Содержание продуктов окислительной модификации белков в плазме крови беременных и лактирующих самок кролика европейского (Oryctolagus cuniculus)
")
Автор: Тарасов Сергей Сергеевич, Корягин Александр Сергеевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Биохимия
Статья в выпуске: 2, 2017 года.
Бесплатный доступ
Работа посвящена изучению уровня окислительной модификации белков путём обнаружения в плазме крови кролика продуктов, прореагировавших с 2,4 - динитрофенилгидразонином. Исследована зависимость от сроков сукрольности, лактации, а также одновременной сукрольности и лактации и уровнем окислительной модификации белков как важнейшего показателя окислительного стресса. Показана зависимость от срока беременности, лактации, а также индуктивного влияния беременность + лактация и уровнем окислительной модификации белков в плазме крови кролика. Выявлено у сукрольных самок кролика в первые 3 дня увеличение всех продуктов, образовавшихся в результате окислительной модификации белков с последующим снижением и увеличением в день окрола. У лактирующих животных после окрола наблюдается динамика снижения продуктов окислительной модификации белков. У животных одновременно сукрольных и лактирующих наблюдается сходная динамика, как и у сукрольных самок с более выраженным изменением за исключением дня окрола, в данном случае роста продуктов окислительной модификации белков не наблюдается.
Окислительная модификация белков (омб), оxidative modification of protein (omp), 4 - динитрофенилгидразоны (2, 4-днфг), физиологический стресс, беременность, лактация
Короткий адрес: https://sciup.org/146278226
IDR: 146278226 | УДК: 577.11:577.122
The content of the products of oxidative modification of proteins in the blood plasma of pregnant and lactating famales of European rabbit (Oryctolagus cuniculus)
We studied the level of oxidative modification of proteins by detection of the products reacted with 2,4-dinitrophenylhydrazonine in rabbit blood plasma. We also studied the dependence on the timing of sucronism, lactation, as well as simultaneous “sokrolynosti” and lactation and the level of oxidative modification of proteins as an important indicator of oxidative stress. Our study shows the dependence on the gestation period, lactation, as well as the inductive effect of pregnancy + lactation and the level of oxidative modification of proteins in rabbit blood plasma. We also found the increase of all the products of oxidative modification of proteins in sucrolous female rabbit in the first 3 days with a subsequent decrease and increase in day of spleen. In lactating animals, the dynamics of the decrease in products of oxidative modification of proteins is observed after the birth. In animals, both sucrol and lactating, a similar dynamic is observed, as in the sucrolous females with a more pronounced change, except for the day of spleen. In this case the growth of products of oxidative modification of proteins is not observed.
Текст научной статьи Содержание продуктов окислительной модификации белков в плазме крови беременных и лактирующих самок кролика европейского (Oryctolagus cuniculus)
Введение. Окислительная модификация белков (ОМБ) -изменение конформации белковой молекулы под действием активных форм кислорода (АФК). Структурные изменения белка при ОМБ могут происходить на разных уровнях организации белковой молекулы, вплоть до разрушения первичной структуры. Это связано с природой и концентрацией АФК в клетках, с особенностями устойчивости живого организма (уровень про и антиоксидантных систем, протеолитических ферментов, биосинтетических систем).
АФК вступают во взаимодействие с белковыми молекулами преимущественно через радикалы аминокислот (Дубинина, 2006,
Вартанян, 1990), но также могут вступать в реакцию с карбоксильной группой, аминогруппой, разрушать ковалентную связь (-СО - NH-) между аминокислотами (Davies, Delsignor, 1987).
ОМБ важнейший показатель окислительного стресса т.к. именно белки являются первичными ловушками АФК (Gebichi, 1997, Singh, Vadasz, 1983). Уровень ОМБ зависит от общего состояния организма, от факторов среды, что может усиливать деструктивные процессы.
Физиологическая беременность, лактация и одновременная совокупность этих процессов является важнейшим фактором меняющим общее состояние животного (Harcourt-Brown, 2002). В литературе имеется ряд данных о влиянии беременности на физиологические процессы (Бурмистров и др., 2004, Шевелькова и др., 2015). В связи, с чем важно изучить влияние беременности, лактации и одновременной беременности и лактации на уровень ОМБ тем самым установить силу данного стресс фактора. Особо важным и актуальным исследованием в нашей работе является изучение влияние кофакторов беременности и лактации на организм животного, что представляет интерес, как с клинической точки зрения, так и с сельскохозяйственной. В связи с чем целью работы явилось: установить зависимость окислительной модификации белков плазмы крови кролика от стадий физиологической беременности, лактации и одновременной беременности и лактации.
Методика. Объектом исследования являлись вторично сукрольные, лактирующие и одновременно сукролные и лактирующие самки кролика европейского породы советская шиншилла в возрасте 10-12 месяцев. Животных содержали на экспериментальной ферме компании «ВитАс», кормили комбикормами концентратами и сухим грубым кормом (сено), поили водой, еда и вода была у животных всегда, т.е. они потребляли пищи столько, сколько была их потребность. В эксперименте было задействовано 9 самок, по три в каждой группе.
Забор крови проводили из ушной вены на 1, 3, 7, 14, 21 день у беременных самок и в день окрола. В случае изучения лактации через 1,3, 7, 14и21 день, а при изучении индуктивного влияния лактации и сукрольности на процессы ОМБ через 1, 2, 3, 7, 14, 21 день и в день окрола.
Определение карбонильных производных, образовавшихся в результате окислительной модификации белков, в плазме крови кролика проводили по модифицированной методике Дубининой. Её принцип основывается на реакции взаимодействия окисленных аминокислотных остатков белков с 2,4 - динитрофенилгидразином (2,4 - ДНФГ) с образованием производных 2,4 - динитрофенилгидразона (Дубинина и др., 1995).
Общий белок определяли биуретовым методом (метод Кингслея
- Вейксельбаума) (Мельников, 1987).
Статистическую обработку полученных результатов производили с помощью программы Microsoft Excel 2010 и Биостатистика вер. 4.03 методами параметрической статистики, включающей - определение средней арифметической (М) и стандартного отклонения. В таблицах и на графиках представлены средние значения и стандартные отклонения не менее чем 3 биологических повторностей, с 3 биохимическими повторностями в каждой из них. Достоверность различий оценивали по t критерию Стьюдента с поправкой Бонферрони. Уровень значимости достоверности различий - 95% (Гланц, 1999).
Результаты и обсуждение. Обнаружены все производные 2,4-ДНФГ в плазме крови опытных и контрольных животных. Эксперимент показал существенное, статистически значимое (Р<0,05) изменение в содержании продуктов окислительной модификации белков в плазме крови всех исследуемых животных. Так основное количество образовавшихся динитрофенилгидразонов относится к альдегидо и кетонпроизводным нейтрального характера. Статистически значимой разницы в их содержании не выявлено (рис. 1-3), за исключением фазы «восстановления» у сукрольных самок и одновременно сукрольных и лактирующих самок (рис. 1, 3). Альдегидо и кетонпроизводные основного характера в исследуемых образцах были ниже, примерно в 1,5 раза, чем те же производные нейтрального характера (рис. 1-3). Существенной разницы в их содержании так же нет, однако содержание альдегидпроизводных основного характера на стадиях тревоги, у беременных и одновременно беременных и кормящих самок, показали статистически значимое увеличение (Р<0,05), чем содержание кетонпроизводных того же характера.
На рисунке 1 показана зависимость срока беременности самки кролика от содержания продуктов ОМБ. Установлено, что в первые 3 дня после спаривания количество продуктов ОМБ увеличивается по отношению к контролю (Р<0,05). При дальнейшем изучении динамики ОМБ на разных стадиях беременности наблюдается снижение продуктов ОМБ (7, 14 и 21 день исследований). В день окрола у самок зафиксировано резкое увеличение продуктом ОМБ как по отношению к предыдущему измерению, так и по отношению к контролю (Р<0,05). Вероятно, это связано с физиологическими и биохимическими изменениями в организме животного, что по видимому усиливает генерацию АФК и приводит к стрессовому состоянию (Владимиров, 1998, Дубинина, 2006).
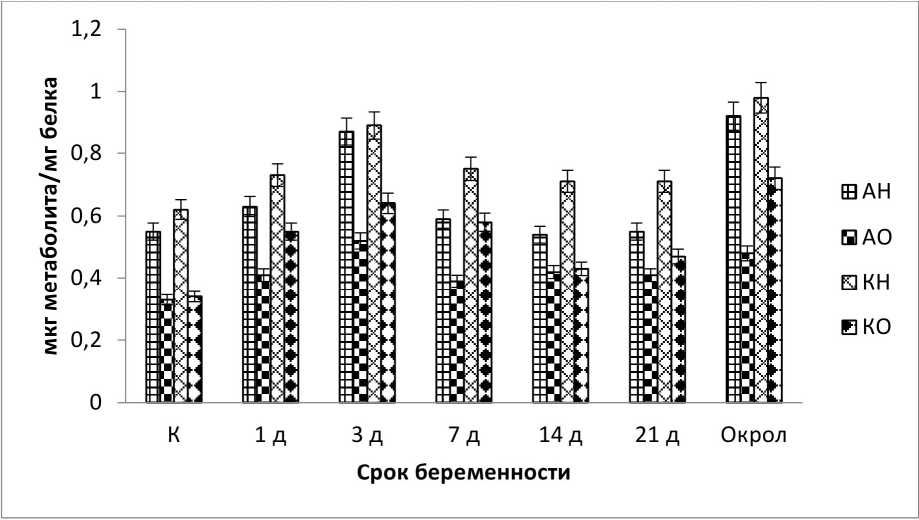
Рис. 1 . Содержание продуктов ОМБ в плазме крови самок кролика на разных стадиях беременности: АН - алифатические альдегид-денитрофенилгидразоны нейтрального характера, АО - алифатические альдегид-денитрофенилгидразоны основного характера, КН - алифатические кетон-денитрофенилгидразоны нейтрального характера, КО - алифатические кетон-денитрофенилгидразоны основного характера
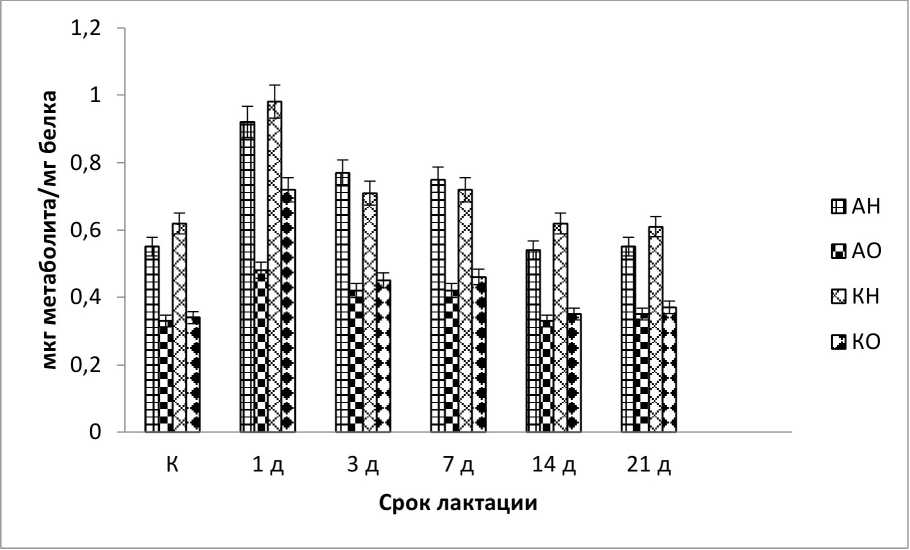
Рис. 2 . Содержание продуктов ОМБ в плазме крови кролика на разных стадиях лактации самки (обозначения см. рис. 1)
Анализируя концентрации продуктов ОМБ в плазме лактирующих самок кролика, видно, что первый день лактации (день окрола) зафиксировано резкое повышение уровня ОМБ, но уже на 3-й день лактации наблюдается снижение всех производных 2,4 - ДНФГ (Р<0,05), особенно заметно снижение основных кетонов, основные же альдегиды изменяются слабо. В последующие дни измерения фиксировалась стабильная динамика снижения уровня ОМБ (рис. 2). Это можно объяснить тем, что после стрессового состояния организм самок постепенно начинает приходить в норму, а так же тем, что процессы лактогенеза сопровождаются повышенным содержанием антиоксидантных ферментов (Бурмистров, 2004).
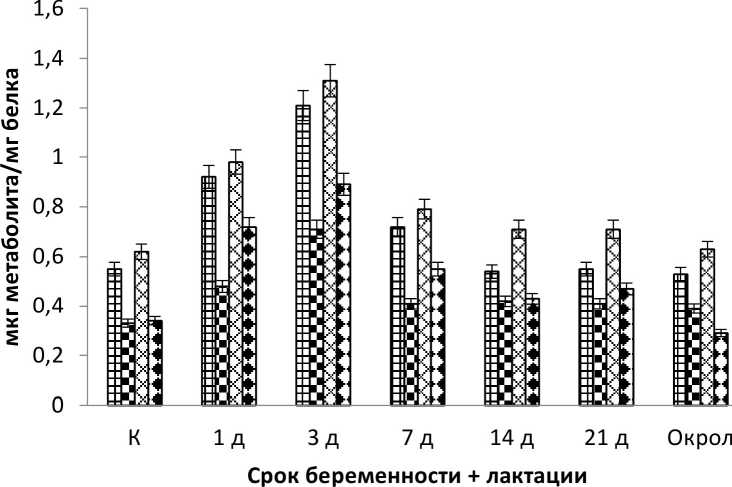
Рис. 3 . Содержание продуктов ОМБ в плазме крови кролика на разных стадиях беременных и одновременно лактирующих самок (обозначения см. рис. 1)
Кролик европейский типичный гиперпродуктивный вид млекопитающих, способный одновременно вынашивать и выкармливать молодняк. Даная способность актуальна лишь в высоко благоприятных условиях (Балакирев и др., 2007). При исследовании зависимости уровня ОМБ и индуктивного эффекта сукрольность + лактация выявлено существенное изменение концентраций продуктов ОМБ в плазме крови кролика. Установлено, что у самок в день окрола, после спаривания и оплодотворения концентрация продуктов ОМБ увеличивается по отношению к контролю (Р<0,05) выше чем у самок без спаривания (рис. 3). На 3-й день зафиксирован больший рост процессов ОМБ по отношению к 1-му дню, что свидетельствует об активной генерации АФК и усилению окислительного стресса. Однако на 7-й день измерения продуктов ОМБ зафиксировано резкое снижение деструктивных процессов, а содержание продуктов ОМБ фактически приближалось к контролю. Последующие измерения (14, 21-й дни) показали положительную динамику снижения некоторых фракций ОМБ. В день окрола уровень ОМБ статистически значимо (Р>0,05) по отношению к предыдущим измерениям не отличался, за исключением основных кетонов, которые показали динамику к понижению. Таким образом, окрол у самок с индуктивным эффектом сукрольность + лактация не сопровождается генерацией АФК и соответственно процессы ОМБ не фиксируются. Данные результаты вероятно можно объяснить адаптационным эффектом животного, который при повышенных физиологических нагрузках первое время не способен сопротивляться побочным продуктом - АФК, но при благоприятных условиях происходит генерация веществ подавляющих АФК и как следствие снижение процессов ОМБ.
Выводы: 1. Содержание продуктов окислительной модификации белков (ОМБ) у сукрольных самок кролика увеличивается в течение первых 3-х дней беременности, с последующим понижением и резко возрастает в день окрола.
Концентрация продуктов окислительной модификации белков (ОМБ) у лактирующих самок кролика снижается с момента окрола на протяжении всего исследуемого периода лактогенеза.
Самки кролика одновременно сукрольные и лактирующие показали резкое увеличение продуктов окислительной модификации белков в плазме крови в первые 3 дня с последующим снижением, во время окрола концентрация данных продуктов не превышала контроль.
КОРЯГИН Александр Сергеевич - доктор биологических наук, профессор, ФГБОУ ВПО «Нижегородский государственный университет им. Н.И. Лобачевского», 603022, Нижний Новгород, пр. Гагарина, 23.
Тарасов С.С. Содержание продуктов окислительной модификации белков в плазме крови беременных и лактирующих самок кролика европейского lOryctolagus cuniculus^ / С.С. Тарасов, А.С. Корягин // Вести. ТвГУ. Сер.: Биология и экология. 2017. №2. С. 102-110.
Список литературы Содержание продуктов окислительной модификации белков в плазме крови беременных и лактирующих самок кролика европейского (Oryctolagus cuniculus)
- Балакирев Н.А., Тинаева Е.А., Тинаев Н.И., Шумилина Н.Н. 2007. Кролиководство. М.: КолосС. 232 с.
- Бурмистров О.С., Опарина Т.И., Арутюнян А.В. 2004. Изучение чувствительности белков сыворотки крови к окислению при беременности//Журнал акушерства и женских болезней. Т. LIII. № 3. С. 11-13.
- Вартанян Л.С. Рашба Ю.Э., Наглер Л.Г. Згурский А.А. Октябрьская Л.А. Биленко М.В. 1990. Мембраны субклеточных органелл как источник супероксидных радикалов при ишемии печени//Бюллетень экспериментальной биологической медицины Т. 109. № 6. С. 550-552
- Владимиров Ю.А. 1998. Свободные радикалы и антиоксиданты//Вестн. РАМН. № 7. С. 43-51.
- Гланц С. 1999. Медико-биологическая статистика М.: Практика. 459 с.
- Дубинина Е.Е. 2006. Продукты метаболизма кислорода в функциональной активности клеток. СПб. 396 с.
- Дубинина Е.Е., Бурмистров С.О., Ходов Д.А., Поротов И.Г. 1995. Окислительные модификации белков сыворотки крови человека, метод ее определения//Вопросы медицинской химии. Т. 41. № 1. С. 24-26.
- Мельников В.В. 1987. Лабораторные методы исследования в клинике. М.: Медицина. 367 с.
- Шевелькова А.А., Айламазян Э.К., Евсюкова И.И. 2015. Особенности окислительной модификации белков и функционального состояния эритроцитов при беременности, осложнённой хронической плацентарной недостаточностью//Журнал акушерства и женских болезней Т. 64. № 5. С. 62-68.
- Davies K.J.A., Delsignor M.E. 1987. Protein damage and degradation by oxygen radicals. III. Modification of secondary and tertiary structure//J. Biol. Chem. Vol. 262. № 20. P. 9908-9913.
- Gebichi J.M. 1997. Protein hydroperoxides as new reactive oxygen species//Redox Rep. Vol. 3. № 2 P. 9-110.
- Harcourt-Brown F. 2002. Textbook of Rabbit Medicine. Oxford, UK: Butterworth Heinemann.
- Singh H., Vadasz J.H. 1983. Effects of gamma radiation E. coli II. Efficienciec of inactivation by free radicals//Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. Vol. 44, № 6. P. 607-613.
