Смена напочвенного покрова на старо- пахотных залежах Каргопольского района Архангельской области

Автор: Голубева Любовь Владимировна, Наквасина Елена Николаевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Сельскохозяйственные науки
Статья в выпуске: 6 (143), 2014 года.
Бесплатный доступ
Рассмотрена смена ценофлоры старопахотных залежей в средней подзоне тайги Архангельской области (Каргопольский район), характеризующейся высокоплодородными дерново-подзолистыми почвами, сформированными на карбонатной морене. Объектом исследования являются 15-130-летние залежи (27 ПП), вышедшие из активного сельскохозяйственного оборота. Для геоботанического описания использовались общепринятые методики. Исследования показали, что на старопахотных залежах на протяжении длительного периода (более 100 лет) происходит смена ценофлоры или вторичная сукцессия, которая имеет определенные этапы сукцессионного перехода. На начальной стадии интенсивно разрастаются луговые травы и долгое время (до 40 лет) сохраняются виды, используемые в севообороте. Формируется Gramineae - Asteraceae - Rosaceae - Leguminosae ассоциация с наибольшим числом сегетальных растений в ее составе. Разнотравье представлено луговыми травами. В 40 лет происходит резкая смена фитоценоза, что доказывается при анализе сходства ценофлор. Формируется Asteraceae - Leguminosae ассоциация с включением лесных трав и появлением мохово-лишайникового подъяруса. Напочвенный покров состоит преимущественно из теневыносливых видов. Через 60 лет (после смыкания древостоя) в напочвенном покрове начинают преобладать лесные виды, происходит стабилизация лесной обстановки. В напочвенном покрове доминируют аборигенные виды, сегетальные вымещаются ближе к опушкам или исчезают совсем. Однако даже в спелом насаждении среди доминантов напочвенного покрова встречаются луговые виды трав.
Залежные поля, напочвенный покров, сукцессия, динамика, виды, сходство ценофлор
Короткий адрес: https://sciup.org/14750718
IDR: 14750718 | УДК: 630*913:581.93
Ground cover change on fallow fields of Kargopol district of Arkhangelsk region
The paper analyses old cultivated herbaceous vegetation deposits located in the middle taiga subzone of Arkhangelsk region (Kargopol district), which are characterized by extremely fertile sod podsol on the carbonate moraine base. While carrying out the research, ancient deposits (15-130 years old; sample plots - 27) no longer used for agricultural purposes were investigated. Conventional geobotanic description techniques were applied. The research revealed occurrences of the herbal phytocenosis rotation (secondary succession) with specific succession in transition stages. The change was observed in the old cultivated herbaceous vegetation deposits for a significant period of time (over 100 years). In the early stages, meadow grass grows intensively, and agricultural species used in the crop rotation survive up to 40 years. Simultaneously, Gramineae - Asteraceae - Rosaceae - Le-guminosae association is formed, its composition includes the largest number of weed species and forbs composed of meadow grass. Then, an abrupt phytocenosis change takes place after 40 year term of rotation. This conclusion is proven by the analysis of phytocenosis similarities. At this stage, Rosaceae - Leguminosae association including forest grass species and the moss lichen sublevel are formed. The canopy predominantly consists of the shade tolerant species. Furthermore, when the stand canopy closed (60 years later), the native forest species started to dominate in the ground cover. The forest conditions began to stabilize. However, the obtained results showed that meadow grass species invade dominating ground cover.
Текст научной статьи Смена напочвенного покрова на старо- пахотных залежах Каргопольского района Архангельской области
По данным ряда авторов, в Российской Федерации в настоящее время на площади более 10 млн га сельскохозяйственных угодий идет процесс зарастания древесной и кустарниковой растительностью. Эти процессы происходят в различных лесорастительных зонах, имеют принципиальные отличия и изучены недостаточно, особенно в отношении трансформации травяного покрова в процессе смены постагро-генной залежи лесом [3], [4], [5], [7], [8]. Вторичная сукцессия постагрогенных залежей происходит поэтапно, но в степной и таежной зонах разительно отличается. В условиях степной и лесостепной зон первые 2–3 года пашня зарастает одно- и двулетними видами, в последующие 5–7 лет – корневищными, которые затем сменяются рыхло-кустовыми и плотнокустовыми злаками [9; 180]. В подзоне южной тайги, где зарастание залежей происходит с участием древесных и кустарниковых пород, Н. Н. Новоселова по времени, прошедшему с момента прекращения использо-
вания, выделяет четыре стадии зарастающих угодий [7]. По ее данным, уже на четвертой стадии (15–20 лет) происходит формирование молодого сомкнутого леса (высота деревьев превышает 12 м) и появляются лесные виды трав при сохранении луговых.
Трансформация растительности на залежных полях более северных территорий до сих пор не изучена.
Наши исследования посвящены изучению напочвенного покрова постаграрных земель Каргопольского района, расположенного на юго-западе Архангельской области (подзона средней тайги). Одной из особенностей района являются почвообразующие породы, представленные карбонатной локальной мореной, распространенной в районах выхода известняков на поверхность. На карбонатной морене формируются высокопродуктивные дерново-карбонатные и дерновоподзолистые остаточно-карбонатные почвы, по свойствам существенно отличающиеся от зональных подзолистых.
История залежеобразования (отчуждения сельскохозяйственных земель из активного пользования) в районе позволила подобрать сукцессионный ряд пробных площадей различных сроков забрасывания (до 130 лет). Наиболее старые залежи образовывались в результате естественного процесса переселения жителей и утрат мелких деревень, позднее – банкротства сельскохозяйственных предприятий.
Геоботанические описания выполняли по общепринятой методике [6], изучение ценофлоры согласно [2]. Латинские названия растений напочвенного покрова даны по С. К. Черепанову [10]. Для оценки разнообразия видового состава использовали индексы: Шеннона, Жаккара, статистическую значимость средних значений показателей устанавливали, используя критерий Стьюдента. На основе индекса Шеннона вычисляли показатель выравненности [1].
Видовое разнообразие напочвенного покрова представлено более чем 20 семействами. В табл. 1 приведен список 10 ведущих семейств растений, формирующих растительные ценозы на старопахотных залежах разного возраста. Самые многочисленные из них: Gramineae Juss. (Poaceae Barnh.) (17 видов), Leguminosae Juss. (8 видов) и Asteraceae Dumort (13 видов). Остальные растительные семейства представлены меньшим числом видов.
Большинство многолетних трав семейства злаковых и бобовых (Dactylis glomerata L., Fes-tuca pratensis Huds., Festuca rubra L., Phleum pratense L., Vicia sativa L., Trifolium pratense L.) использовались в севооборотах, поэтому некоторые виды (Dactylis glomerata L., Trifolium pratense L.) спустя 25–40 лет продолжают доминировать и имеют среднее проективное покрытие 30–20 % и 10–15 % соответственно. На залежах 60–130 лет сохраняется доминирование вида только Trifolium pratense, среднее проективное покрытие которого на ПП варьирует от 30 до 25 %. Встречаются старопахотные 80–130-летние залежи с проективным покрытием Trifolium pratense до 80 % под пологом древостоя.
По мере сукцессионного развития залежей прежде всего резко уменьшается число видов семейства Gramineae: с 10–12 видов на ранних стадиях (в возрасте залежей до 40 лет) до 5 видов в возрасте залежей 80–130 лет, заросших лесом. Причем динамика смены неравномерна. В возрасте 40–60 лет залежеобразования происходит резкое снижение их количества, что подтверждается рассчитанным критерием Стьюдента (см. табл. 1). Между числом видов II–III сроков забрасывания различия статистически значимы (t005 = 2,6 при tst = 2,17). Далее численность видов в, напочвенном покрове остается достаточно стабильной, что связано с формированием на залежах лесной обстановки за счет смыкания крон деревьев.
При анализе ценофлоры все виды на старопахотных залежах были разделены на биогруппы, при этом выделяли: аборигенные (автохтонные), рудеральные, сегетальные, адвентивные (иммигранты) и виды-космополиты.
На всех залежах разных возрастов преобладают аборигенные (местные) виды, характерные для средней подзоны тайги (рис. 1). Число сегетальных ( Avena tatua L., Pimpinella saxifraga L., Achillea millefolium L. и др.) и рудеральных ( Arctium lappa L., Urtica dioica L.) видов сохраняется до 40 лет, затем постепенно сокращается. Из адвентивных видов встречается только Heracleum sosnowskyi Manden. на залежах 15– 25 лет. С течением времени он исчезает в зарослях и сохраняется только на открытых местах, лесных опушках, около лесовозных дорог. Неморальные виды на всей изученной территории отсутствуют.
Таблица 1
Динамика представленности видов ведущих семейств на залежах различных лет забрасывания
|
Семейство |
Среднее число видов, шт. |
Существенность различия (эмпирические значения критерия Стьюдента, tst) между группами залежей по возрасту |
|||||||
|
I* |
II |
III |
IV |
V |
I–II |
II–III |
III–IV |
IV–V |
|
|
Gramineae |
12 |
10 |
4 |
3 |
5 |
1,1 |
2,6 ** |
0,2 |
0,1 |
|
Asteraceae |
12 |
7 |
4 |
2 |
4 |
14,1 |
1,3 |
0,6 |
0,1 |
|
Rosaceae |
7 |
5 |
8 |
6 |
7 |
1,9 |
1,7 |
9,0 |
0,5 |
|
Leguminosae |
6 |
4 |
5 |
4 |
3 |
1,3 |
0,5 |
0,5 |
0,2 |
|
Umbelliferae |
4 |
3 |
2 |
1 |
2 |
0,6 |
0,5 |
0,2 |
0,1 |
|
Ranunculaceae |
3 |
2 |
1 |
4 |
3 |
0,4 |
0,3 |
1,4 |
0,8 |
|
Equisetaceae |
1 |
2 |
1 |
3 |
1 |
0,5 |
0,5 |
1,4 |
1,2 |
|
OrchIdaceae |
2 |
2 |
0 |
0 |
1 |
0,3 |
3,3 |
0,3 |
0,3 |
|
Pyrolaceae |
0 |
0 |
3 |
2 |
1 |
0,3 |
2,8 |
1,8 |
0,3 |
|
Liliaceae |
0 |
0 |
2 |
1 |
2 |
0,3 |
3,4 |
0,6 |
0,4 |
Примечание. * – длительность залежи: I – 15–25 лет, II – 25–40 лет, III – 40–60 лет, IV – 60–80 лет, V – 80–130 лет. ** – жирным шрифтом выделены показатели различия, достоверные при 5 % уровне значимости.
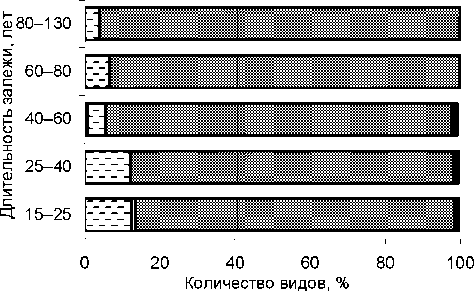
□ рудеральные и сегетальные
□ сегетальные растения
□ адвентивные (заносные)
S аборигенные ■ космополиты
Рис. 1. Представленность растений по биогруппам в динамике залежеобразования
Все виды растений были распределены на эко-лого-ценотические группы: лесные, луговые, комбинированные (растут на лугу и в лесу) (рис. 2). Залежные пахотные земли постепенно зарастают сначала луговыми травами ( Leucanthemum Hill vulgare Lam., Campanula patula L., Achillea L. s. l. millefolium L., Alchemilla glomerulans Bus., Briza media L., Centaurea scabiosa L., Campanula persicifolia L. и др.), затем появляются лесные ( Athyrium fi lix-femina (L.) Roth, Oxalis acetosella L., Maianthemum Wigg. bifoilium (L.) F. W. Schmidt, Pyrola rotundifolia L. и др.).
Луговые травы начинают терять свое преимущество в травостое уже через 25 лет зале-жеобразования. Достоверность различия по числу луговых трав между залежами I–II групп возраста доказана (t005 = 14,2 при tst = 1,9). Смена луговой ценофлоры н, а лесную, в состав которой включаются лесные виды трав ( Rubus saxatilis L., Trientalis europaea L., Paris quadrifolia L. и др.),
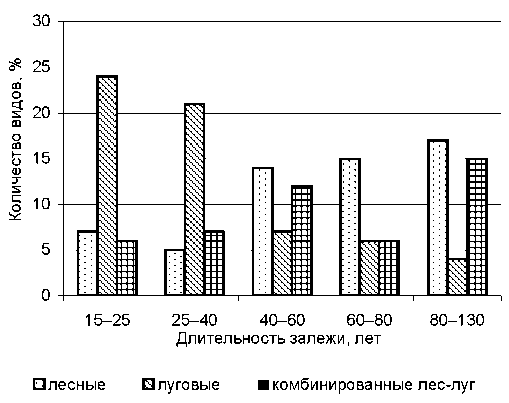
Рис. 2. Принадлежность растений к различным эколого-ценотическим группам происходит после 40 лет, при этом в напочвенном покрове долгое время сохраняются виды, применяемые в севообороте до забрасывания земель (Dactylis glomerata L. и Trifolium pratense L.). Идет формирование лесной обстановки под пологом древостоя. Различия доказываются критерием Стьюдента при 5 % значимости по всем представленным эколого-ценотическим группам: t005 = 3,01; 3,70; 3,00 при табличном tst = 2,05 (воз-ра, ст залежей III–IV, табл. 2).
Видовое разнообразие трав связано как с запасом семян в почвах залежей, так и с заносом их из окружающих ценозов. Залежи всех сроков забрасывания имеют высокое разнообразие трав, что доказывается информационной мерой разнообразия Шеннона (табл. 3), согласно которому разнообразие сообщества тем выше, чем больше в нем количество видов и выше его выравнен-ность [1]. Это подтверждается показателем вы-равненности, указывающим на численное обилие видов, который во всех случаях приближается к 1 (E = 0,8…0,9).
Таблица 2
Сравнение представленности видов различных ценофлор в динамике залежеобразования (критерий Стьюдента, 5 %)
|
Эколого-ценотическая группа |
Критерий Стьюдента |
|||||||
|
Эмпирические значения, tst |
Табличные значения, t0,05 |
|||||||
|
I*–II |
II–III |
III–IV |
IV–V |
I–II |
II–III |
III–IV |
IV–V |
|
|
Лесные |
1,11 |
0,4 |
3,01 |
-2,4 |
2,09 |
2,2 |
2,05 |
2,1 |
|
Луговые |
14,2 |
0,8 |
3,70 |
2,2 |
1,9 |
2,01 |
2,05 |
2,05 |
|
Комбинированные (лес – луг) |
0,66 |
0 |
3,00 |
-1,3 |
2,08 |
2,02 |
2,05 |
2,01 |
Примечание. * – длительность залежи: I – 15–25 лет, II – 25–40 лет, III – 40–60 лет, IV – 60–80 лет, V – 80–130 лет.
Таблица 3
Индекс Шеннона биоразнообразия напочвенного покрова (при t0,05=1,9)
|
Длительность залежи, лет (группа) |
Коэффициент Шеннона и стандартная ошибка, Н |
Показатель вырав-ненности коэффициента Шеннона, Е |
Коэффициент Стьюдента (t005) между группами залежей разного ,возраста |
|||
|
15–25 (I) |
3,90 ± 0,72 |
–0,90 |
10,9* |
|||
|
25–40 (II) |
3,41 ± 0,64 |
–0,86 |
0,07 |
|||
|
40–60 (III) |
3,20 ± 0,65 |
–0,82 |
1,14 |
|||
|
60–80 (IV) |
2,96 ± 0,52 |
–0,80 |
6,2 |
|||
|
80–130 (V) |
3,26 ± 0,63 |
–0,86 |
||||
Примечание. * – жирным шрифтом выделены различия индекса Шеннона, доказанные на 5 % уровне значимости.
Информационная мера разнообразия – показатель Шеннона – в природных популяциях обычно варьирует в пределах 1,5–3,5, редко превышая эту цифру [1]. В нашем случае на свежих залежах показатель превышает значения природных сообществ и достигает 3,90 ± 0,72. Позднее показатель Шеннона закономерно снижается, достигая наиболее низких значений в залежах 60–80 лет (Н = 2,96 ± 0,52). Наиболее сильно видовое разнообразие меняется в 15–25 лет, то есть на начальных стадиях сукцессии, и в 60–80 лет, когда происходит стабилизация лесной обстановки. Так, доказано различие (табл. 3) между показателями видового разнообразия залежей I и II сроков забрасывания (t005 = 10,9) и IV и V сроков забрасывания (t005 = 6,2), .
В первые год,ы залежеобразования происходит стабилизация ценоза, тогда как в 40–60 лет после забрасывания полей после пахоты – постепенная смена флористического состава (рис. 3), что подтверждается коэффициентом Жаккара, который показывает сходство видового состава сообществ. Высокий процент сходства установлен между относительно молодыми залежами (15–40 лет) (60 %), а также между старыми залежами (60–130 лет) (56 %). Снижение показателя сходства наблюдается в период залежеобразова-ния с 40 до 60 лет. Сходство видового состава между молодыми и старыми залежами (начало и конец сукцессии), согласно коэффициенту Жаккара, одинаково низкое и составляет 7 %.
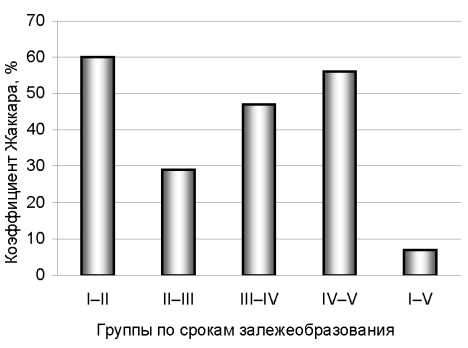
Рис. 3. Оценка видового сходства сообществ по срокам длительности залежи: I – 15–25 лет, II – 25–40 лет, III – 40–60 лет, IV – 60–80 лет, V – 80–130 лет
Таким образом, исследования показали, что на старопахотных залежах происходит смена цено-флоры, или вторичная сукцессия, которая имеет определенные этапы сукцессионного перехода.
В 15–25 лет после забрасывания сельскохозяйственных угодий на стадии залужения формируется Gramineae – Asteraceae – Rosaceae – Leguminosae ассоциация с наибольшим числом сегетальных растений в ее составе. Разнотравье представлено луговыми травами. Высота травос- тоя и повышенная плотность сухой ветоши мешают естественному возобновлению древесных пород. На пробных площадях в большей степени растут подлесочные виды вегетативного происхождения, такие как ивы.
В 25–40 лет стадия залужения продолжается с формированием Gramineae – Asteraceae – Rosaceae – Leguminosae ассоциации. В составе разнотравья все также присутствуют луговые светолюбивые виды трав. Высота травостоя сохраняется, что препятствует росту древесного яруса. Смыкания крон древесных пород не происходит. Между 15–40-летними залежами наблюдается высокий процент видового сходства, что объясняется высоким плодородием почвы, сохранившимся после сельскохозяйственного использования, наличием семян трав в почве и отсутствием явной конкуренции видов (большие площади залежных полей).
После 40 лет залежеобразования происходит резкая смена фитоценозов и формируется Rosaceae – Leguminosae ассоциация с включением лесных трав и появлением мохово-лишайникового подъяруса. Напочвенный покров состоит преимущественно из теневыносливых видов. В два раза снижается высота травостоя. Конкурентные отношения между травяным и древесным ярусами исчезают. Происходит смыкание крон древесных пород. Данный период можно считать переходным между луговым и лесным фитоценозом.
На залежах старше 60 лет под пологом древесного яруса формируется лесная обстановка, наступает стабилизация лесного напочвенного покрова. В составе травостоя доминируют аборигенные виды. Сегетальные виды вымещаются ближе к опушкам или выпадают совсем. Выпадения луговых трав полностью не происходит, но в их числе больше трав теневыносливых и комбинированных (растущих на лугу и в лесу). Древесный ярус полностью сформирован, кроны сомкнуты. Полная смена фитоценозов подтверждается низким коэффициентом Жаккара (7 %), между 15-летними и 80–130-летними залежами.
Залежи до 40-летнего срока забрасывания могут быть использованы как сенокосы, под лесокультурное или плантационное лесоразведение, а при необходимости и слабом зарастании древесными породами – восстановлены для активного сельскохозяйственного пользования (пашни). Однако возникнет проблема засоренности полей, подбора технологий создания лесных культур, возможно, подбора гербицидов. Залежи старшего возраста, заросшие лесом, потребуют лесоводственных уходов за древесным ярусом для формирования высокополнотных древостоев.
GROUND COVER CHANGE ON FALLOW FIELDS OF KARGOPOL DISTRICT OF ARKHANGELSK REGION
Список литературы Смена напочвенного покрова на старо- пахотных залежах Каргопольского района Архангельской области
- География и мониторинг биоразнообразия/Под ред. Н. С. Касимова. М., 2002. 432 с.
- Гнатюк Е. П., Крышень А. М. Методы исследования ценофлор (на примере растительных сообществ вырубок Карелии): Учебно-методическое пособие. Петрозаводск: Карельский научный центр РАН, 2005. 68 с.
- Горяинова И. Н., Леонова Н. Б., Феодоритов В. М. Процессы зарастания сельскохозяйственных земель в средней тайге Архангельской области//Вестник Московского университета. М., 2012. Вып. 3. С. 41-47.
- Люри Д. И., Горячкин С. В., Короваева Н. А. и др. Динамика сельскохозяйственных земель России в XX веке и постагрогенное восстановление растительности и почв. М.: ГЕОС, 2010. 426 с.
- Морозов А. М. Формирование насаждений на землях, исключенных из сельскохозяйственного оборота, в подзоне предлесостепных сосново-березовых лесов Свердловской области: Автореф. дисс.. канд. с.-х. наук. Екатеринбург: Уральский гос. лесотехнический ун-т, 2008. 20 с.
- Наквасина Е. Н., Шаврина Е. В. Геоботанические исследования. Архангельск: Поморский гос. ун-т, 2001. 43 с.
- Новоселова Н. Н. Формирование лесных насаждений на землях, вышедших из-под сельскохозяйственного использования, в таежной зоне Пермского края: Автореф. дисс.. канд. экон. наук. Екатеринбург, 2007. 20 с.
- Соколов Н. Н. Рост и продуктивность сосновых древостоев по старым пашням//Лесной журнал. 1978. Вып. 4. С. 22-25.
- Сукачев В. Н. Растительные сообщества (введение в фитоценологию). 4-е изд. Л.; М.: Книга, 1928. 232 с.
- Черепанов С. К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР). СПб.: Мир и семья, 1995. 992 с.
