоит из двух α- и двух β-субъединиц []. Показано, что субстратная специфичность обусловливается α-субъединицей БДО [Parales, Resnick, 2006; Solyanikova et al., 2015]. Анализ нуклеотидной последовательности гена benA, кодирующего α-субъединицу БДО, выявил существенные различия данного гена у грамположительных и грамотрицательных бактерий [Haddad, Eby, Neidle, 2001; Field, Sierra-Alvarez, 2008a; Solyanikova et al., 2015]. Высказано предположение, что эволюция гена benA у данных групп бактерий протекала независимо друг от друга, однако они имеют общего предка [Haddad, Eby, Neidle, 2001; Field, Sierra-Alvarez, 2008a].
Цель настоящей работы – изучить разнообразие гена benA , кодирующего α-субъединицу бензоат 1,2-диоксигеназы, в тотальной ДНК бактериальных ассоциаций, полученных в результате селекции почвенных микробиоценозов в присутствии бифенила/полихлорированных бифенилов.
Материалы и методы исследования
Бактериальные ассоциации
Бактериальные ассоциации CHN1, CHN2, CHN3, CHN4, CHN5, CHN6 были получены в результате накопительного культивирования в присутствии бифенила/ПХБ из почв, отобранных на территории ОАО «СВЗХ» (г. Чапаевск, Самарская обл., Россия), а PN1 и PN2 – в результате селекции бактериоценоза почв, отобранных на территории ОАО «Пермский завод смазок и СОЖ» (г. Пермь, Пермский край, Россия) с применением в качестве селективного фактора бифенила/ПХБ.
Культивирование бактериальных ассоциаций
Все бактериальные ассоциации, использованные в настоящем исследовании, поддерживаются в метаболически активном состоянии методом периодического культивирования в минеральной среде К1 состава (г/л): K 2 HPO 4 ×3H 2 O – 3.2, NaH 2 PO 4 ×2H 2 O – 0.4, (NH 4 ) 2 SO 4 – 0.5,
MgSO 4 ×7H 2 O – 0.15, Ca(NO 3 ) 2 – 0.01 с внесением в качестве селективного фактора бифенила в концентрации 1 г/л.
Тотальная ДНК
Тотальную ДНК из бактериальных ассоциаций выделяли с использованием коммерческого набора реактивов FastDNA Spin Kit for Soil (MP Biomedicals, USA). Концентрацию ДНК измеряли на приборе QubitTM Fluorometer (Invitrogen, США) с приминением реактивов производителя.
Амплификация гена benA, кордирующего α-субъединицу бензоат диоксигеназы
Амплификацию гена benA на матрице тотальной ДНК, выделенной из бактериальных ассоциаций, проводили с использованием бактериальных праймеров прямого benA-F [5’-GCCCACGAGAGCCAGATTCCC-3’] и обратного benA-R [5’-GGTGGCGGCGTAGTTCCAGTG-3’] [Baggi et al., 2008]. Праймеры подобраны к консервативному участку гена benA штамма Acineto-bacter baylyi ADP1, амплифицируемая область со 175 нуклеотида до 712 нуклеотида (размер фрагмента 521 пн) [Baggi et al., 2008]. ПЦР проводили в 25 мкл смеси, содержащей 1х буфер для Taq -полимеразы с MgCl 2 (Синтол, Россия), 0.25 мМ дНТФ, 0.5 мкМ каждого праймера, 2 ед. акт. Taq -полимеразы (Синтол, Россия) и 2 мкл ДНК матрицы. Амплификацию осуществляли на приборе С1000 Touch («Bio-Rad Laboratories», США) в режиме: начальный денатурирующий шаг при 95°С в течение 5 мин., далее 30 циклов: 40 сек. при 94°С, 50 сек. при 60°С с понижением при каждом шаге на 0.4°С, 1 мин. при 72°С, завершающий шаг 7 мин. при 72°.
Визуализация амплифицированных фрагментов ДНК
Продукты ПЦР разделяли электрофорезом в агарозном геле (концентрация агарозы 0.8%) в 1х Трис-боратном буфере (Thermo scientific, Литва) при напряжении 10V/см и визуализировали в проходящем УФ-свете с использованием системы Gel Doc XRtm («Bio-Rad Laboratories», США) после окрашивания в растворе бромистого этидия.
Определение нуклеотидных последовательностей амплифицированных фрагментов ДНК
Определение нуклеотидных последовательностей гена benA осуществляли на автоматическом секвенаторе Genetic Analyzer 3500xl (Applied Biosystems, США), с применением реактивов Big Dye Terminator Ready Reaction Kit v 3.1 (Applied Biosystems, США), согласно рекомендациям производителя, в молекулярно-генетической лабора- тории кафедры ботаники и генетики растений ПГНИУ.
Анализ нуклеотидных последовательностей амплифицированных фрагментов гена benA
Поиск гомологичных последовательностей был произведен по базе данных GenBank . С применением пакета программ Mega версия 7.0, выявленные по базе данных и секвенированные в настоящем исследовании нуклеотидные последовательности гена benA были выровнены с последующим расчетом их сходства, которое было отображено в виде графической модели эволюционного дерева.
Размещение нуклеотидных последовательностей в международных базах данных
Нуклеотидные последовательности фрагмента гена benA , полученные в настоящем исследовании, были депонированы в международной базе данных GenBank. Номера, присвоенные данным последовательностям, представлены в разделе «Результаты и обсуждение».
Результаты и их обсуждение
Бактериальные ассоциации, полученные в результате селекции на минеральной среде в условиях, когда источником углерода является труднодоступное ароматическое вещество (бифенил/поли-хлорированные бифенилы), характеризуются наличием определенного набора генов, обусловливающих способность членов ассоциации использовать данный источник углерода в качестве ростового субстрата. Одним из таких генов является ген benA, кодирующий α-субъединицу бензоат диоксигеназы.
На ДНК-матрице бактериальных ассоциаций CHN1, CHN2, CHN3, CHN4, CHN5, CНN6, PN1 и PN2 с помощью праймеров benA-F и benA-R были амплифицированы фрагменты гена benA . Со всех образцов тотальной ДНК были получены ПЦР-продукты ожидаемого размера – 500 п.н. (рис. 1).
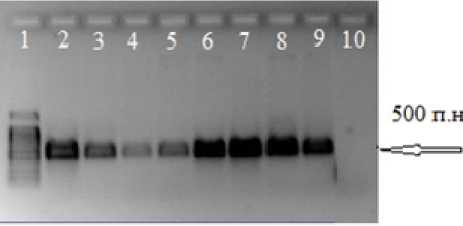
Рис. 1 . Электрофореграмма продуктов амплификации гена benA , кодирующего α-субъеди-ницу бензоат диоксигеназы:
1 – молекулярный маркер O'GeneRulerTM 100bp Plus DNA Ladder (Fermentas, Lithuania), 2 – CHN1, 3 – CHN2, 4 – CHN3, 5 – CHN4, 6 – CHN5, 7 – CHN6, 8 – PN1, 9 – PN2, 10 – отрицательный контроль
В результате проведения секвенирующей реакции и последующего секвенирования по Сэнгеру ампли-фицированных фрагментов, были получены хроматограммы, позволяющие осуществлять дальнейший анализ нуклеотидных последовательностей.
Проведены филогенетический и кластерный анализы полученных фрагментов гена benA с гомологичными последовательностями из базы данных GenBank (таблица, рис. 2).
Филогенетический анализ амплифицированных фрагментов гена benA из тотальной ДНК бактериальных ассоциаций
|
Ассоциация / номер в GenBank |
Штамм с наиболее близким геном benA , номер GenBank Штамм-деструктор с наиболее близким геном benA , номер GenBank |
Сходство, % / Перекрывание, % |
Количество проанализированных нуклеотидов |
|
CHN1 / MN396893 |
Pseudomonas putida JBC17 (CP029693.1) Pseudomonas putida KT2440 (LT799039.1) |
100 / 100 89.7 / 96 |
540 |
|
CHN2 / MN396894 |
Ralstonia euthropha JMP134 (CP000091.1) |
82.3 / 100 |
485 |
|
CHN3 / MN396895 |
Methylobacterium currus PR1016A (CP028843.1) Ralstonia euthropha JMP134 (CP000091.1) |
100 / 100 83.3 / 99 |
517 |
|
CHN4 / MN396896 |
Rhodococcus sp. S2-17 (CP021354.1) Rhodococcus jostii RHA1 (CP000431.1) |
100 / 100 96 / 100 |
476 |
|
CHN5 / MN396897 |
Ralstonia solanacearum SL3022 (CP023016.1) Ralstonia euthropha JMP134 (CP000091.1) |
100 / 99 88.8 / 99 |
493 |
|
CHN6 / MN396898 |
Burkholderia gladioli Co14 (CP033431.1) Ralstonia euthropha JMP134 (CP000091.1) |
100 / 100 79.5 / 98 |
498 |
|
PN1 / MN396900 |
Pseudomonas stutzeri CCUG 29243 (CP003677.1) Pseudomonas putida KF715 (AP015029.1) |
100 / 100 82.5 / 100 |
491 |
|
PN2 / MN396899 |
Pseudomonas putida JBC17 (CP029693.1) Pseudomonas knackmussii B13 (HG322950.1) |
100 / 100 89.5 / 100 |
524 |
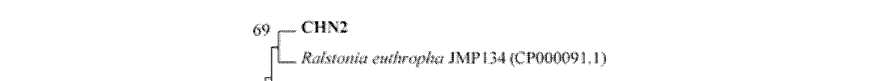

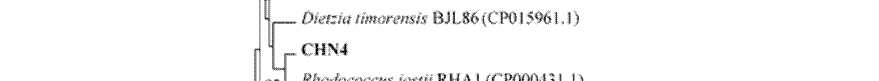
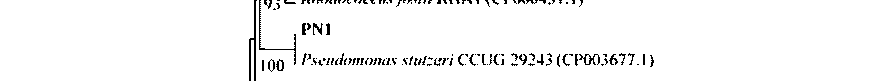


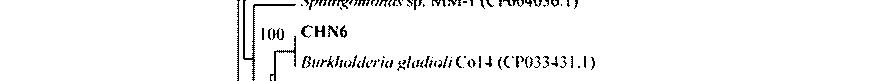

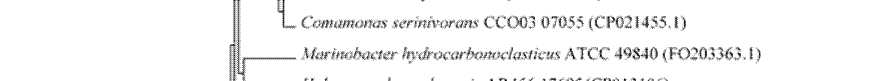
Сот.ишпшх чтатгар ( ( 003 (17055 ICP021455 I)
Marinч:кп ter iiydnivarbonoclnxtivus Al ((' 40X40 (H >20 3363. !)
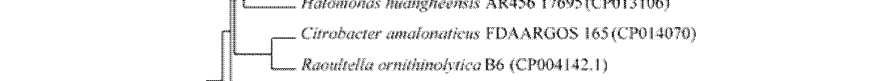

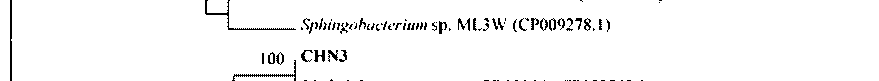

L PN2
—t-----------1-----------1-----------1-----------1-----------1-----------(
0 6 0,5 0.4 0.5 0.2 ill 0 0
Рис. 2. Дерево сходства выявленных генов с известными генами α-субъединицы бензоат 1,2-ди-оксигеназы, построенное методом UPGMA.
Масштаб соответствует 10 нуклеотидным заменам на каждые 100 нуклеотидов. «Bootstrap»-анализ проведен на 1000 повторностях. Значения рядом с «ветвями» показывают вероятность расположения последовательностей в данных группах. Жирным шрифтом выделены нуклеотидные последовательности, исследуемые в настоящей работе
Выявлен высокий уровень сходства амплифи-цированных участков гена benA исследуемых ассоциаций (100%) с генами подсемейства бензоат диоксигеназ бактерий различных таксономических групп, осуществляющих деструкцию ароматических соединений (таблица). Установлено, что в
ДНК исследуемых ассоциаций преимущественно амплифицировались гены бензоат диоксигеназ, характерных для бактерий отдела/филума Proteobacteria . Наиболее филогенетически близким среди генов бензоат диоксигеназы активных штаммов-деструкторов ароматических соединений для фрагментов гена benA из ассоциаций CHN2, CHN3, CHN5 и CHN6 является соответствующий ген штамма Ralstonia euthropha JMP134 (уровень сходства 79.5–88.8%). Известно, что штамм R. euthropha JMP134 осуществляет разложение широкого спектра ароматических соединений, в том числе бензойной и хлорбензойных кислот [Field, Sierra-Alvarez, 2008а]. Стоит отметить, что для фрагмента гена benA ассоциации CHN1 наиболее близким является ген бензоат диоксигеназы другого известного штамма-деструктора Pseudomonas putida KT2440 (таблица). Штамм P. putida KT2440 осуществляет разложение ароматических соединений и является одним из модельных штаммов, при анализе генетических систем [Kahlon R.S., 2016].
Фрагмент гена benA ассоциации CHN4, формирует единый кластер с соответствующим геном штаммов рода Rhodococcus класса Actinobacteria (таблица, рис. 2). Стоит отметить, что benA CHN4 имеет высокий уровень сходства (96%) с соответствующим геном известного штамма-деструктора бифенила/ПХБ Rhodococcus jostii RHA1 [Kitagawa et al., 2001].
Таким образом, в ассоциациях, селектированных из почв, отобранных на территории ОАО «СВЗХ» (г. Чапаевск, Самарская обл., Россия), выявлены фрагменты гена бензоат диоксигеназы, обладающие высоким уровнем сходства с benA -генами штаммов разных таксономических групп.
Иная картина получена при анализе нуклеотидных последовательностей фрагментов гена бензоат диоксигеназы, амплифицированных с тотальной ДНК ассоциаций PN1 и PN2, селектированных из почв ОАО “Пермский завод смазок и СОЖ” (г. Пермь, Пермский край, Россия). Наиболее близкими в данном случае по benA гену являются штаммы рода Pseudomonas, в том числе деструкторы Pseudomonas putida KF715 и Pseudomonas knackmussii B13 (уровни сходства 82.5 и 89.5%, соответственно). Штамм P. putida KF715 является высокоактивным деструктором хлорированных бифенилов, содержащих различное количества атомов хлора в молекуле бифенила, а также смесей ПХБ и обладает ферментативными путями, обусловливающими разложение бифенила до соединений основного обмена клетки [Suenaga et al., 2017; Kimura et al., 2018]. Штамм P. knackmussii B13 активно разлагает хлорбензойные кислоты, а в результате переноса ряда его генов в реципиент-ные штаммы получены трансконъюганты, способные использовать широкий спектр моно- и ди- хлорбензойных кислот [Kahlon, 2016]. Ранее было показано, что при культивировании почвенных микробиоценозов в присутствии бензола и 4-хлорбензойной кислоты в качестве селективных факторов, так же формируются сообщества, в которых присутствует бензоат диоксигеназа бактерий рода Pseudomonas [Назарова, Кирьянова, Егорова, 2019]. Стоит отметить, что кластерный анализ показал наличие существенных различий в нуклеотидных последовательностях выявленных генов benA ассоциаций PN1 и PN2 (рис. 2). Вероятно, ген бензоат диоксигеназы имеет значительную вариабельность в пределах рода Pseudomonas, что подтверждается формированием нескольких кластеров при графической визуализации результата анализа (рис. 2). Такое разнообразие может быть обусловлено как длительными эволюционными изменениями, так и адаптационными процессами, вызванными присутствием в окружающей среде химических поллютантов.
Заключение
В результате проведенных исследований изучены фрагменты гена benA , кодирующего α-субъ-единицу бензоат 1,2-диоксигеназы, присутствующие в тотальной ДНК восьми ассоциаций аэробных бактерий, селектированных на бифениле/по-лихлорированных бифенилах.
Показано, что амплифицированные участки гена benA имеют высокий уровень сходства (до 100%) с генами бензоат диоксигеназ штаммов, принадлежащих родам Burkholderia , Methylobacte-rium, Pseudomonas, Ralstonia и Rhodococcus . Установлено, что выявленные нуклеотидные последовательности были сходны на 79.5–96.0% с ben A-генами известных штаммов-деструкторов бензой-ных/хлорбензойных кислот и бифенила/поли-хлорированных бифенилов. Полученный результат позволяет высказать предположение, что в результате селекции в присутствии бифенила/ПХБ формируются бактериальные сообщества, обладающие набором генетических систем, обусловливающих биотрансформацию ароматических соединений.
Таким образом, в результате проведенных исследований изучено разнообразие гена benA , представленного в аэробных бактериальных ассоциациях, селектированных из почвенных микробиоценозов под воздействием бифенила/полихло-рированных бифенилов.
Работа выполнена при поддержке Российского фонда фундаментальных исследований, грант №18-29-05016мк.
Список литературы Скрининг гена альфа-субъединицы бензоат диоксигеназы в бактериальных ассоциациях, полученных в результате селекции на (хлор)ароматических соединениях
- Назарова Э.А., Кирьянова Т.Д., Егорова Д.О. Разнообразие гена бензоат диоксигеназы в бактериальных ассоциациях, сформировавшихся под давлением хлорорганического загрязнения // Экологическая генетика. 2019. Т. 17, № 3. С. 13-22. DOI: 10.17816/ecogen17313-22
- Abd El-Mawla A.M., Beerhues L. Benzoic acid biosynthesis in cell cultures of Hypericum androsaemum // Planta. 2002. Vol. 214. P.727. DOI: 10.1007/s004250100657
- Baggi G. et al. Co-metabolism of di- and trichlorobenzoates in a 2-chlorobenzoate-degrading bacterial culture: Effect of the position and number of halo-substituents // International Biodeterioration and Biodegradation. 2008. Vol. 62, № 1. P. 57-64. DOI: 10.1016/j.ibiod.2007.12.002
- Field J.A., Sierra-Alvarez R. Microbial transformation of chlorinated benzoates // Reviews in Environmental Science and BioTechnology. 2008a. Vol. 7. P. 191-210. DOI: 10.1007/s11157-0089133-z
- Field J.A., Sierra-Alvarez R. Microbial transformation and degradation of polychlorinated biphenyls // Environmental Pollution. 2008b. Vol. 155, № 1. P. 1-12.
- Haddad S., Eby D.M., Neidle E.L. Cloning and expression of the benzoate dioxygenase genes from Rhodococcus sp. strain 19070. // Applied Environmental Microbiology. 2001. Vol. 67. P. 2507-2514.
- Kahlon R.S. Pseudomonas: Molekular and Applied Biology. Springer International Publishing Switzerland, 2016. 519 p.
- DOI: 10.1007/978-3-319-31198-2_4
- Kimura N. et al. Pseudomonas furukawaii sp. nov., a polychlorinated biphenyl-degrading bacterium isolated from biphenyl-contaminated soil in Japan // International Journal Systematic Evolution Microbiology. 2018. Vol. 68, № 5. P. 1429-1435
- Kitagawa W. et al. Cloning and characterization of benzoate catabolic genes in the gram-positive polychlorinated biphenyl degrader Rhodococcus sp. strain RHA1 // Journal of Bacteriology. 2001. Vol. 183. P. 6598-6606.
- Parales R.E., Resnick S.M. Aromatic Ring Hydroxylating Dioxygenases // Pseudomonas / J.L. Ramos, R.C. Levesque, eds. Boston: Springer, 2006. P. 287-340.
- DOI: 10.1007/2F0-387-28881-3_9
- Pieper D.H. Aerobic degradation of polychlorinated biphenyls. // Applied Microbiology Biotechnology. 2005. Vol. 67, № 2. P. 170-191.
- DOI: 10.1007/s00253-004-1810-4
- Solyanikova I.P. et al. Pecularities of the degradation of benzoate and its chloro- and hydraxysubstituted analogs by Actinobacteria. // International Biodeterioration and Biodegradation. 2015. Vol. 100. P. 155-164.
- DOI: 10.1016/j.ibiod.2015.02.028
- Suenaga H. et al. Insights into the genomic plasticity of Pseudomonas putida KF715, a strain with unique biphenyl-utilizing activity and genome instability properties // Environmenmental Microbiology Reports. 2017. Vol. 9, № 5. P. 589-598.
