Содержание пигментов в растениях Lepidium sativum в условиях хлоридно-натриевого засоления и ощелачивания

Автор: Еремченко О.З., Кусакина М.Г., Лузина Е.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Экология
Статья в выпуске: 1, 2014 года.
Бесплатный доступ
Представлены результаты исследований влияния засоления и ощелачивания корневого субстрата на ростовые процессы и пигментный аппарат растений кресс-салата (Lepidium sativum L., Brassicaceae). Показано, что повышение содержания фотосинтетических пигментов в листьях способствует адаптации растений к действию неблагоприятных факторов среды. Установлены корреляции между высотой, массой растений и содержанием в них хлорофиллов и каротиноидов.
Кросс-адаптация, засоление, ощелачивание, хлорофиллы, антиок-сидантная защита, каротиноиды
Короткий адрес: https://sciup.org/147204663
IDR: 147204663 | УДК: 581.1:632.1
The content of pigments in Lepidium sativumin the conditions of chloride and sodium salinization and the alkalization
Results of researches of influence of salinization and alkalization of a root substratum on thepigmental device of plants of Lepidium sativum (Brassicaceae) are presented. It is shown, that increase of the contents and a ratio of photosynthetic pigments in leaves promote adaptation of plants to action of adverse factors of the environment. Correlations between height, mass of plants and the content in them of a chlorophyll and carotinoids are established.
Текст научной статьи Содержание пигментов в растениях Lepidium sativum в условиях хлоридно-натриевого засоления и ощелачивания
Введение фотоокислительного стресса и защищают клетки
Неблагоприятные условия среды, такие как низкие температуры, засуха, засоление и др., оказывают существенное влияние на фотосинтетический аппарат растений. Фотосинтез и дыхание являются процессами, которые обеспечивают материально-энергетическую основу жизнедеятельности растений. Они поставляют необходимые для роста и поддержания растительных клеток АТФ, НАД(Ф)Н и метаболиты.
Высокий уровень засоления отрицательно действует на структурно-функциональное состояние хлоропластов, процесс циклического фосфорилирования, на реакционные центры фотосистемы [Структура …, 1970; Белецкий, Шевякова, Карнаухова, 1990].
Стрессовые факторы, в том числе засоление, существенно повышают вероятность развития фотоо-кислительных повреждений. В клетках растений образование активных форм кислорода наиболее интенсивно идет в хлоропластах. В составе липидов мембран хлоропластов содержится большое количество ненасыщенных жирных кислот, значительная часть супероксидных анионов возникает при функционировании электрон-транспортной цепи фотосинтеза, образование кислородных радикалов происходит при фотоокислении воды [Перекисное ..., 1997].
В тушении избыточной энергии триплетных состояний хлорофилла и синглетного кислорода важная роль принадлежит каротиноидам. Являясь антиоксидантами, каротиноиды уменьшают последствия растений от повреждений [Чиркова, 2002; Половни-кова, Воскресенская, 2008; Bolwell, 1999; Mittler, 2002; Foyer, Shigeoka, 2011].
Повышение устойчивости растений к засолению и другим неблагоприятным факторам среды связано со значительными перестройками физиолого-биохимических процессов и с изменением экспрессии довольно большого количества генов [Сигнальная роль ..., 2012].
В настоящее время обсуждается возможность существования комплексной устойчивости, которая обусловлена реакцией различных метаболических систем. В ответ на действие определенного стрессора устойчивость клеток повышается не только к нему, но и к другим повреждающим факторам. Литературные данные свидетельствуют о функционировании одних и тех же систем устойчивости к двум или нескольким факторам различной природы [Hale, 1969; Общие системы ..., 1990; Стрессорный ответ ..., 1994; Маевская, Николаева, 2013]. Эти механизмы направлены на экономию энергетических и структурных ресурсов растительных организмов в стрессовых условиях.
Участие пигментного аппарата в устойчивости и адаптации растений к комбинированному действию засоления и ощелачивания является недостаточно исследованным. В связи с этим целью нашей работы было изучение раздельного и комбинированного действия засоления и ощелачивания на ростовые
процессы и содержание фотосинтетических пигментов в растениях кресс-салата.
Объект и методы исследования
В качестве тест-культуры были взяты растения кресс-салата ( Lepidium sativum L., Brassicaceae ), достигшие состояния технической спелости. Поставлены модельные опыты, субстратом являлся вермикулит. Для ощелачивания корневой среды (рН = 8, 9, 10) использовали раствор гидроксида натрия, для засоления – 0.5%-ный раствор хлорида натрия. Кресс-салат выращивался в сосудах на среде Кнопа, pH раствора контролировали при помощи pH-метра. Исследовали раздельное и комбинированное действие засоления и ощелачивания. В качестве контроля служили растения, выращенные на среде Кнопа с рН = 7.
Показатели роста и сырой биомассы определяли путем измерения надземной части 25 растений и их взвешивания на аналитических весах.
Количество пигментов определяли спектрофотометрическим методом при длинах волн 665, 649 и 440 нм. Для расчета концентрации хлорофиллов а , б и каротиноидов использовали формулы Винтер-манса де Мотса [Гавриленко, Жигалова, 2003]. Повторность опытов трехкратная. Полученные данные статистически обработаны с помощью компьютерной программы Stadia. Показатель достоверности различий между опытными вариантами и контрольным был установлен с использованием однофакторного дисперсионного анализа. Определяли корреляционные зависимости между содержанием пигментов, высотой и массой растений.
Результаты и их обсуждение
Рост как интегральный процесс является одним из важных показателей адаптации растений к условиям обитания. Он является сложной функцией многих физиолого-биохимических процессов.
Изучение раздельного влияния изучаемых факторов показало, что в вариантах с рН корнеобитаемой среды, равной 8 и 9, масса растений и их вы- сота не уступали контролю или даже чуть превы-
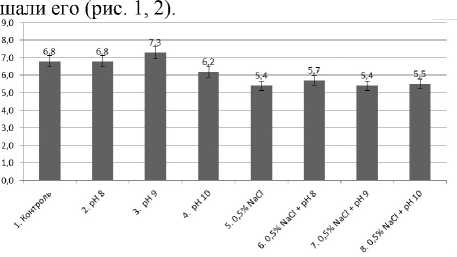
Рис. 1 . Влияние ощелачивания и засоления на высоту надземной части кресс-салата, см
Вероятно, это связано с тем, что корневая система растений способна смещать рН окружающей их питательной среды и создавать в зоне, непосредственно прилегающей к корням, благоприятные условия рН. Таким образом, находясь в зависимости от среды, растения способны в известной степени сами воздействовать на величину рН, как бы приспосабливая среду к своим потребностям. Регулирование рН достигается путем различного поглощения растением из питательной среды анионов и катионов [Якушкина, Бахтенко, 2005].
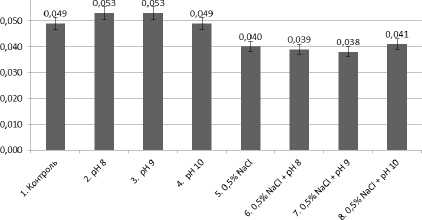
Рис. 2. Влияние ощелачивания и засоления на массу растений, г
Проведенный однофакторный дисперсионный анализ свидетельствует о том, что действие засоления (0.5% NaCl) достоверно ингибировало рост и массу кресс-салата по отношению к контролю. Отдельное действие высокого уровня ощелачивания (рН = 10) повлияло на высоту растений.
В природных условиях растения кресс-салата произрастают в условиях засушливых и полузасу-шливых районов, где им приходится переживать периоды дефицита воды. У кресс-салата отсутствуют эволюционно сложившиеся механизмы соле-устойчивости. Адаптация таких растений может идти за счет включения эффективных механизмов засухоустойчивости, к которым относится способность к осморегуляции, стабильность водного и осмотического потенциалов содержимого клеток растений [Hare, Gress, Van Stader, 1998; Patakas, Noitsakis, 2000].
Ингибирующего влияния ощелачивания на фоне засоления корневой среды не установлено, даже при рН = 10, вероятно, при комбинировании факторов имеет место кросс-адаптация – повышение устойчивости за счет неспецифических внутриклеточных процессов.
Как видно из данных табл. 1, наблюдается сильная прямая корреляция (r = 0.94) между высотой растений и их массой. Неблагоприятное действие засоления на рост и массу растений связано как со снижением доступности воды, так и с токсическим действием ионов натрия и хлора на клетки растений [Захарин, Паничкин, 2009]. Под влиянием засоляющих ионов происходит торможение синтеза белков и нуклеиновых кислот, что влечет за собой снижение ростовых процессов и сокращение темпов нарастания биомассы. Торможение роста ли- стьев является защитной реакцией на дефицит воды за счет снижения транспирации [Lu, воды, так как уменьшение площади листовой по- Neumann, 1998; Иванов, 2010].
верхности позволяет растениям сократить потери
Таблица 1
Коэффициенты корреляции между изучаемыми показателями
|
Показатели |
Высота, см |
Масса, г |
Хлорофилл а , мг/г |
Хлорофилл в , мг/г |
Хлорофиллы а + в , мг/г |
Каротиноиды, мг/г |
|
Высота, см |
1.00 |
0.94 |
–0.81 |
–0.69* |
–0.81 |
–0.65* |
|
Масса, г |
0.94 |
1.00 |
–0.71 |
–0.74 |
–0.77 |
–0.48* |
|
Хлорофилл а , мг/г |
–0.81 |
–0.71 |
1.00 |
0.69* |
0.95 |
0.91 |
|
Хлорофилл в , мг/г |
–0.69* |
–0.74 |
0.69* |
1.00 |
0.86 |
0.37* |
|
Хлорофиллы а + в , мг/г |
–0.81 |
–0.77 |
0.95 |
0.86 |
1.00 |
0.74 |
|
Каротиноиды, мг/г |
–0.65* |
–0.48* |
0.91 |
0.37* |
0.74 |
1.00 |
Примечание. Данные статистически достоверны при 0.95%-ном уровне значимости, за исключением помеченных *.
При стрессовых воздействиях, таких как засуха, засоление, действие низких температур и др., большое значение принадлежит гормональной системе регуляции. Снижение уровня гормонов-сти-муляторов, таких как индолилуксусная кислота (ИУК), гиббереллины и накопление ингибиторов роста – абсцизовой кислоты (АБК) и этилена, которые называют стрессовыми гормонами, имеет адаптивное значение, поскольку приводит к снижению интенсивности обмена веществ, торможению процессов клеточного деления и растяжения [Чиркова, 2002]. Абсцизовая кислота ингибирует деятельность Н+-АТФаз, что ведет к снижению рН цитоплазмы и усилению гидролитических процессов.
В условиях солевого стресса поддержание ионного и водного гомеостаза клеток осуществляется с помощью протонных насосов плазма-леммы и тонопласта. В результате их работы на мембранах плазмалеммы и тонопласта генерируется электрохимический потенциал протонов ∆µН+, от которого зависит не только вторично-активный, но и пассивный транспорт ионов, в том числе Na+и Cl- [Палладина, 1991].
ИУК способна усиливать функционирование протонного насоса плазмалеммы путем изменения содержания в мембранах ферментативных белков, а также их молекулярной активности. Высказывается мнение, что ИУК, также как и АБК, играет в растениях роль «стрессового сигнала» при засолении [Сигнальная роль ..., 2012].
Исследование состояния пигментного аппарата имеет важное значение для понимания механизмов адаптации растительных организмов к действию стрессовых факторов. Хлорофиллы а и б являются основными фотосинтетически деятельными компонентами листьев растений. Содержание и соотношение разных форм пигментов в хлоропластах может служить одним из показателей их фотохимической активности. Поддержание структуры и функции мембран хлоропластов в стабильном состоянии при действии неблагоприятных факторов среды обеспечивает более скоординированную работу компонентов электрон-транспортной цепи [Влияние гена ..., 2011].
Литературные данные о влиянии засоления на уровень хлорофиллов в листьях растений противоречивы. Одни авторы [Структура ..., 1970; Га-рифзянов, Горелова, 2006] приводят данные о снижении количества этих пигментов при засолении, другие [Шахов, 1956; Удовенко, Семухина, Петроченко, 1971; Кусакина, Еремченко, Четина, 2013] отмечают их повышение или отсутствие существенных изменений.
Результаты определения содержания зеленых пигментов свидетельствуют о том, что в вариантах с увеличением степени ощелачивания корневой среды количество хлорофиллов а и б было достоверно выше по сравнению с контрольным вариантом (табл. 2).
Таблица 2
Влияние ощелачивания и засоления на содержание пигментов в растениях кресс-салата, мг/г сырой массы
|
Вариант |
Хлорофилл а |
Хлорофилл в |
Хлорофиллы а + в |
Каротиноиды |
|
Контроль |
2.44±0.15 |
0.64±0.03 |
3.08±0.17 |
0.88±0.05 |
|
pH 8 |
2.78±0.06 |
0.78±0.04 |
3.55±0.11 |
1.02±0.02 |
|
pH 9 |
2.89±0.10 |
0.80±0.05 |
3.69±0.15 |
1.05±0.04 |
|
pH 10 |
3.08±0.22 |
0.90±0.05 |
3.98±0.27 |
1.12±0.09 |
|
0.5 % NaCl |
3.24±0.11 |
0.83±0.03 |
4.07±0.13 |
1.17±0.06 |
|
0.5 % NaCl + pH 8 |
3.08±0.17 |
1.42±0.11 |
4.49±0.28 |
1.02±0.05 |
|
0.5 % NaCl + pH 9 |
3.43±0.13 |
1.44±0.05 |
4.87±0.18 |
1.12±0.05 |
|
0.5 % NaCl + pH 10 |
3.52±0.12 |
1.13± 0.28 |
4.98±0.19 |
1.18±0.04 |
В большей степени это проявилось в отношении такого фотосинтетического пигмента, как хлорофилл а . Увеличение биосинтеза хлорофилла а , вероятно, является одной из приспособительных реакций кресс-салата к действию засоления и ощелачивания корневой среды.
Комбинированное действие ощелачивания и засоления способствовало более значительному повышению содержания хлорофиллов а и б , а также их суммы в сравнении с контролем и раздельным влиянием данных изучаемых факторов. Это явление можно рассматривать как кросс-адаптацию, когда растения, адаптируясь к действию одного фактора, приобретают устойчивость к другому. Сопряженное повышение устойчивости к обезвоживанию и перегреву, тепловому стрессу и засолению [Общие системы ..., 1990].
Сопоставляя полученные данные с ростовыми параметрами, можно отметить, что повышение содержания пигментов коррелировало со снижением роста культуры. Из табл. 1 видно, что между высотой, массой и содержанием хлорофиллов а установлена сильная обратная корреляция (r = –0.81 и r = –0.71 соответственно). Накопление пигментов может быть следствием торможения ростовых процессов под влиянием токсического действия солей при продолжающемся биосинтезе пигментов [Удовенко, 1977]. Повышение количества этих соединений является активной реакцией растений в противовес неблагоприятному действию засоляющих ионов.
Представляет интерес отношение содержания хлорофиллов а и б . Хлорофилл а является компонентом как коровых комплексов, так и периферической антенны фотосистемы I (ФС I) и фотосистемы II (ФС II), тогда как хлорофилл б является компонентом периферической антенны обеих фотосистем. Он входит преимущественно в белковый светособирающий комплекс ФСII, так называемый ССК II [Бухов, Бондарь, Дроздова, 1998; Vermaas, 1993]. Поэтому изменение отношения хлорофиллов а / в может рассматриваться как нарушение в стехиометрии между комплексами реакционных центров фотосистем и ССК II [Бухов, Бондарь, Дроздова, 1998; Дымова, Головко, 2007].
Результаты опытов показали, что как отдельное действие засоления, так и ощелачивания, существенно не отразились на величине отношения хлорофилла а к хлорофиллу б . Отсутствие значительных изменений в величине этого показателя позволяет предположить, что эти факторы не оказывают влияния на светособирающий комплекс тилакоидных мембран данной тест-культуры (рис. 3).
При совместном действии засоления и ощелачивания происходило снижение этого показателя в листьях кресс-салата по сравнению с контролем, что, вероятно, может свидетельствовать о повышении доли хлорофиллов, принадлежащих ССК II.
Одной их функций ССК II является стабилизация мембраной структуры хлоропластов [Бухов, Бондарь, Дроздова, 1998].
Высокие концентрации хлорида натрия вызывают усиление генерации активных форм кислорода (АФК). Они могут образовываться в электрон-транспортной цепи хлоропластов и митохондрий. Окислительный стресс может вызывать перекисное окисление липидов, инактивацию белков и ферментов, нарушение целостности мембран и деградацию хлорофилла [Кения, Лукаш, Гуськов,

Рис. 3 . Влияние засоления и ощелачивания на отношение хлорофилла а к хлорофиллу в
При адаптации растений к действию неблагоприятных факторов среды важное значение имеют такие соединения, как каротиноиды, что связано с их антиоксидантными свойствами [Чиркова, 2002; Mittler, 2002]. Каротиноиды являются обязательными компонентами фотосинтетического аппарата растений. Это полиеновые соединения, имеющие систему сопряженных связей. Они играют роль вспомогательных светособирающих комплексов в той части солнечного спектра, в которой слабо поглощает хлорофилл, выполняют фотопротектор-ную, структурную и защитную функции, а также принимают участие в фотохимических процессах ФС Iи ФС II [Merzlyak, Solovchenko, 2002; Содержание ликопина ..., 2009].
Было установлено, что действие ощелачивания корневой среды, особенно в сочетании с засоляющим фактором, достоверно повысило количество каротиноидов в растениях кресс-салата по сравнению с контрольным вариантом (табл. 2). Повышение содержания этих соединений имеет адаптивное значение, т. к. каротиноидам принадлежит протекторная роль. Они являются компонентами антиоксидантной системы, в функции которой входит защита мембран от повреждающего действия свободных радикалов [Чиркова, 2002; Дымова, Головко, 2007].
Каротиноиды могут также защищать растения от окислительного стресса, изменяя физические свойства фотосинтетических мембран при участии ксантофилового цикла [Кузнецов, Дмитриева, 2005]. Они способны переводить триплетный хлорофилл и синглетный кислород в основное состояние за счет образования триплетного каротиноида, который диссипирует энергию в тепло, препят- ствуя, таким образом, развитию окислительных процессов [Чиркова, 2002].
Из данных табл. 1 видно, что имеется средняя обратная корреляция между высотой (r = –0.65) и массой растений (r = –0.48) с содержанием в них каротиноидов. Более сильная прямая корреляция (r = 0.91) установлена между содержанием каротиноидов и хлорофилла а по сравнению с хлорофиллом в (r = 0.37). Высокая корреляция (r = 0.74) обнаружена между суммарным количеством хлорофиллов и содержанием каротиноидов в растениях кресс-салата ( см. табл. 1).
Учитывая особую роль вспомогательных пигментов фотосинтеза в устойчивости хлорофиллов к фотоокислению, увеличение пула каротиноидов можно рассматривать как один из факторов, обеспечивающих резистентность растений кресс-салата к действию засоляющих ионов и ощелачивания. Повышение уровня каротиноидов указывает на активное функционирование адаптивных механизмов в фотосинтетическом аппарате растений при действии неблагоприятных условий среды.
Выводы
-
1. Засоление корневой среды (0.5%-ный NaCl) оказало ингибирующее влияние на высоту и массу кресс-салата.
-
2. Повышение уровня содержания хлорофиллов а и б при комбинированном действии засоления и ощелачивания, по-видимому, является одной из адаптивных реакций кресс-салата, обеспечивающих резистентность к действию данных неблагоприятных факторов.
-
3. При комбинированном действии засоления и ощелачивания корневой среды более значительно возрастало количество хлорофилла б , соотношение между хлорофиллами заметно снижалось.
-
4. Увеличение пула каротиноидов при отдельном и совместном действии засоления и ощелачивания свидетельствует об активации защитных механизмов антиоксидантной системы растений.
Список литературы Содержание пигментов в растениях Lepidium sativum в условиях хлоридно-натриевого засоления и ощелачивания
- Белецкий Ю.Д., Шевякова Н.И., Карнаухова Т.Б. Пластиды и адаптация растений к засолению. Ростов н/Д.: Изд-во Ростов. ун-та, 1990. 48 с.
- Бухов Н.Г., Бондарь В.В., Дроздова И.С. Действие низкоинтенсивного синего и красного света на содержание хлорофиллов а и б и световые кривые фотосинтеза у листьев ячменя//Физиология растений. 1998. Т. 45, № 4. С. 507-512.
- Влияние гена desA Л12-ациллипидной десатуразы на структуру хлоропластов и устойчивость к гипотермии растений картофеля/Н.В. Астахова, И.Н. Демин, Н.В. Нарайкина, Т.И. Трунова//Физиология растений. 2011. Т. 58, № 1. С. 2127.
- Гавриленко В.Ф., Жигалова Т.В. Большой практикум по фотосинтезу. М.: Академия. 2003. 254 с.
- Гарифзянов А.Р., Горелова С.В. Окислительный стресс и устойчивость растений//Тульский экологический бюллетень. 2006. Вып. 2. С. 364-368.
- Дымова О.В., Головко Т.К. Состояние пигментного аппарата растений живучки ползучей в связи с адаптацией к световым условиям произрастания//Физиология растений. 2007. Т. 54, № 1. С. 47-53.
- Захарин А.А., Паничкин Л.А. Феномен солерезистентности гликофитов//Физиология растений. 2009. Т. 56, № 1. С. 107-116.
- Иванов А.А. Влияние световых условий выращивания пшеницы на чувствительность фотосинтетического аппарата к солевому стрессу//Физиология растений. 2010. Т. 57, № 6. С. 826-834.
- Кения М.В., Лукаш А.И., Гуськов Е.Н. Роль низкомолекулярных антиоксидантов при окислительном стрессе//Успехи современной биологии. 1993. Т. 113, вып. 4. С. 456-470.
- Кузнецов Вл.В., Дмитриева Г.А. Физиология растений. М.: Высш. шк., 2005. 736 с.
- Кусакина М.Г., Еремченко О.З., Четина О.А. Влияние техногенных солей на пигментную систему растений, произрастающих в зоне воздействия солеотвалов//Охрана природной среды и эколого-биологическое образование: материалы III Всерос. с междунар. участием науч.-практ. конф. Елабуга, 2013. С. 60-63.
- Маевская С.Н., Николаева М.К. Реакция антиоксидантной и осмопротекторной систем проростков пшеницы на засуху и регидратацию//Физиология растений. 2013. Т. 60, № 3. С. 351-359.
- Общие системы устойчивости хлопчатника к засолению и высокой температуре: факты и гипотезы/Вл.В. Кузнецов, Б.В. Хыдыров, Б.В. Ро-щупкин, Н.Н. Борисова//Физиология растений. 1990. Т. 37, вып. 5. С. 987-993.
- Палладина Т.А. Роль протонных насосов плазмо-леммы и тонопласта в устойчивости растений к солевому стрессу//Успехи современной биологии. 1999. Т. 119, № 5. С. 451-461.
- Перекисное окисление липидов и антиоксидантаная система защиты в хлоропластах гороха при тепловом шоке/Л.Н. Курганова, А.П. Весолов, Т.А. Гончарова, Ю.В. Синицина//Физиология растений. 1997. Т. 44. № 5. С. 725-730.
- Половникова М.Г., Воскресенская О.Л. Изменение активности компонентов системы антиоксидантной защиты и полифенолоксидазы у газонных растений на разных этапах онтогенеза в условиях городской среды//Физиология растений. 2008. Т. 55, № 5. С. 777-785.
- Сигнальная роль активных форм кислорода при стрессе у растений/В.Д. Креславский, Д.А. Лось, С.Н. Аллахвердиев, Вл.В. Кузнецов//Физиология растений. 2012. Т. 59, № 2. С. 163-178.
- Содержание ликопина и других каротиноидов в плодах томата белорусской и зарубежной селекции/О.В. Булда, Л.А. Мишин, Г.Н. Алексейчук, Н.А. Ламан//Весщ Нацыянальнай акадэмп навук Беларусь Серыя бiялагiчных навук. 2009. № 1. С. 36-41.
- Стрессорный ответ клеток Nicotianasylvestris L. на засоление и высокую температуру. 1. Аккумуляция пролина, полиаминов, бетаинов и сахаров/Н.И. Шевякова, Б.В. Рощупкин, Н.В. Парамонова, Вл.В. Кузнецов//Физиология растений. 1994. Т. 41, № 4. С. 558-565.
- Структура и функции клеток растений при засолении/Б.П. Строгонов и др. М.: Изд-во Наука, 1970. 318 с.
- Удовенко Г.В. Солеустойчивость культурных растений. Л.: Колос. 1977. 215 с.
- Чиркова Т.В. Физиологические основы устойчивости растений: учеб. пособие. СПб.: Изд-во СПбГУ, 2002. С. 58-68.
- Шахов А.А. Солеустойчивость растений. М., 1956. 850 с.
- Якушкина Н.И., Бахтенко Е.Ю. Физиология растений. М.: Владос, 2005.
- Bolwell J.P. Role of active Oxygen species and NO in Plant Defence responses//Cur. Opin Plant.Biol. 1999. Vol. 2, № 4. P. 287-294.
- Foyer C.H., Shigeoka S. Understanding Oxidation stress and Antioxidant Functions to Enhance Photosynthesis//Plant Physiol. 2011. Vol. 155. P. 93-100.
- Hale H.B. Crossadaptation//Environ Res. 1969. Vol. 2, № 2. P. 324.
- Hare P.D., Gress W.A., Van Stader J. Dissecting the Roles of Osmolyte Accumulation during stress//Plant, Cell and Environ. 1998. Vol. 21, № 6. P. 535-553.
- Lu Z., Neumann P. Water-stressed Maize, Barley and Rice seedling Show species Diversity in Mechanisms of Leas Inhibition//I. Exp. Bot. 1998. Vol. 49. P. 1945-1952
- Merzlayak M.N., Solovchchenko A.E. Photostability of Pigments in Ripening Apple Fruit: A possible photoprotective Role of Carotenoids during Plant senescence//Plant Sci. 2002. Vol. 163. P. 881888.
- Mittler R. Oxidative stress, Antioxidants and Stress Tolerance//Trends Plant Sci. 2002. Vol. 7. P. 405-410.
- Patakas A., Noitsakis B. Mechanisms involved in diurnal changes of asmotic potential in grapevines under drought conditions//Vitis: Viticulat and Enol. Abstr. 2000. Vol. 39, № 1-2. P. 14.
- Vermaas W. Molecular-Biological Approaches to Analyze Photosystem IIstructure and function//Annu. Rev. Plant Physiol. Plant Molecul.Biol. 1993. Vol. 44. P. 457-481.
- Удовенко Г.В., Семухина Л.А., Петроченко Н.Г. Характер и вероятные причины изменения фотосинтезирующей деятельности при засолении//Физиология растений. 1971. Т. 18, вып. 4. С. 708-715.
