Содержание и состав липидов в листьях растений на Южном Тимане (ботанический заказник «Сойвинский»)
")
Автор: Е.В. Силина, Г.Н. Табаленкова, Т.К. Головко
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Статья в выпуске: 7 (83), 2025 года.
Бесплатный доступ
Изучены содержание и состав липидов в листьях Aconitum septentrionale Koelle. (аконит высокий), Astragalus danicus Retz. (астрагал датский) и Medicago lupulina L. (люцерна хмелевидная) на Южном Тимане. Установлено, что содержание и состав липидов больше зависели от условий местообитания, чем от вида растения. Листья затененных растений накапливали в 1,5 раза больше липидов и характеризовались более низким вкладом фосфои нейтральных липидов в общий пул липидов по сравнению с растениями, произрастающими на освещенных участках. Доля насыщенных жирных кислот в листьях M. lupulina, предпочитающего открытые местообитания, была заметно выше, чем в листьях A. danicus, произрастающего в густом травостое. Полученные результаты свидетельствуют о значимости изучения содержания и состава липидов для понимания механизмов адаптации растений к условиям окружающей среды.
Общие липиды, фосфолипиды, нейтральные липиды, жирные кислоты, освещенность, листья, растения, Южный Тиман
Короткий адрес: https://sciup.org/149149450
IDR: 149149450 | УДК: 581.1:53.009:577.115:574.24 | DOI: 10.19110/1994-5655-2025-7-93-99
The content and composition of lipids in plant leaves in the Southern Timan (Soivinsky Botanical Reserve)
The paper is concerned with the content and composition of lipids in the leaves of Aconitum septentrionale Koelle (monkshood high), Astragalus danicus Retz. (danish astragalus), and Medicago lupulina L. (hop-shaped alfalfa) in the Southern Timan. The content and composition of lipids have been found to depend rather on the habitat conditions than on the plant species. Leaves of plants grown in shaded conditions accumulate by 1.5 time more lipids and have a lower contribution of phospho- and neutral lipids to the total lipid pool compared to plants grown in light areas. The proportion of saturated fatty acids in the the leaves of M. lupulina, which prefers open habitats, is noticeably higher than that in leaves of A. danicus, which grows in dense herbage. Our results indicate the importance of studying changes in the content and composition of lipids for understanding the mechanisms of plant adaptation to environmental conditions.
Текст научной статьи Содержание и состав липидов в листьях растений на Южном Тимане (ботанический заказник «Сойвинский»)
В настоящее время вопросы выявления, оценки и рационального использования растительных ресурсов приобретают особое значение. Для рационального использования ресурсов важно не только оценивать запасы сырья и содержание целевого продукта, необходимо более полное знание об их эколого-биологических и биохимических свойствах. Среди растений природной флоры таежной зоны Республики Коми большой интерес представляет Aconitum septentrionale Koelle (аконит высокий), Astragalus danicus Retz. (астрагал датский) и Medicago lupulina L. (люцерна хмелевидная).
Липиды – один из основных классов биомолекул, которые являются структурной основой клеточных мембран. Благодаря способности модулировать физические свой- ства мембран, выступать в роли сенсоров и сигнальных молекул, а также взаимодействовать с белками, влияя на их внутриклеточную локализацию и активность, липиды вносят существенный вклад в функционирование клеток [1-3]. Липиды участвуют в регуляции многих процессов, включая рост и деление клеток, мембранный транспорт, внутриклеточную сигнализацию и межклеточную коммуникацию, формирование ответных реакций на факторы среды [4]. Указанное многообразие функций липидов подчеркивает важность изучения данных соединений для характеристики свойств растений, изменения метаболизма в процессе их роста и/или при адаптации к условиям местообитания.
В данной работе приведены результаты изучения содержания липидов и жирных кислот в листьях трех видов многолетних травянистых растений, произрастающих на территории ботанического заказника «Сойвинский» (Южный Тиман).
Материалы и методы
Район исследований ( 62.74904° с. ш., 55.82610° в. д.) характеризуется умеренно-континентальным климатом с длинной, умеренно холодной зимой и коротким, умеренно теплым летом. Средняя температура самого теплого месяца – июля – +16–17° С. В период проведения исследований (первая декада июля) погода выдалась сравнительно прохладной с выпадением обильных осадков и высокой облачностью; среднесуточная температура была ниже нормы на 3–4° С.
Объектами исследования служили листья Aconitum septentrionale Koelle. (аконит высокий), Astragalus dani-cus Retz. (астрагал датский) и Medicago lupulina L. (люцерна хмелевидная). Растения A. septentrionale (сем. Ranunculaceae) произрастали в двух типичных для данного района cообществах на левом берегу р. Сойвы. Це-нопопуляция 1 (ЦП 1) охарактеризована как ельник круп-нотравно-злаковый, а ЦП 2 описана как сероольшанник аконитовый [5]. Растения ЦП 1 получали меньше света, чем растения ЦП 2. Растения A. danicus и M. lupulina (сем. Faboideae) обитали в разных по световому режиму условиях. Растения A. danicus произрастали в густом травостое на лугу, растения M. lupulina – на хорошо освещенном участке, вдоль дороги. Растения всех изученных видов находились в фазе цветения.
Образцы листьев отбирали в утренние часы, высушивали в сушильном шкафу при температуре +80° С до постоянного веса и хранили в бумажных пакетах в сухом месте до проведения анализов. Перед анализом пробы листьев измельчали до порошковидного состояния с помощью аналитической мельницы «IKA A 11 basic» (Япония).
Выделение общих липидов (ОЛ) проводили методом Фолча [6]. Общие липиды экстрагировали из сухой массы листьев смесью хлороформ-метанол (2 : 1) из расчета 20 частей экстрагирующей смеси на одну часть ткани. Экстракцию осуществляли трижды при постоянном перемешивании. Полученный экстракт упаривали на ротационном испарителе «Heidolph Hei VAP Value G3» (Россия), в вакууме досуха при температуре не выше +40° С в предварительно взвешенных круглодонных колбах. Сухой остаток взвешивали и рассчитывали выход ОЛ. Содержание ОЛ выражали в мг/г сухой массы.
Для выделения фосфолипидов (ФЛ) сухой остаток ОЛ растворяли в минимальном количестве хлороформа. К раствору добавляли охлажденный в морозильной камере ацетон до появления мутного осадка и оставляли в морозильной камере для оседания ФЛ. Затем смесь фильтровали через предварительно взвешенные бумажные фильтры, высушивали в сушильном шкафу и взвешивали. Содержание ФЛ выражали в мг/г сухой массы.
Для извлечения нейтральных липидов (НЛ) использовали неполярный растворитель – гексан. С целью наиболее полного перехода исследуемых липидов в органическую фазу навески высушенных образцов листьев подвергали трехкратной экстракции при постоянном перемешивании. Экстракт фильтровали и упаривали на ротационном испарителе «Heidolph Hei VAP Value G3» (Россия) в вакууме досуха при температуре не выше +40° С в предварительно взвешенных круглодонных колбах. Сухой остаток взвешивали, рассчитывали выход нейтральных липидов [7].
Определение жирнокислотного состава общих липидов проводили путем их метилирования по методу [8]. Жирные кислоты (ЖК) определяли в виде их метиловых эфиров (МЭЖК) после проведения кислотного метанолиза липидов с последующей этерификацией выделившихся кислот. Метиловые эфиры разделяли на газовом хроматографе «КРИСТАЛЛ 2000М» (Хроматэк) с пламенно-ионизационным детектором в изотермическом режиме температуры термостата колонок (200° C) на кварцевой капиллярной колонке 30 м × 0,2 мм, газ-носитель – гелий, чистота – 99,99 %. Анализ высших ЖК проводили в экоана-литической лаборатории Института биологии ФИЦ Коми НЦ УрО РАН. Регистрацию и обработку хроматограмм осуществляли с помощью системы сбора и обработки хроматографических данных «Хроматэк». Идентификацию и количественное определение МЭЖК проводили методом хромато-масс-спектрометрии на приборе «TRACE-DSQ» (Thermo Fisher Scientific, США) в режиме полного ионного тока (энергия электронов: 70 эВ). Сканирование МЭЖК осуществляли в диапазоне от 30 до 650 массовых чисел.
Статистическую обработку данных проводили с использованием программы Statistica 10 («StatSoft Inc.», США). Значимость различий между средними значениями измеряемых показателей оценивали с применением критерия Стьюдента. Расчеты осуществляли при заданном уровне значимости р ≤ 0,05. В таблицах и на рисунках приведены средние арифметические значения со стандартной ошибкой.
Результаты и их обсуждение
На рис. 1 представлены сравнительные данные о содержании липидов в листьях растений A. septentrionale из разных ценопопуляций. В ельнике крупнотравно-зла-ковом (ЦП 1) содержание липидов в сухой массе листьев A. septentrionale превышало 10 %, тогда как в серооль-шаннике (ЦП 2) величина этого показателя была достоверно ниже и составляла около 7 %. Растения из ЦП 1 и ЦП 2 не отличались достоверно по содержанию фосфо- и нейтральных липидов, на долю которых приходилось соответственно около 3 и 1 % сухой биомассы листьев. Однако относительное содержание ФЛ и НЛ (в % от ОЛ) было существенно выше в листьях растений из ЦП 2, чем в листьях растений из ЦП 1 (табл. 1). Вклад ФЛ и НЛ в пул ОЛ у листьев растений ЦП 2 составлял 39 и 13 %, а в листьях растений ЦП 1–27 и 11 % соответственно.
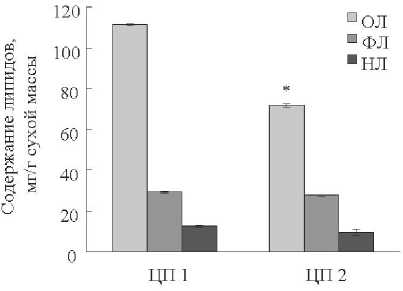
Рисунок 1. Содержание и состав липидов в листьях растений Aconitum septentrionale , произрастающих в ельнике крупнотравно-злаковом (ЦП 1) и сероольшаннике (ЦП 2).
Условные обозначения. Здесь и в рис. 2: ОЛ – общие липиды; ФЛ – фосфолипиды; НЛ – нейтральные липиды. * статистически значимые различия исследуемого показателя (р ≤ 0,05).
Figure 1. The content and composition of lipids in the leaves of Aconitum septentrionale plants growing in large-herb-cereal spruce forest (ЦП 1) and grey alder forest (ЦП 2).
Keys. Here and in Figure 2: ОЛ – total lipids; ФЛ – phospholipids; НЛ – neutral lipids. * statistically significant differences in the studied parameter (р ≤ 0.05).
Таблица 1
Относительное содержание нейтральных и фосфолипидов в листьях растений Aconitum septentrionale из разных ценопопуляций (% от суммы общих липидов)
Table 1
Relative content of neutral and phospholipids in the leaves of Aconitum septentrionale plants from different coenopopulations (as a percentage of the total lipid content)
|
Местообитания / Habitats |
Фосфолипиды |
Нейтральные липиды |
|
Ельник крупнотравно-злаковый |
27,1±1,0 |
11,2±1,0 |
|
Сероольшанник |
39,2±2,2* |
13,1±3,3 |
Условное обозначение. Здесь и в табл. 2: * статистически значимые различия исследуемого показателя (р ≤ 0,05).
Key. Here and in Table 2: * statistically significant differences in the studied parameter (р ≤ 0.05).
Изучение липидов у двух видов бобовых показало, что содержание липидов в сухой массе листьев растений A. danicus , обитающих на лугу в густом травостое составляло около 12 % сухой биомассы (рис. 2). Содержание ФЛ и НЛ в листьях A. danicus составляло соответственно около 3 и 1,5 % сухой биомассы. Содержание липидов в растениях M. lupulina , произрастающих вдоль дороги, на открытом участке составляло чуть более 7 % сухой биомассы листьев. Содержание ФЛ и НЛ в листьях A. danicus составляло соответственно около 3 и 1,5 % сухой биомассы. Содержание ФЛ и НЛ в листьях растений M. lupulina мало отличалось от данных показателей в листьях A. danicus . Однако на долю ФЛ в листьях M. lupulina приходилось почти 45 % ОЛ, тогда как у листьев A. danicus вклад ФЛ в ОЛ был вдвое меньше (табл. 2). Значимых различий в относительном содержании НЛ у листьев исследованных видов бобовых не обнаружено.
Липиды включают разнообразные жирные кислоты (ЖК). Исследования листьев бобовых растений показало наличие в их составе девяти ЖК, причем концентра-
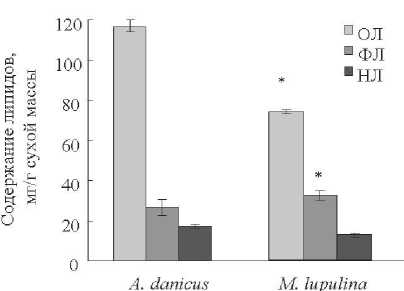
Рисунок 2. Содержание и состав липидов в листьях Astragalus danicus и Medicago lupulina .
Figure 2. Content and composition of lipids in the leaves of Astragalus danicus and Medicago lupulina .
Таблица 2
Относительное содержание нейтральных и фосфолипидов в листьях растений Astragalus danicus и Medicago lupulina (% от суммы общих липидов)
Table 2
Relative content of neutral and phospholipids in the leaves of Astragalus danicus and Medicago lupulina plants (as a percentage of the total lipid content)
Липиды являются основным структурным компонентом мембран, входят в состав запасных веществ. Содержание общих липидов в сухой массе листьев варьирует в зависимости от вида и условий произрастания растений. Общие липиды обычно представляют собой смесь полярных (глико- и фосфолипиды) и нейтральных липидов [4]. Структурной основой всех клеточных мембран являются фосфо- и гликолипиды [9]. Помимо них, в составе мембранных липидов присутствуют также стерины и сфинголипиды. Фосфолипиды являются главными липидами плазмалеммы, митохондрий и других эндомембран. Мем-
Таблица 3
Содержание жирных кислот в листьях Astragalus danicus и Medicago lupulina , % от суммы высших жирных кислот общих липидов
Table 3
Fatty acid content in the leaves of Astragalus danicus and Medicago lupulina , % of the higher fatty acid content of total lipids
Нами получены данные об общем содержании липидов, фосфолипидов и нейтральных липидов в листьях трех видов растений, обитающих в различных условиях на территории заказника «Сойвинский» (Южный Тиман).
Результаты исследования показали, что содержание и соотношение липидов в листьях Aconitum septentrionale зависело от условий местообитания растений. Концентрация всех исследованных групп липидов была выше в листьях растений, произрастающих в ельнике круп-нотравно-злаковом. Листья растений A. septentrionale , обитающего в сероольшаннике, содержали на 36 % меньше ОЛ. В то же время растения достоверно не отличались по содержанию фосфо- и нейтральных липидов, доля которых составляла соответственно 3 и 1 % сухой биомассы листьев. Однако суммарный вклад ФЛ и НЛ в пул ОЛ листьев растений ЦП 2 был в 1,4 раза больше, чем в листьях растений ЦП 1, и составлял 52 %.
Анализ данных, полученных при исследовании липидов Astragalus danicus и Medicago lupulina , выявил различия между этими видами. Растения A. danicus , обитающие в густом травостое, содержали больше липидов, чем растения M. lupulina , произрастающие на открытом месте вдоль дороги. При этом суммарный пул ФЛ и НЛ в листьях люцерны хмелевидной составлял более 60 %, а астрагала датского – 37 % от содержания ОЛ. Изучение жирнокислотного состава выявило тенденцию к снижению соотношения ННЖК и НЖК в листьях M. lupulina , что свидетельствует о более высокой доле НЖК в пуле ЖК листьев у этого вида по сравнению с A. danicus .
В совокупности эти результаты позволяют полагать, что как в случае с двумя ценопопуляциями A. septentrionale, так и в случае с двумя видами бобовых одной из основных причин различия в содержании и качественном составе липидов являются условия местообитания, сглаживающие возможные межвидовые различия. Местообитания отличались по обеспеченности растений важнейшим для фотоавтотрофных организмов ресурсом – фотосинтетической радиацией. Свет является не только драйвером фотосинтеза, но и регулятором многих процессов жизнедеятельности, включая рост и развитие растений. Тот факт, что растения, получающие меньше света, содержали больше липидов, а суммарный вклад ФЛ и НЛ в липидный пул у них был ниже, вероятно, отражает более высокое содержание гликолипидов, связанных с пластидными мембранами. По имеющимся в литературе сведениям, 1 г листьев может содержать до 1 м пластидных мембран [12]. Доля пластидных липидов – глицеролипидов у арабидопсиса – составляла более 50 % всех липидов листа [9]. У исследованных нами растений вклад глицеролипидов в ОЛ листьев может варьи- ровать от 45 до 60 % в зависимости от светового режима местообитания. По-видимому, различия достигаются за счет повышенного содержания гликолипидов в листьях A. septentrionale и A. danicus, произрастающих в условиях более низкого поступления световой радиации. Это согласуется с данными об отличиях световых и теневых растений A. septentrionale по удельной поверхностной плотности листьев и содержанию фотосинтетических пигментов [13].
Растения способны синтезировать более чем 200 различных ЖК. Мы идентифицировали в листьях исследованных видов девять высших ЖК. Модификации пула ЖК отводят важную роль в процессах адаптации растений к условиям произрастания [14]. В литературе имеются данные, свидетельствующие о значении количественных и качественных изменений ЖК для репарации ФС II при фотоингибировании [15]. Изменение состава и содержания ЖК является одним из механизмов поддержания микровязкости плазмалеммы в определенных пределах. Широкий диапазон изменчивости содержания различных ЖК связан как с видовыми особенностями, так и с условиями обитания растений [3, 16, 17]. Нами выявлена тенденция к изменению соотношения ННЖК и НЖК у бобовых видов растений, произрастающих в условиях различного светового режима. Растения на открытых участках, как правило, получают не только больше света, но и тепла. Поэтому снижение относительного содержания ННЖК и повышение НЖК в листьях M. lupulina можно рассматривать как адаптивную реакцию. Насыщенные жирные кислоты уменьшают текучесть мембран и снижают опасность их повреждения в результате перекисного окисления липидов.
Таким образом, полученные нами результаты свидетельствуют о влиянии местообитания на липидный комплекс растений и значимости изучения изменений в содержании и составе липидов для понимания механизмов адаптации растений к условиям окружающей среды.
