Содержание маннита в разных частях таллома Fucus vesiculosus Баренцева моря в различные сезоны года

Автор: Клиндух Мария Петровна
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Гидробиология
Статья в выпуске: 2 т.21, 2018 года.
Бесплатный доступ
Представлены впервые полученные данные по содержанию маннита в разных частях таллома бурой водоросли F. vesiculosus Баренцева моря в различные сезоны года. Водоросли для исследования собирали в губе Белокаменная Кольского залива один раз в сезон в течение года. Определение содержания маннита проводили в разных по физиологической активности и выполняемым функциям участках таллома водоросли (апикальной и средней части таллома, воздушных пузырях и рецептакулах). Установлено существенное влияние климатических особенностей сезона сбора водорослей на содержание маннита в разных частях таллома F. vesiculosus. Определена тенденция изменения содержания маннита во всех исследуемых частях таллома: наименьшее содержание маннита выявлено в зимний период (90,9-99,2 мг/г сухой массы), в летне-осенний период его содержание повышалось до наибольшего уровня (111,4-150,4 мг/г сухой массы). Полученные данные по изменению содержания маннита в разных частях таллома бурой водоросли F. vesiculosus в различные сезоны года соответствовали ранее полученным данным по изменению содержания маннита в средней пробе. В отличие от Saccharina latissima ( Laminaria saccharina ), произрастающей в бухте Оскара Баренцева моря, высокой гетерогенности в распределении маннита по таллому бурой водоросли F. vesiculosus не наблюдалось. В январе, апреле и октябре значительных различий в содержании маннита в разных частях таллома бурой водоросли F. vesiculosus выявлено не было. В августе установлена наибольшая гетерогенность в содержании маннита в разных частях таллома F. vesiculosus. Содержание маннита в рецептакулах F. vesiculosus в апреле и августе оказалось практически одинаковым, несмотря на разную степень зрелости репродуктивных клеток.
Маннит, разные части таллома, сезонные изменения, бурые водоросли
Короткий адрес: https://sciup.org/142215135
IDR: 142215135 | УДК: 581.19 | DOI: 10.21443/1560-9278-2018-21-2-237-244
Mannitol content in different parts of Fucus vesiculosus thallus from the Barents Sea in different seasons
The data on the mannitol content in different parts of the brown algae F. vesiculosus thallus from the Barents Sea have been obtained for the first time in different seasons. Seaweed for research was collected in the Belokamennaya Bay of the Kola Bay once a season during the year. Determination of mannitol has been carried out in different parts of the thallus of algae in the physiological activity and functions performed: the apical part, the middle part of the thallus, the vesicles and the receptacles. It has been determined that the season of algae harvest significantly affects the mannitol content in different parts of the F. vesiculosus thallus. The trend of changes in the mannitol content in all researched parts of the thallus has been found: the lowest mannitol content is detected in winter (90.9-99.2 mg/g dry weight), in the summer - autumn period its content has increased to the highest within a year (111.4-150.4 mg/g dry weight). The obtained data on the change in the mannitol content in different parts of the brown alga F. vesiculosus thallus in different seasons corresponded to the previously obtained data on the change in the mannitol content in the mean sample. Unlike Saccharina latissima ( Laminaria saccharina ) growing in the Oscar Cove of the Barents Sea, high heterogeneity in the mannitol distribution over the brown alga F. vesiculosus thallus has not been observed. In January, April and October, there was no significant difference in the mannitol content in different parts of the brown alga F. vesiculosus thallus. In August, the greatest heterogeneity in the mannitol content in different parts of the F. vesiculosus thallus was determined. The mannitol content in F. vesiculosus receptacles in April and August was almost identical despite the varying maturity of the reproductive cells.
Текст научной статьи Содержание маннита в разных частях таллома Fucus vesiculosus Баренцева моря в различные сезоны года
Маннит (шестиатомный алифатический спирт) является одним из наиболее часто встречающихся многоатомных спиртов в природе. Маннит был обнаружен в более чем 100 видах высших растений [1]; также он синтезируется многими водорослями разных отделов [2–6].
В растительном организме маннит выполняет различные функции: накапливается в качестве запасного питательного вещества, используется при синтезе других соединений, выполняет осморегуляторную и антиоксидантную функции [7; 8]. В бурых водорослях маннит играет важную роль в энергетическом обмене, являясь первичным стабильным продуктом фотосинтеза и субстратом для дыхания [9; 10].
В зависимости от вида водоросли, стадии развития растения, сезона сбора, внешних факторов среды произрастания содержание маннита может варьировать от совсем незначительного количества (менее 0,5 %) до 33 % сухой массы водоросли [5; 11–14]. Сезонные изменения уровня содержания маннита у некоторых видов семейства Sargassaceae связывают со стадиями развития водоросли. Так, наибольшая концентрация маннита в бурых водорослях наблюдается на стадии роста. После закладки репродуктивных структур его содержание снижается до минимума [5; 11; 15]. Но подобные изменения характерны не для всех водорослей. Для водорослей Sargassum myriocystum, S. ilicifolium и Turbinaria decurrens не было выявлено четких изменений в содержании маннита в зависимости от сезона или стадии развития растений. Причиной подобных нерегулярных изменений могут быть медленно протекающие ростовые процессы или непостоянство сроков размножения у одного и того же вида [16–18]. Сезонные изменения содержания маннита также были изучены и у бурых водорослей, произрастающих на побережье Баренцева моря. При этом наиболее высокое содержание маннита наблюдалось в летне-осенний период, а зимой его содержание снижалось до минимальных значений [19; 20].
Также известно, что маннит перераспределяется внутри таллома бурых водорослей в зависимости от энергетических потребностей и физиологических функций различных участков слоевища [21–24]. Были исследованы изменения содержания маннита в разных частях таллома некоторых видов водорослей. В зависимости от стадии роста таллома или степени зрелости репродуктивных структур различий в содержании маннита либо вообще не наблюдалось, либо его концентрация различалась более чем в пять раз в разных участках таллома [19; 25; 26].
Для широко распространенной по побережью Баренцева моря бурой водоросли Fucus vesiculosus Linnaeus (1753) подобных работ не проводилось. Цель данной работы заключалась в исследовании концентрации маннита в разных по физиологической активности и выполняемым функциям частях таллома F. vesiculosus в разные периоды года; полученные сведения позволят оценить гетерогенность его содержания в талломе в зависимости от сезона года.
Материалы и методы
В качестве объекта исследования был выбран широко распространенный на побережье Баренцева моря вид бурых водорослей Fucus vesiculosus Linnaeus (1753), произрастающий в губе Белокаменная Кольского залива. Материал для исследования собирали один раз в сезон в течение года. Возраст собранных растений составлял 5–7+ лет.
Для исследования брали отдельные участки таллома водоросли: апикальную и среднюю части, воздушные пузыри и рецептакулы; схема исследуемых участков таллома представлена на рис. 1.
Определение содержания маннита пробы (в мг/г сухой массы) проводили согласно методу Кэмерона [27]. Все полученные данные были представлены в виде арифметического среднего трех параллельных измерений с доверительным интервалом.
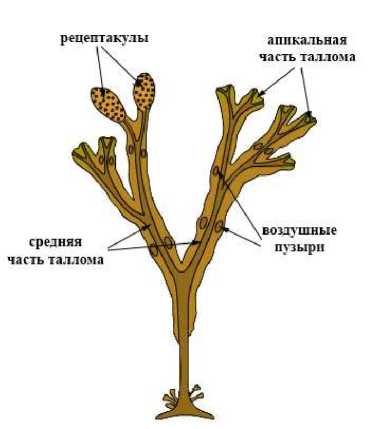
Рис. 1. Схематическое расположение исследуемых участков таллома F. vesiculosus
Fig. 1. Schematic location of researched parts of thallus F. vesiculosus
Результаты и обсуждение
Данные по содержанию маннита в разных частях таллома F. vesiculosus в различные сезоны года представлены на рис. 2.
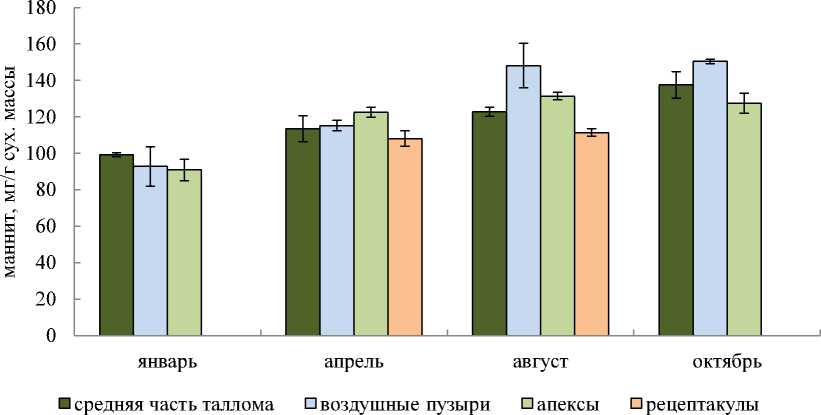
Рис. 2. Содержание маннита в разных частях таллома F. vesiculosus в различные сезоны года Fig. 2. The mannitol content in different parts of thallus F. vesiculosus at different seasons
В январе для исследуемой водоросли F. vesiculosus характерно минимальное содержание маннита (90,9–99,2 мг/г сухой массы) во всех участках таллома. Между разными частями таллома существенной разницы в содержании маннита с учетом доверительного интервала не наблюдалось.
В апреле содержание маннита оказалось более высоким, чем в январе, во всех исследуемых частях таллома. В разных участках увеличение содержания маннита оказалось различным: в средней части таллома количество маннита увеличилось на 14,5 % по сравнению с содержанием в январе, в воздушных пузырях – на 24,1 %, а в апексах – на 34,8 %. При этом между частями таллома различий в содержании маннита (так же, как и в январе) не наблюдалось. А вот концентрация маннита в рецептакулах с учетом доверительного интервала оказалась ниже, чем в апикальных частях таллома.
В августовских пробах водорослей содержание маннита увеличилось в воздушных пузырях и апексах по сравнению с весенними пробами на 28,6 и 7,3 % соответственно. В средней части таллома и рецептакулах изменений в содержании маннита не произошло. По количеству маннита в августе части таллома отличались между собой: больше всего его содержалось в воздушных пузырях (148,1 мг/г сухой массы), поменьше – в апексах (131,4 мг/г сухой массы), еще меньше – в средней части таллома (122,8 мг/г сухой массы). В рецептакулах концентрация маннита оказалась самой низкой в августе по сравнению с другими рассматриваемыми частями таллома и составила 111,4 мг/г сухой массы. Различия в содержании маннита между участками таллома F. vesiculosus в августе составили 24,8–32,9 % от содержания в пробах.
В октябрьских пробах водоросли концентрация маннита увеличилась только в средней части таллома на 12,1 % по сравнению с летним содержанием. В остальных участках таллома с учетом доверительного интервала изменений в содержании маннита по сравнению с предыдущим периодом года не произошло. Между частями таллома по количеству маннита различия выявлены только между воздушными пузырями и остальными исследуемыми участками таллома. В воздушных пузырях содержалось на 9,3 и 18 % больше маннита, чем в средней части таллома и апексах соответственно.
Для определения значимости влияния сезона сбора на содержание маннита в разных частях таллома бурой водоросли F. vesiculosus был проведен однофакторный дисперсионный анализ. Определено, что содержание маннита в разных участках таллома F. vesiculosus подвергается сильным изменениям в течение года: в зимний период содержание маннита наименьшее, к лету – осени прослеживается увеличение содержания маннита (табл., рис. 2). В рецептакулах содержание маннита определялось лишь в апреле и августе и практически не изменилось за этот промежуток времени (рис. 2).
В исследованиях, проведенных на других видах бурых водорослей, указывается на связь изменения в содержании маннита со стадиями развития водорослей и развитием репродуктивных органов. Во время интенсивной стадии роста наблюдается наибольшее количество маннита, а снижение его концентрации связано с развитием репродуктивных органов [5; 11]. Некоторые исследователи считают, что динамика в содержании маннита свидетельствует об изменениях фотосинтетической деятельности водорослей, поскольку он является основным продуктом фотосинтеза у бурых водорослей [25].
Полученные данные по изменению содержания маннита в разных частях таллома бурой водоросли F. vesiculosus в различные сезоны года совпадают с ранее полученными данными по содержанию в средней пробе: увеличение содержания маннита в летне-осенний период и уменьшение его содержания зимой как для средней пробы, так и для отдельных частей таллома [20].
Таблица. Изменения содержания маннита в разных участках таллома F. vesiculosus и результаты однофакторного дисперсионного анализа по динамике содержания маннита в течение года в разных участках таллома
Table. Changes in mannitol content in different parts of thallus F. vesiculosus and the results of ANOVA in the dynamics of mannitol content throughout the year in different parts of the thallus
|
Участок таллома F.vesiculosus |
Содержание маннита, мг/г сухой массы (мин. – макс.) |
Результаты дисперсионного анализа |
|
|
F* |
p |
||
|
Апикальная часть |
90,9–131,4 |
326,85 |
–0,0001 |
|
Средняя часть |
99,2–137,6 |
172,98 |
–0,0001 |
|
Воздушные пузыри |
92,8–150,4 |
207,53 |
–0,0001 |
Примечание. *F – значение критерия Фишера.
В полярную ночь (зимой) у водорослей-макрофитов Мурмана дыхание преобладает над фотосинтетическими процессами. Только с середины февраля фотосинтез начинает преобладать над дыханием. Вместе с тем многие многолетние водоросли способны начинать рост в полярную ночь в районе исследования. Рост водорослей в это время может идти только за счет запасных веществ. Период активного роста длится с марта по август и делится на две фазы: 1) интенсивное формирование фотосинтетической поверхности (март – июнь), когда интенсивность роста у большинства видов достигает своих максимальных значений; 2) накопление массы и завершение вегетативного развития (июль – август), когда интенсивность роста снижается, но значения скорости роста длины и массы растений могут быть еще высокими [28]. Формирование рецептакул у F. vesiculosus и F. distichus начинается осенью, а зрелые рецептакулы встречаются в конце весны – начале лета. Выход репродуктивных клеток и развитие проростков наблюдается летом [29]. В летне-осенний период происходит накопление запасных веществ в макрофитах.
В зимний период из-за низкого уровня освещенности и начала ростовых процессов наблюдается минимальное содержание маннита во всех частях таллома F. vesiculosus.
Увеличение содержания маннита весной по сравнению с зимним периодом в разных частях таллома водоросли было наибольшим, что, скорее всего, связано с возрастанием фотосинтетической активности. В этот период маннит активно образуется в процессе фотосинтеза, но он также и расходуется в процессе роста и созревания органов размножения.
В летний период происходит снижение скорости роста таллома и завершается процесс созревания репродуктивных клеток. Существенное увеличение маннита происходит только в воздушных пузырях и небольшое – в апексах. Вероятно, расход маннита в растении на синтез других веществ (запасных, структурных) возрастает по сравнению с его образованием в процессе фотосинтеза. Наибольшая гетерогенность по содержанию маннита в разных частях таллома была определена в августе, что, возможно, связано с его неравномерным использованием в каждом отдельном участке таллома: в рецептакулах завершается процесс созревания гамет, в талломе происходит синтез запасных полисахаридов и продолжается отток маннита в апикальную часть, где он расходуется на рост таллома и синтез полисахаридов.
В октябре наблюдалось небольшое увеличение содержания маннита в средней части таллома по сравнению с августом, тогда как в остальных частях таллома содержание не поменялось. Причиной этому, возможно, является снижение уровня освещенности и, как следствие, снижение фотосинтетической активности и синтеза маннита в водоросли.
Изменений содержания маннита в рецептакулах в процессе созревания репродуктивных клеток выявлено не было.
Для Saccharina latissima (Laminaria saccharina), собранной в бухте Оскара Баренцева моря, показана высокая гетерогенность в распределении маннита в пределах одной пластины. Наибольшее содержание маннита обнаружено у основания пластины в зоне интеркалярного роста, по мере приближения к верхнему дистальному концу оно снижается [19]. В данном исследовании значительных различий в содержании маннита в апикальной и средней частях таллома не наблюдалось. В январе, апреле и октябре содержание маннита в этих частях было практически одинаковым, а в августе разница в содержании составила всего 8,6 мг/г сухой массы. Возможно, это связанно с тем, что интенсивность фотосинтетических процессов и скорость расхода маннита (в том числе его отток из средней части таллома в апикальную) в данных участках таллома у F. vesiculosus протекают в соотношении, обеспечивающем практически одинаковое содержание маннита в этих участках таллома.
Заключение
В результате проведенных исследований получены данные, расширяющие имеющиеся сведения по содержанию маннита в разных участках таллома бурой водоросли F. vesiculosus Баренцева моря в различные сезоны года.
Тенденция изменения содержания маннита в течение года во всех исследуемых частях таллома была одинакова: наименьшее содержание маннита в водоросли наблюдалось зимой (90,9–99,2 мг/г сухой массы), в летне-осенний период его содержание повышалось до наибольшего значения (111,4–150,4 мг/г сухой массы).
Наибольшие различия в содержании маннита в исследованных частях таллома F. vesiculosus были выявлены в августе. Распределение маннита по таллому F. vesiculosus в остальные месяцы с учетом доверительного интервала было равномерным.
Работа выполнена в рамках государственного задания ММБИ КНЦ РАН на 2018 г. по теме № 0228-2018-0005 "Научные основы инновационных технологий БАВ водорослей Баренцева моря".
Список литературы Содержание маннита в разных частях таллома Fucus vesiculosus Баренцева моря в различные сезоны года
- Stoop J. M. H., Williamson J. D., Pharr D. M. Mannitol metabolism in plants: a method for coping with stress//Trends in Plant Science. 1996. V. 1, N 5. P. 139-144. DOI: https://doi.org/10.1016/S1360-1385(96)80048-3.
- Mehta B. R., Parekh R. G. Mannitol content in brown algae of the coast of Saurashtra//Botanica Marina. 1978. V. 21, Iss. 4. P. 251-252. DOI: https://doi.org/10.1515/botm.1978.21.4.251.
- Репина О. И., Муравьева Е. А., Подкорытова А. В. Динамика химического состава промысловых бурых водорослей Белого моря//Труды ВНИРО. 2004. Т. 143. С. 93-99.
- Аминина Н. М., Вишневская Т. И., Гурулева О. Н., Ковековдова Л. Т. Состав и возможности использования бурых водорослей дальневосточных морей//Вестник Дальневосточного отделения РАН. 2007. № 6. С. 123-130.
- Zubia M., Payri C., Deslandes E. Alginate, mannitol, phenolic compounds and biological activities of two range-extending brown algae, Sargassum mangarevense and Turbinaria ornate (Phaeophyta: Fucales), from Tahiti (French Polynesia)//Journal of Applied Phycology. 2008. V. 20, Iss. 6. P. 1033-1043. DOI: https://doi.org/10.1007/s10811-007-9303-3.
- Кайшева Н. Ш., Архипова М. Н., Кайшев А. Ш. Морфолого-анатомические и фитохимические характеристики некоторых водорослей//Фармация и фармакология. 2014. № 3. С. 28-42. DOI: https://doi.org/10.19163/2307-9266-2014-2-3(4)-28-42.
- Iwamoto K., Shiraiwa Y. Salt-regulated mannitol metabolism in algae//Marine Biotechnology. 2005. V. 7, Iss. 5. P. 407-415. DOI: https://doi.org/10.1007/s10126-005-0029-4.
- Gylle A. M., Nygård C. A., Ekelund N. G. A. Desiccation and salinity effects on marine and brackish Fucus vesiculosus L. (Phaeophyceae)//Phycologia. 2009. V. 48, N. 3. P. 156-164 DOI: https://doi.org/10.2216/08-45.1
- Bidwell R. G. S. Photosynthesis and metabolism of marine algae: II. A survey of rates and products of photosynthesis in C14O2//Canadian Journal of Botany. 1958. V. 36, N 3. P. 337-349. 10.1139/b58-029 DOI: https://doi.org/
- Yamagughi T., Ikawa T., Nisizawa K. Pathway of mannitol formation during photosynthesis in brown algae//Plant & Cell Physiology. 1969. V. 10, Iss. 2. P. 425-440. oxfordjournals.pcp.a074421 DOI: https://doi.org/10.1093/
- Umamaheswara Rao M., Kalimuthu S. Changes in mannitol and alginic acid contents of Turbinaria ornata (Turner) J. Agardh in relation to growth and fruiting//Botanica Marina. 1972. V. 15, Iss. 1. P. 57-59. DOI: https://doi.org/10.1515/botm.1972.15.1.57.
- Joshi G.V., Gowda C.A. Seasonal variations in chemical composition of Sargassum ilicifolium Grun. and sea water//Indian Journal of Marine Sciences. 1975. V. 4. P. 165-168. URL: http://nopr.niscair.res.in/handle/123456789/39690.
- Gomez I. M. Life strategy and ecophysiology of Antarctic macroalgae = Lebensstrategie und Ökophysiologie mariner Braunalgen der Arktis//Berichte zur Polarforschung; Reports on Polar Research. Bremerhaven, 1997. V. 238.
- Облучинская Е. Д. Сравнительное исследование бурых водорослей Баренцева моря//Прикладная биохимия и микробиология. 2008. Т. 44, № 3. С. 337-342.
- Umamaheswara Rao M. Seasonal variations in growth, alginic acid and mannitol contents of Sargassum wightii and Turbinaria conoides from the gulf of Manna, India//Proceedings of International Seaweed Symposium. 1969. V. 6. P. 579-584.
- Kaliaperumal N., Kalimuthu S. Changes in growth, alginic acid and mannitol contents of Turbinaria decurrens Bory//Botanica Marina. 1976. V. 19, Iss. 3. P. 157-159. DOI: https://doi.org/10.1515/botm.1976.19.3.157.
- Kalimuthu S. Variations in growth and mannitol and alginic acid contents of Sargassum myriocystum J. Agardh//Indian Journal of Fisheries. 1980. V. 27, Iss. 1-2. P. 265-266.
- Chennubhotla V. S. K., Kaliaperumal N., Kalimuthu S., Selvaraj M., Ramalingam J. R. Seasonal changes in growth and alginic acid and mannitol contents in Sargassum ilicifolium (Turner) J. Agardh and S. myriocystum J. Agardh//Indian Journal of Marine Sciences. 1982. V. 11. P. 195-196. URL: http://nopr.niscair.res.in/handle/123456789/38956.
- Промысловые и перспективные для использования водоросли и беспозвоночные Баренцева и Белого морей/отв. ред. Г. Г. Матишов. Апатиты: КНЦ РАН, 1998. 628 с.
- Клиндух М. П., Облучинская Е. Д., Матишов Г. Г. Сезонные изменения содержания маннита и пролина в бурой водоросли Fucus vesiculosus L. Мурманского побережья Баренцева моря//Доклады академии наук. 2011. Т. 441, № 1. С. 133-136.
- Nicholson N. L., Briggs W. R. Translocation of photosynthate in the brown alga Nereocystis//American Journal of Botany. 1972. V. 59, N 1. P. 97-106. DOI: https://doi.org/10.1002/j.1537-2197.1972.tb10067.x.
- Schmitz K., Luning K., Willenbrink J. CO2-fixierung und stofftransport in benthischen marinen algen. II. zum ferntransport 14C-markierter assimilate bei laminaria hyperborea und laminaria saccharina = CO2-fixation and translocation in benthic marine algae. II. On translocation of 14C-labelled assimilates in Laminaria hyperborea and L. saccharina//Zeitschrift für Pflanzenphysiologie. 1972. Bd. 67. S. 418-429. DOI: https://doi.org/10.1016/S0044-328X(72)80042-4.
- Schmitz K., Srivastava L. M. On the fine structure of sieve tubes and the physiology of assimilate transport in Alaria marginata//Canadian Journal of Botany. 1975. Vol. 53, N 9. P. 861-876 DOI: https://doi.org/10.1139/b75-104
- Schmitz K., Srivastava L. M. Long distance transport in Macrocystis integrifolia. I. Translocation of 14C-labelled assimilates//Plant Physiology. 1979. V. 63, Iss. 6. P. 995-1022 DOI: https://doi.org/10.1104/pp.63.6.995
- Thomas P. C., Subbaramaiah K. Seasonal variations in growth, reproduction, alginic acid, mannitol, iodine and ash contents of brown alga Sargassum wightii//Indian Journal of Marine Sciences. 1991. V. 20. P. 169-175. URL: http://nopr.niscair.res.in/handle/123456789/38278.
- Конева А. А., Аминина Н. М. Химический состав различных частей слоевища Alaria marginata P. ET R. (Laminariales, Phaeophyta)//Известия ТИНРО. 2013. Т. 175. С. 333-338.
- Cameron M. C., Ross A. G., Percival E. G. V. Methods for the routine estimation of mannitol, alginic acid, and combined fucose in seaweeds//Journal of the Society of Chemical Industry. 1948. V. 67. P. 161-164 DOI: https://doi.org/10.1002/jctb.5000670410
- Шошина Е. В. Динамика ростовых и репродуктивных процессов у водорослей Баренцева моря: дис.. д-ра биол. наук: 03.00.05. Мурманск, 2001. 266 с.
- Кузнецов Л. Л., Шошина Е. В. Фитоценозы Баренцева моря: Физиологические и структурные характеристики/отв. ред. Г. Г. Матишов, А. Д. Чинарина. Апатиты: КНЦ РАН, 2003. 308 с.
