Сообщества беспозвоночных и микроорганизмов в почвах полей фильтрации Льговского сахарного завода (Курская область)
")
Автор: Рахлеева А.А., Михеев П.В., Замотаев И.В., Грачева Р.Г., Конопляникова Ю.В., Карелин Д.В., Белоновская Е.А.
Журнал: Бюллетень Почвенного института им. В.В. Докучаева @byulleten-esoil
Рубрика: Статьи
Статья в выпуске: 121, 2024 года.
Бесплатный доступ
Изучены особенности состава макро- и микробиоты в почвах, сформированных в условиях очистных сооружений (полей фильтрации) сахарной промышленности в черноземной зоне России. Значительное влияние на сообщества микроорганизмов и беспозвоночных оказывают влажность почв, обилие и разнообразие органических и минеральных веществ, поступающих на очистные сооружения со сточными водами. Макрофауна почв периодически действующих прудов-отстойников (карт) представлена преимущественно хищниками и фитофагами; в карте, выведенной из эксплуатации 45 лет назад, преобладают сапрофаги, в составе которых доминируют дождевые черви. Все группы макрофауны в почвах полей фильтрации характерны для почв лесостепных регионов, при этом их численность существенно ниже, по сравнению с черноземами под дубравами Центрально-Черноземного заповедника, но заметно выше, чем в окружающих пахотных черноземах. Микрофлора в почвах полей фильтрации и естественном черноземе Курской биосферной станции идентична по составу физиологических групп, но различается по их количеству и распределению по профилю, что связано с разным водным режимом, определяемым частотой и длительностью воздействия сточных вод, и поступлением отходов сахарного производства, обогащенных карбонатом кальция, органическим веществом и биофильными элементами.
Отходы свеклосахарного производства, карты полей фильтрации, макрофауна, микрофлора, technosols, cambisols, chernozems
Короткий адрес: https://sciup.org/143184038
IDR: 143184038 | УДК: 631.4 | DOI: 10.19047/0136-1694-2024-121-152-199
Communities of invertebrates and microorganisms in the soils of filtration fields of the Lgov Sugar Factory (Kursk region)
The composition of macro- and microbiota in soils formed in treatment facilities (filtration fields) of the sugar industry in the Chernozem zone of Russia was studied. Soil moisture and the abundance and diversity of organic and mineral substances entering treated plants with wastewater have a significant impact on communities of microorganisms and invertebrates. The macrofauna of the soils of periodically operating settling ponds (called checks, or carts) consists mainly of predators and phytophages; the soil of the checks that was decommissioned 45 years ago consists of saprophages, which were dominated by earthworms. All groups of macrofauna in the soils of filtration fields are characteristic of the soils of forest-steppe regions, while their numbers are significantly lower compared to the Chernozems under the oak groves of the Central Chernozem Reserve, but noticeably higher than in the surrounding arable Chernozems. The microflora in the soils of the filtration fields and the background Haplic Chernozem (Pachic) is identical in the composition of physiological groups, but differs in their number and distribution along the soil profile. Differences in the diversity and distribution of microorganisms in the studied soils depend on different water regimes, determined by the frequency and duration of exposure to wastewater and the input of sugar production waste enriched with calcium carbonate, organic matter and biophilic elements.
Текст научной статьи Сообщества беспозвоночных и микроорганизмов в почвах полей фильтрации Льговского сахарного завода (Курская область)
119991, Москва, Ленинские горы, 1, стр. 12, * , e-mail:
2ФБУН “ФНЦГ им. Ф. Ф. Эрисмана” Роспотребнадзора, Россия, 141014, г. Мытищи, ул. Семашко, д. 2, **, e-mail:
119017, Москва, Старомонетный пер., д. 29, ***, e-mail: , ****, e-mail: , *****,
e-mail: , ******, e-mail: , *******, e-mail:
12, 1 Leninskie Gori, Moscow 119234, Russian Federation, *, e-mail:
2 Semashko Str., Mytishchi 141014, Russian Federation, **, e-mail:
29 Staromonetniy lane, Moscow 119017, Russian Federation, ***, e-mail: , ****, e-mail: , *****,
e-mail: , ******, e-mail: , *******, e-mail:
Поля фильтрации (в дальнейшем ПФ), используемые для естественной биологической очистки сточных вод сахарных заводов, состоят из карт (выемок) с межсекционными насыпными валами, включающими различные технические элементы. ПФ действующего Льговского сахарного завода (площадь 175 га) расположены на водораздельной поверхности рек Бык и Опока, сложенной четвертичными отложениями. Карты действуют в режиме периодического заполнения сточными водами и осушения. Отдельные карты могут забрасываться, когда объем производства недостаточен для заполнения всей площади ПФ.
Ранее проведенные исследования (Замотаев и др., 2022, 2023; Карелин и др., 2023) показали, что в искусственно созданных картах, окруженных насыпными валами (их высота 1.5–4 м, ширина до 3 м), создается особый комплекс условий, отличный от окружающего фона (Мониторинг…, 2021). К таким особым условиям карт относятся: а) более мощный и продолжительный снежный покров, что обуславливает более высокую температуру почвы в зимний период; б) пониженная температура (на 1 .7 °С, в среднем) и повышенная влажность (27.8 > 18.8% (фон) в слое почвы 0– 10 см, тест Манна–Уитни, р < 0.05) в вегетационный период, с апреля по август; в) более высокая температура воздуха и почвы в осенний период, с сентября по октябрь; г) периодическое обводнение используемых карт с дополнительным поступлением со сточными водами в смеси с дефекатом и транспортерно-моечным осадком биогенных и других веществ, применяемых в свеклосахарном производстве (органические соединения, карбонаты, фосфаты, питательные элементы, технологические вспомогательные средства и др.); д) доминирование гидрофильных и гигрофильных растительных сообществ, не характерных для фоновых лесостепных ландшафтов; е) почвообразующими породами на ПФ являются высохшие осадки сточных вод многокомпонентного состава и карбонатные лессовидные суглинки и/или нижние горизонты черноземных почв, трансформированные техногенными стоками. Все это формирует в картах ПФ особые условия существования сообществ почвенных беспозвоночных и микроорганизмов, не характерные для региональных природных местообитаний.
Беспозвоночные и микроорганизмы, достигая в почвах высоких значений численности и биомассы, особенно в верхних горизонтах, существенно влияют на трансформацию органического вещества и минерального компонента, переорганизацию почвенной массы, а также регулируют циклы биогенных элементов (Гиляров, Криволуцкий, 1985; Микроорганизмы…, 1989; Всеволодо-ва–Перель, 1997; Мордкович, 1991, 2013; Андрусевич и др., 2014; Кутовая и др., 2016; Мордкович, Любечанский, 2020; Рахлеева, 2022; Lavelle et al., 2016; Bayon et al., 2017; Coggan et al., 2018). Для ряда групп почвообитающих животных Черноземья списки видов и сведения об их экологии требуют дальнейшего изучения (Щеглов и др., 2006; Снегин, Артемчук, 2018; Мордкович, Любе-чанский, 2020; Кутовая и др., 2021). Информация по микроорганизмам в черноземной зоне в основном относится к сравнению структуры и численности сообществ в почвах разных типов и подтипов, оценке влияния на них локального переувлажнения (Козеев и др., 2006), различных видов землепользования, севооборотов, приемов обработки и удобрений (залежь, пашня, многолетние полевые опыты, применение технологии no-till), изучению состояния микробиоты при загрязнении почв нефтью (Возняковская и др., 1996; Пухова и др., 2011; Железова и др., 2017; Кутовая и др., 2021; Никитин и др., 2020; Минникова и др., 2023). Известны некоторые биологические параметры субстратов ПФ и агропочв при внесении в них отходов свеклосахарного производства в чистом виде или в сочетании с удобрениями (Иванов, 2004; Пузанова, 2009; Коноплина, 2011; Балабко и др., 2014; Гурин, Гнеушева, 2018). Вместе с тем практически отсутствуют исследования состояния комплексов микроорганизмов и беспозвоночных животных, населяющих почвы территорий, вовлеченных в свеклосахарное производство.
Цель исследования – изучение особенностей состава биоты в почвах на ПФ сахарной промышленности в черноземной зоне.
ОБЪЕКТЫ И МЕТОДЫ
Для исследований были выбраны почвы ПФ Льговского сахарного завода (Курская область) и, для сравнения, агропочва прилегающей территории. Морфологическое описание почвенных разрезов проведено с использованием цветовых таблиц Манселла. Твердость почв определяли с помощью пенетрометра, плотность – бура конструкции Качинского. В лабораторных условиях проводились мезоморфологические исследования с использованием оптического микроскопа Leica MZ6 (рабочие увеличения – ×6–×80).
Почвенные анализы выполнены в химической лаборатории Института географии РАН. Гранулометрический состав определяли пирофосфатным методом, рН водн – потенциометрическим, количество карбонатов – методом алкалиметрического титрования (Аринушкина, 1970; Вадюнина, Корчагина, 1986; Воробьева, 1998).
Содержание углерода и азота измеряли методом сухого сжигания на анализаторе в Центре коллективного пользования (ЦКП) “Лаборатория радиоуглеродного датирования и электронной микроскопии” ИГ РАН.
Количественный учет макрофауны (макрофауна – принятое размерное деление по международной классификации, мезофауна 2–30 мм – по отечественной размерной классификации, введенной М.С. Гиляровым) проводился на трех пробных участках в сентябре 2022 г.: в днищах действующей карты, освободившейся от воздействия сточных вод 2 года назад, и карты, заброшенной 45 лет назад (разрезы LG-01-22 и LG-05-20 соответственно), а также на пашне под озимой пшеницей, сопредельной с ПФ (разрез LG-0422; условный агрофон).
Для количественных учетов макрофауны использовался метод ручной разборки проб по Гилярову (Методы почвеннозоологических исследований, 1975). На каждом пробном участке в трехкратной повторности отбирались почвенные монолиты площадью 25 см 2 , на глубину 0–10, 10–20, 20–30 см, а также подстилка при наличии. Были получены значения численности и биомассы макрофауны с пересчетом на 1 м 2 . Определение состава почвенной макрофауны проводилось до уровня семейств для кольчатых червей и жесткокрылых. Для остальных представителей – до уровня классов и отрядов. Для большинства таксонов был определен их трофический статус: хищники, фитофаги, сапрофаги.
Муравьи в анализе не учитывались. Для этой социальной группы беспозвоночных существуют иные методы количественных учетов. Пустые раковинки гастропод в расчеты численности и биомассы не включались, однако их присутствие отмечалось в общей интерпретации результатов.
На каждом пробном участке был определен доминантный комплекс макрофауны (таксономические группы, чья численность составляла 5% и более от общей численности на участке).
Для сравнительного анализа состава комплексов макрофауны рассчитывался коэффициент Жаккара:
Кж=С/(A+B-C)*100%, где А – число таксонов на одном пробном участке, B – число таксонов на втором пробном участке, С – число общих для двух участков таксонов.
Необходимо учитывать, что пробы были отобраны во время периода сильного иссушения почв. Лето 2022 г. выдалось жаркое, особенно июль и август. Этот фактор мог повлиять на количественные показатели макрофауны, которая в такие периоды проявляет меньшую активность.
Общее сравнение полученных результатов проводилось с опубликованными для Центрально-Черноземного заповедника имени Алехина данными по комплексам макрофауны (Злотин, Ходашова, 1974: цит. по Мордкович, Любечанский, 2020), а также с данными, полученными для почв зонального ряда Русской рав- нины (Гиляров, Чернов, 1975; Мордкович, Любечанский, 2020).
Отбор проб для микробиологических исследований проводился по генетическим горизонтам (р. LG-01-22 и LG-05-20). Биологическую активность образцов почв исследовали методами посевов на элективные среды. Общее микробное число (ОМЧ) подсчитывали на чашках Петри при посеве разведений почвенной суспензии на ГРМ-агаре (ОМЧ1 – потенциально патогенные бактерии, обладающие высокой скоростью роста, выросшие за 24 часа в термостате при 37 °С), а также при инкубации в течение 3 суток при 22 °С на разбавленном в 10 раз ГРМ-агаре (ОМЧ2 – сапрофитные бактерии, т. е. господствующая группа, существующая в почве в отсутствие свежих питательных веществ). Актиномице-ты выявляли на крахмало-аммиачном агаре, плесневые грибы – на среде Чапека, число спор аэробных бактерий – на ГРМ-агаре после прогрева почвенной суспензии на водяной бане в течение 15 минут при температуре 75 °С. Полученные результаты сравнивали с опубликованными ранее для ненарушенных черноземов Курской биосферной станции (Mikheev et al., 2021).
Видовой состав бактерий наиболее распространенных видов, растущих в виде колоний на поверхности агара с почвенной вытяжкой, определяли с использованием масс-спектрометрии (MALDI-TOF MS), являющейся одной из самых последних систем идентификации микроорганизмов, для получения “отпечатков пальцев интактных бактериальных клеток” (Баранцевич, Баранце-вич, 2014 ; Holland et al., 1996; Giebel et al., 2010).
Классификационное положение почв определено в формате новой классификации и диагностики почв России (2004) и Полевого определителя почв России (2008), с дополнениями по классификационной разработке (Прокофьева и др., 2014), а также по международной классификации WRB (IUSS Working Group WRB, 2015). Следует отметить, что при описании почв ПФ использован малый индекс “zoo”, не выделяемый в новой классификации почв России, для обозначения многочисленных и разнообразных признаков биогенной проработки материала в профиле.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфологические свойства почв. Ниже приведены основные морфологические особенности почв и их отличительные черты, связанные со свеклосахарным производством (рис. 1).
В днище карты, которая последний раз заливалась сточными водами в 2020 г., под эхиноцистисом дольчатым ( Echinocystis labata ) (“бешеный огурец”) описана темногумусовая техногенная арти-стратифицированная почва на абраземе аккумулятивно-карбонатном. Разрез LG-01-22 (51°36.827' N, 35°15.363' E), формула профиля: Оса-AUdc-AU-ТСН1ca,rr-ТСН2ca,rr-ТСН3lc,rr-[BCAmc]-[BCmc]. Согласно WRB (2015), почву можно отнести к Technosol Alcalic Chernic Calcariс.
I О1cа – 2–0 см – сухие растительные остатки, преимущественно фрагменты тростника; примесь минерального известкового материала, бурно вскипающего от НСl.
I О2са – 0–5 см – 10YR 2/2, сухая рыхлая рассыпающаяся механическая смесь растительных остатков, семян и мелкозернистого черного минерального материала. Бурно вскипает от НСl.
I AUdc,h – 5–10 см – 10YR 2/1, сухой рыхлый, черный, полностью оструктурен, мелко-среднезернистый, суглинистый, липкий. Обилие мелких корешков, частично мертвых. Переход резкий, граница ровная. Бурно вскипает от НСl.
II Стехн AU-ТСН1ca,rr (дефекат) – 10(20)–30 см – 10YR 5/2 – 4/2, неоднородно окрашен: на буровато-сером фоне темносерые гумусированные пятна, разводы; уплотнен, легкосуглинистый, четко ореховатый. Плотный, сильно пористый, в порах черное органическое вещество, часто копрогенное; обилие тонких корешков. Переход резкий, граница ровная. Бурно вскипает от HCl.

Рис. 1. Профили почв, сформированных в днищах карт полей фильтрации (LG-01-22, LG-05-20) и прилегающей пашни (LG-04-22).
Fig. 1. Profiles of soils formed at the bottoms of settling ponds (checks) of filtration fields (LG-01-22, LG-05-20) and adjacent arable land (LG-04-22).
II ТСН2ca,rr (дефекат) – 30–50 см – 10 YR 5/2, сухой светло-серый однородный по составу и цвету, плотный, сильно пористый, поры крупные (1–3–5 мм), поры и трещины заполнены черным органическим веществом; единичные тонкие сухие корни. Распадается на четкие ореховатые отдельности. Бурно вскипает от HCl. Переход резкий по цвету, граница ровная.
III ТСН3lc,rr (органический транспортерно-моечный осадок) – 50–75 см – 7.5 YR 2.5/1, темно-серый с коричневатостью, однородный по цвету, уплотнен, вязкий, липкий, ореховатый, легкосуглинистый, единичные белые точечные карбонатные выцветы (~1 мм), желтоватые карбонатные стяжения (1–5 мм). Переход резкий, граница ровная. Локально слабо кипит от HCl.
IV [BCAmc] – 75(85)–100 см – 10 YR 4/2 + 10 YR 3/1, буровато-серый, участками прокрашен гумусом, плотный, хорошо оструктурен, ореховатый, легкосуглинистый, слегка опесчанен, белые карбонатные пятна, прожилки, трубочки (псевдомицелий). Переход постепенный. Бурно вскипает от HCl.
IV [BCmc] – 100–120 см – 10 YR 5/2, светлее предыдущего, серовато-бурый, очень плотный, ореховатый, легкосуглинистый; обилие псевдомицелия. Бурно вскипает от HCl.
В днище карты, заброшенной 45 лет назад, очищенной от осадков сточных вод, под разреженным покровом клена американского исследована темногумусовая почва на абраземе аккуму-лятивно-карбонатном, разрез LG-05-20 (51°36.902' N, 35°14.913' E), формула профиля: AU1-AU1+B-AUBdc-BС1-BC2q-[BCA3dc,q](WRB: Histic Gleyic Eutric Cambisol (Loamic)).
О фрагментарный – сухой, сохранившие строение ветки и листья.
AU1 – 0–10 см – 10 YR 2/1, сухой, рыхлый, темно-серый, почти черный, зернистый и мелко-, среднекомковатый. Преимущественно копрогенный, суглинистый, пылеватый, липкий. Пронизан тонкими и средними корнями. Переход постепенный, граница ровная. Не кипит от HCl.
AU1+B – 10–20(25) см – свежий, уплотнен. Механическая смесь темно-серого материала (А1) 10 YR 2/2 и серовато-желтовато-бурого материала горизонта B. Темно-серый материал полностью оструктурен, зернисто-мелкокомковатый с присутствием мелкоореховатых отдельностей. Суглинистый, липкий. Желтовато-серо-бурые участки: прокрашены точечно гумусом, сильнопористые, много тонких и средних корней, много червороин, переход заметный, граница неровная. Локально сильно вскипает от HCl (на бурых участках).
AUBdc – 20(25)–35(40) см – свежий, плотный. На общем желтовато-буром фоне 10 YR 6/4 размытые и четкие пятна диаметром 3–5 см серого цвета 10 YR 5/3. Преимущественно оструктурен. Крупноореховатый (глыбистый). На правой стенке на 20–25 см прослой черного материала, структура горизонтальной ориентации – грубые плитки/ призмы. Корней мало, мелких и средних. Переход заметный по цвету, граница неровная. Слабо вскипает, локально.
BС1 zoo – 35(40)–60 см – увлажнен, плотный, на желто-буром фоне 10 YR 6/4 темно-серая пятнистость 10 YR 5/3 диаметром 0.5–1 см (червороины). Вертикальные ходы-трубки (трещины) (начинаются с 25 см – идут до нижней части профиля) заполнены темно-серым гумусированным материалом, с копролитами. Диаметр 1.5 см. Крупноореховато-глыбистый, пористый, обилие копролитов в ходах корней и червороинах. Единичные точечные железистые скопления. Переход заметный, по уменьшению корневых ходов и червороин, граница ровная. Локально, слабо вскипает.
BC2 q,zoo – 60–80 см – 10 YR 5/4-6/4, свежий, плотный, желтовато-бурый суглинок, пронизан трещинами (трубками), заполненными копролитами и гумусово-глинистым материалом, пятна червороин диаметром до 1 см, корни средние, единично. Полностью оструктурен, структура крупноореховатопризматическая, четкая. Сильнопористый. Обилие железистых скоплений в “волосяных” трещинах. Единичные железистые пятна на поверхности педов. Переход постепенный, граница ровная. Не кипит от HCl.
BC3 dc,q / [ВСАdc,q] –80–100 см – 10 YR 6/4–6/6, свежий, плотный, желтовато-светло-бурый суглинок (более однородно окрашен), круглые червороины, ходы тонких корней. Структура призматическая и глыбистая, крупная. Суглинисто-пылеватый, пористый. Поры и тонкие ходы корней сильно ожелезнены. Бурно кипит от HCl. Видимых новообразований карбонатов нет.
В пределах пашни, расположенной по периферии очистных сооружений, описан агрочернозем дисперсно-карбонатный квази-глееватый, разрез LG-04-22 (51°36.903' N; 035°14.864' E) формула профиля: PU1(dc)-PU2-AUB1-AUB2dc,tr-BCAdc,tr-BCAdc,q-ВСdc. Согласно WRB (2015), почва отвечает определению Chernozem Loamic Aric Turbic.
PU1(dc) – 0–10 см – 10 YR 3/1-2/1, коричневато-темносерый, свежий, плотный, мелкозернисто-комковатый с глыбистыми агрегатами, частично бесструктурный, суглинистый; много тонких корней. Переход постепенный по цвету и пористости. Слабо вскипает от НСl.
PU2 – 10–28(30) см – 10 YR 2/1, черный свежий плотный зернисто-мелкокомковатый копрогенный суглинистый, тонкие корни, переход ясный граница волнистая. Не вскипает от НСl. Подплужная подошва.
AUB1 – 30–40 см – 10 YR 3/2, свежий буровато-темно-серый пятнистый, на темно-сером фоне бурые пятна 5–10 мм, уплотнен, биотурбированный, ореховато-комковатый копрогенный, тонкие корни, переход ясный, граница ровная. Локально вскипает от НСl (бурые участки).
AUB2dc,tr – 40–50 см – 10 YR 4/3/+3/2, сухой, на желтовато-буром фоне темно-серые гумусированные участки, крупная кротовина (12 см) темно-серая с черными пятнами копролитов. Комковато-ореховатый, нечеткая структура, рассыпается при взятии, суглинистый. На правой стенке четкая кротовина 5–6 см светло-бурая с черными пятнами копролитов. В порах черный органический материал. Тонкие корни. Переход постепенный. Не вскипает от НСl.
BCAdc,tr – 50–70 см – 10 YR 4/3-10 YR 3/2, пятнистый желто-бурый, на буром фоне светло-желтые пятна 1–3 см, темносерые округлые пятна 0.5–10 см; уплотнен, нечетко оструктурен, ореховатый, переход постепенный. Локально вскипает от НСl
BCAdc,q – 70–90 см – 10 YR 4/3-10 YR 5/3, сухой, пестрый, на светло-буром фоне множество мелких ржавых пятен, уплотнен, распадается на призмовидные и ореховатые отдельности, непрочные (рассыпаются при взятии), пористый, в порах железистые трубочки, железистые стяжения и пятна на поверхности педов, на стенках трещин гумусовые кутаны и пропитки. Переход постепенный. Бурно вскипает от НСl.
ВСdc – 90–100 см – 10 YR 6/4-10 YR 5/3, сухой сероватобурый с серыми гумусовыми пятнами; скопления рыхлого гумусированного материала по расширениям трещин, мелкокомковатый преимущественно копрогенный, рассыпчатый; обилие тонких корешков. Бурно вскипает от НСl.
Физические, физико-химические и химические свойства почв. Темногумусовая техногенная арти-стратифицированная почва (LG-01-22 ), сформированная на остаточном материале абра-дированных исходных почв, имеет щелочную реакцию (рН = 7.5– 8.5) и высокое содержание карбонатов во всех горизонтах, диагностируемое по бурному вскипанию мелкозема от НСl. В органогенном (О2са), состоящем из механической смеси растительных остатков, семян и мелкозернистого черного минерального материала, и темногумусовом (AUdc,h) горизонтах с обилием слабо-разложившегося органического вещества, содержание С орг 23.95% и 17.91% соответственно (табл. 1). Почва сильно обогащена азотом во всех горизонтах (C : N = 10–13).
Таблица 1. Некоторые химические и физико-химические свойства почв карт полей фильтрации, выведенных из эксплуатации в разное время
Table 1. Some chemical and physicochemical properties of soils in filtration field checks decommissioned at different times
|
Горизонт |
Глубина образца, см |
рН вод |
С орг , % |
C/N |
CaCO 3 , %, вскипание от HCl |
|
Темногумусовая зоотурбированная почва. Разрез LG-05-20. Заброшенная 45 лет назад очищенная карта |
|||||
|
AU1 |
0–10 |
8.03 |
3.32 |
12 |
1.23 |
|
AU1+B |
10–20(25) |
8.16 |
1.75 |
14 |
1.55 |
|
AUBdc |
20(25)– 35(40) |
8.33 |
1.15 |
11 |
0.39 |
|
BС1 |
35(40)–60 |
8.18 |
0.67 |
11 |
0.27 |
|
BC2 q |
60–80 |
8.07 |
0.50 |
9 |
0.45 |
|
BC3 dc.q |
80–100 |
8.31 |
0.53 |
42 |
10.37 |
Продолжение таблицы 1
Table 1 continued
|
Горизонт |
Глубина образца, см |
рН вод |
С орг , % |
C/N |
CaCO 3 , %, вскипание от HCl |
|
Темногумусовая техногенная арти-стратифицированная почва. Разрез LG-01-22. Карта последний раз использовалась (затоплялась сточными водами) 2 года назад |
|||||
|
О2са |
0–5 |
Не опр. |
23.95 |
10.77 |
Бурно вскипает |
|
AUdc,h |
5–10 |
7.54 |
17.91 |
10.23 |
Бурно вскипает |
|
AU-ТСН1ca,rr |
10–30 |
8.50 |
5.86 |
12.36 |
Бурно вскипает |
|
ТСН2ca,rr |
30–50 |
8.50 |
5.14 |
11.92 |
Бурно вскипает |
|
ТСН3lc,rr |
50–75 |
8.34 |
2.49 |
13.75 |
Бурно вскипает |
|
[BCAmc] |
75–100 |
8.45 |
1.39 |
12.40 |
Бурно вскипает |
|
[BCmc] |
100–120 |
8.35 |
0.98 |
11.37 |
Бурно вскипает |
Периодический привнос на поверхность карт со сточными водами твердофазного материала дефеката и транспортерномоечного осадка приводит к дифференциации гранулометрического состава почв. Поверхностные горизонты имеют среднесуглинистый иловато-крупнопылеватый состав. Горизонт II ТСН2ca,rr на глубине 30–50 см характеризуется более тяжелым гранулометрическим составом (табл. 2). Наблюдающееся здесь повышение содержания физической глины подтверждается максимальным значением плотности на этой глубине – 0.96 г/см 3 по сравнению с рыхлым верхним темногумусовым горизонтом (0.57 г/см 3 ).
При сливе вод и обсыхании днища появляющийся растительный покров и почвенная макрофауна фрагментируют, диспергируют известковый (дефекатный) материал, смешивая его с органическим веществом сточных вод. При мезоморфологическом описании разреза обнаружено, что в горизонтах, сложенных дефе-катом и транспортерно-моечным осадком, многочисленны ходы макрофауны, заполненные органическим веществом; встречаются копролиты различного размера (рис. 2).
Для темногумусовой зоотурбированной почвы характерна щелочная реакция (рН = 8.0–8.3) по всему профилю (табл. 1). В профиле почвы хорошо выражена дифференциация по содержанию карбонатов. В верхних гумусово-аккумулятивных и срединных горизонтах почвы количество карбонатов незначительно (0.27–1.55%, слабокарбонатные, локально слабо вскипают); в нижнем, содержащем мицелярные формы карбонатов, на глубине 80–100 см содержание карбонатов достигает 10.37%, что позволяет отнести горизонт к сильнокарбонатным (бурно вскипает). Содержание органического вещества максимально в поверхностных горизонтах (С орг = 3.32%) и постепенно убывает с глубиной, оставаясь достаточно высоким в срединных горизонтах за счет активных биотурбационных процессов. При микроморфологических исследованиях обнаружены ходы макрофауны (червороины) и корней с перемещением почвенной массы, копролиты различного размера, морфологии и степени сохранности. Соотношение C : N в гумусовых и срединных горизонтах колеблется от 9 до 14, а в нижнем горизонте составляет 42.
Таблица 2. Гранулометрический состав почв карт полей фильтрации, выведенных из эксплуатации в разное время
Table 2. Soil particle-size distribution in the checks of infiltration fields decommissioned at different times
|
Горизонт |
Глубина образца, см |
Содержание фракций (%), размер частиц, мм |
||||||
|
1–0.25 |
0.25–0.05 |
0.05–0.01 |
0.01–0.005 |
0.005– 0.001 |
<0.001 |
<0.01 |
||
|
Темногумусовая зоотурбированная почва. Разрез LG-05-20. Заброшенная 45 лет назад очищенная карта |
||||||||
|
AU1 |
0–10 |
0.18 |
0.49 |
56.3 |
7.31 |
9.7 |
26.02 |
43.03 |
|
AU1+B |
10–20(25) |
0.12 |
0.36 |
56.74 |
8.73 |
8.6 |
25.45 |
42.78 |
|
AUBdc |
20(25)–35(40) |
0.11 |
0.87 |
58.89 |
7.97 |
7.39 |
24.77 |
40.13 |
|
BС1 |
35(40)–60 |
0.03 |
1.5 |
61.05 |
6.24 |
7.92 |
23.26 |
37.42 |
|
BC2 q |
60–80 |
0.09 |
0.26 |
59.3 |
8.78 |
7.11 |
24.46 |
40.35 |
|
[ВСАdc,q] |
80–100 |
0.18 |
0.5 |
57.11 |
8.15 |
9.22 |
24.84 |
42.21 |
|
Темногумусовая техногенная арти-стратифицированная почва. Разрез LG-01-22. Карта последний раз использовалась 2 года назад |
||||||||
|
AUdc,h |
5–10 |
3.07 |
17.89 |
36.76 |
9.79 |
13.52 |
18.97 |
42.28 |
|
AU-ТСН1ca,rr |
10–30 |
0.25 |
8.62 |
34.21 |
28.21 |
16.41 |
12.31 |
56.93 |
|
ТСН2ca,rr |
30–50 |
0.21 |
9.82 |
29.21 |
22.62 |
23.72 |
14.41 |
60.76 |


Рис. 2 . Биогенная проработка дефеката в горизонте AU-ТСН1ca,rr на глубине 10–30 см (LG-01-22).
Fig. 2. Biogenic transformation of the sewage sludge in the AU-TSN1ca,rr horizon at a depth of 10–30 cm (LG-01-22).
Для темногумусовой зоотурбированной почвы характерен по всему профилю среднесуглинистый гранулометрический состав (табл. 2). Отмечено некоторое понижение содержания физической глины (37.42%) на глубине 35(40)–60 см. В мелкоземе темногумусовой почвы, как и в других ранее исследованных почвах днищ карт ПФ (Замотаев и др., 2023) преобладающими являются фракции крупной пыли (0.05–0.01 мм) и илистой фракции (<0.001 мм).
Подробная аналитическая характеристика агрочерноземов Льговского района, к которым относится и описанный разрез LG-04-22 , приведена в предыдущих публикациях (Замотаев и др., 2001, 2016, 2018). Почвы характеризуются тяжелым гранулометрическим составом, отсутствием дифференциации по илу и признаков иллювиирования глины. Содержание гумуса в агротемно-гумусовом гор. PU колеблется от 3.9 до 5.7%. Близкая к нейтральной реакция среды наблюдается в большей части профиля до глубины с максимумом карбонатов, где она резко меняется на щелочную.
Верхняя часть горизонта PU агрочернозема дисперсно-карбонатного (LG-04-22), как показали морфологические исследования, мелкозернисто-комковатая с глыбистыми агрегатами, плотная (плотность 1.3 г/см 3 , твердость 27–30 мм). В нижних горизонтах ВСА проявляются признаки сегрегации железа: в порах железистые трубочки, стяжения и пятна на поверхности педов. На стенках трещин наблюдаются гумусовые кутаны и бурно вскипающие карбонатные пропитки. Все эти признаки вполне типичны для ранее описанных в литературе агрочерноземов Курской области (Афанасьева, 1966; Щеглов, 1999; Караваева, Денисенко, 2009; Базыкина, Овечкин, 2012; Мамонтов и др., 2020; Булышева и др., 2021; Белобров и др., 2023).
Макрофауна в почвах карт полей фильтрации. Почвенная макрофауна формируется представителями различных функциональных групп, являющимися индикаторами адаптации организмов к конкретным экологическим условиям.
Состав макрофауны. В исследованных почвах карт ПФ Льговского сахарного завода, а также прилегающей к ним пашни обнаружена разнообразная почвенная макрофауна. В целом на момент отбора проб выявлено 19 таксонов (табл. 3). Все таксоны характерны для естественных черноземных почв лесостепной зоны (Злотин, Ходашова, 1974: по Мордкович, Любечанский, 2020). Обнаружены кольчатые черви (люмбрициды и энхитреиды), пауки, сенокосцы, улитки – гастроподы, многоножки (хилоподы и диплоподы), личинки чешуекрылых, перепончатокрылых, скорпи-онниц, имаго и личинки жесткокрылых (жужелицы, стафилиниды, щелкуны, долгоносики), личинки двукрылых. Это – представители из 8 классов и 16 отрядов. На момент отбора проб ни на одном из исследованных участков не найдены пластинчатоусые жуки и жуки-чернотелки, характерные для лесостепной зоны. Однако делать выводы об их полном отсутствии нельзя, и эти группы, возможно, будут обнаружены при последующих учетах макрофауны в другие периоды времени в связи с циклами развития и условиями окружающей среды.
По числу таксонов исследуемые почвы карт ПФ, выведенные из использования в разное время, близки между собой. В них установлено по 12 таксонов макрофауны. Однако сходство между участками по таксонам составляет не более 43% по коэффициенту Жаккара. Общие для участков ПФ – это дождевые черви, пауки, мокрицы, диплоподы, улитки-гастроподы, жужелицы, стафилини-ды, личинки двукрылых. В темногумусовой техногенной арти-стратифицированной почве карты ПФ (LG-01-22), которая последний раз использовалась 2 года назад, обнаружены личинки чешуекрылых и скорпионниц. В темногумусовой зоотурбирован-ной почве в заброшенной 45 лет назад карте (LG-05-20) – энхит-реиды, хилоподы, сенокосцы, жуки-долгоносики.
Для обоих участков характерно большое количество улиток ( Gastropoda ). Однако в почве карты, которая затапливалась сточными водами 2 года назад, это живые особи, а на давно выведенной из эксплуатации – пустые раковинки. Улитки-гастроподы активно развиваются поздней осенью и в весенний период, что связано с разложением растительного опада и увлажнением почвы. Их пустые раковинки обычно обнаруживаются в конце летнего периода в верхнем горизонте почвы.
Таблица 3. Состав, численность, биомасса и тип питания почвенной макрофауны исследованных участков
Table 3. Composition, abundance, biomass and type of feeding of soil macrofauna in the studied areas
|
Состав макрофауны |
LG-01-22 |
LG-05-20 |
LG-04-22 |
Тип питания |
|
Класс Sedentaria, отряд Haplotaxida |
||||
|
Семейство: |
||||
|
Lumbricidae ( дождевые черви ) |
3.0 |
18.0 |
2.7 |
сапрофаг |
|
Lumbricidae(p) ( коконы дождевых червей ) |
0.7 |
0.3 |
- |
сапрофаг |
|
Семейство: |
||||
|
Enchytraeidae ( энхитреиды ) |
- |
4.0 |
2.0 |
сапрофаг |
|
Класс Arachnida, отряд Aranea ( пауки ) |
1.7 |
2.3 |
- |
хищник |
|
Класс Arachnida, отряд Opiliones ( сенокосцы ) |
1.0 |
- |
хищник |
|
|
Класс Gastropoda (улитки-гастроподы ) |
11.7 |
+ |
- |
фитофаг |
|
Класс Malacostraca, отряд Isopoda, подотряд Oniscoidea (ракообразные - мокрицы ) |
1.7 |
0.3 |
- |
сапрофаг |
|
Класс Chilopoda, отряд Geophilomorpha, семейство Geophilidae ( многоножки - геофилиды ) |
- |
2.3 |
0.3 |
хищник |
|
Класс Diplopoda ( многоножки - диплоподы ) |
0.7 |
1.7 |
- |
сапрофаг |
|
Класс Insecta: |
||||
|
Отряд Lepidoptera (l) ( чешуекрылые ) |
0.7 |
- |
- |
фитофаг |
|
Отряд Mecoptera (l) ( скорпионницы ) |
0.3 |
- |
- |
хищник |
|
Отряд Hymenoptera (l) ( перепончатокрылые ) |
- |
- |
0.3 |
Продолжение таблицы 3
Table 3 continued
|
Состав макрофауны |
LG-01-22 |
LG-05-20 |
LG-04-22 |
Тип питания |
|
Отряд Coleoptera, семейство Carabidae (i) ( жуки жужелицы имаго ) |
3.3 |
0.3 |
0.3 |
хищник |
|
Carabidae (l) ( жуки жужелицы личинки ) |
0.7 |
0.3 |
- |
хищник |
|
Отряд Coleoptera, семейство Staphylinidae (i) ( жуки стафилиниды имаго ) |
9.3 |
0.7 |
0.3 |
хищник |
|
Staphylinidae (l) ( жуки стафилиниды личинки ) |
0.3 |
- |
- |
хищник |
|
Отряд Coleoptera, семейство Elateridae (i) ( жуки щелкуны имаго ) |
- |
- |
0.7 |
фитофаг |
|
Elateridae (l) ( жуки щелкуны личинки ) |
- |
- |
1.0 |
фитофаг |
|
Отряд Coleoptera, семейство Curculionidae (l) ( жуки долгоносики личинки ) |
- |
1.3 |
- |
фитофаг |
|
Отряд Coleoptera (i)* ( жесткокрылые имаго ) |
0.7 |
- |
- |
|
|
Отряд Coleoptera (i)** ( жесткокрылые имаго ) |
- |
0.7 |
- |
|
|
Отряд Coleoptera (p) ( жесткокрылые куколки ) |
0.3 |
- |
- |
|
|
Отряд Diptera (l) ( двукрылые личинки ) |
- |
2.0 |
1.0 |
|
|
Отряд Diptera (p) ( двукрылые пупарий ) |
0.3 |
- |
0.3 |
|
|
Численность, экз./м2 |
565 ± 400 |
565 ± 70 |
144 ± 73 |
|
|
Биомасса, г/м2 |
26.6 ± 21.4 |
42.6 ± 14.08 |
9.3 ± 7.68 |
Обращает внимание отсутствие хищных многоножек – хи-лопод (семейство геофилиды), в темногумусовой техногенной ар-ти-стратифицированной почве. Эта группа беспозвоночных типична для верхних минеральных горизонтов почв лесостепной зоны. Ее отсутствие может свидетельствовать о влиянии физикохимических параметров, в первую очередь, присутствии не типичных для природных почв химических соединений, поступающих со сточными водами.
В агрочерноземе дисперсно-карбонатном число таксонов снижено до 8 (табл. 3). Здесь отсутствуют пауки, сенокосцы, улитки, мокрицы, диплоподы, личинки чешуекрылых, скорпионниц и жуков-долгоносиков. Общее сходство по составу макрофауны с почвами карт ПФ не более 25% по расчетам коэффициента Жаккара.
Численность макрофауны оказалась сходной в почвах на ПФ – 565 экз./м 2 , что в несколько раз превышает численность в агрочерноземе на участке пашни, где обнаружено только 144 экз./м 2 . Все полученные значения численности ниже показателей, характерных для естественных черноземных почв под лесными формациями Центрально-Черноземного заповедника (рис. 3), где численность может достигать 3 300 экз./м 2 . В то же время они сопоставимы со степным нативным участком заповедника, численность почвенной макрофауны для которого составляет 484 экз./м 2 .
Однако если исключить из анализа численность энхитреид – кольчатых малощетинковых червей (рис. 4), сборы которых методом ручной разборки могут быть неполноценными из-за малых размеров беспозвоночных, неприметной окраски (существуют отдельные методики учетов именно энхитреид), то полученные результаты по почвам ПФ становятся даже выше известных для Центрально-Черноземного заповедника. Численность макрофауны агрочернозема и в таком случае в несколько раз ниже известных данных (Мордкович, Любечанский, 2020).
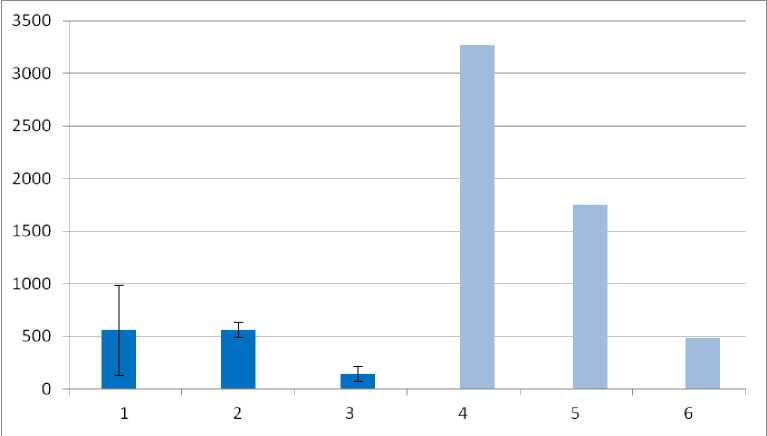
Рис. 3 . Численность почвенной макрофауны (экз./м2). Пробные участки: 1 – LG-01-22, 2 – LG-05-20, 3 – LG-04-22, 4 – дубрава, 5 – лесные луга на полянах, 6 – нативная луговая степь. Участки 4, 5, 6 – Центрально-Черноземный заповедник (Злотин, Ходашова, 1974, по Мордкович, Любечанский, 2020).
Fig. 3. Number of soil macrofauna (individuals/m2). Test plots: 1 – LG-01-22, 2 – LG-05-20, 3 – LG-04-22, 4 – oak grove, 5 – forest meadows, 6 – native meadow steppe. Plots 4, 5, 6 – Central Chernozem Reserve (Zlotin, Khodashova, 1974, after Mordkovich, Lyubechansky, 2020).
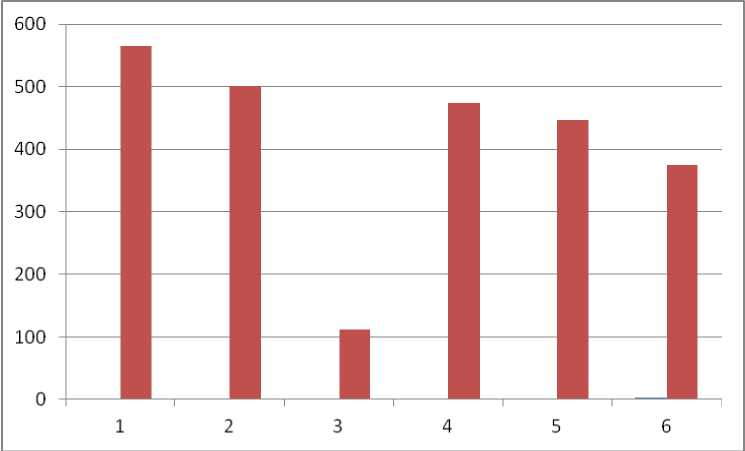
Рис. 4. Численность почвенной макрофауны без учета энхитреид (экз./м2). Пробные участки: 1 – LG-01-22, 2 – LG-05-20, 3 – LG-04-22, 4 – дубрава, 5 – лесные луга на полянах, 6 – нативная луговая степь. Участки 4, 5, 6 – Центрально-Черноземный заповедник (Злотин, Ходашова, 1974; по Мордкович, Любечанский, 2020).
Fig. 4. Number of soil macrofauna excluding enchytraeids (individuals/m2). Test plots: 1 – LG-01-22, 2 – LG-05-20, 3 – LG-04-22, 4 – oak grove, 5 – forest meadows, 6 – native meadow steppe. Plots 4, 5, 6 – Central Chernozem Reserve (Zlotin, Khodashova, 1974; after Mordkovich, Lyubechansky, 2020).
Доминантные комплексы. Анализ доминантных комплексов показал (табл. 4), что, при сходных значениях численности макрофауны почв карт ПФ, набор доминирующих групп, чья численность составляет от 5% и выше от общей на участке, значительно различается. В почвах обеих карт ПФ на момент отбора проб многочисленны дождевые черви и пауки, причем в темногумусовой зоотурбированной почве на заброшенной карте обилие дождевых червей составляет более 50% от общей численности макрофауны на участке. Только в темногумусовой техногенной арти-стратифицированной почве в состав доминантного комплекса входят улитки, мокрицы, жуки жужелицы и стафилиниды. Тогда как в темногумусовой зоотурбированной почве обильное количество составляют многоножки хилоподы семейства геофилид, ди-плоподы, энхитреиды и личинки двукрылых. Набор доминантов в почвах давно неиспользуемой (заброшенной) карты ПФ ближе к агрочерноземам. На этих участках установлены три группы беспозвоночных с высокой численностью – дождевые черви, энхитре-иды и личинки двукрылых. При этом на долю дождевых червей в агрочерноземе пашни и в темногумусовой почве заброшенной карты приходится более 30% от общей численности макрофауны. Кроме того, в агрочерноземе в составе доминантного почвенного комплекса присутствуют жуки щелкуны – типичные обитатели агроценозов; в почвах карт ПФ жуки щелкуны не были обнаружены ни в личиночной стадии, ни в стадии имаго.
Трофическая структура. Если сопоставить обнаруженные на исследуемых участках доминантные комплексы макрофауны с характером питания беспозвоночных (трофические предпочтения), то обнаруживаются следующие особенности. В темногумусовой техногенной арти-стратифицированной почве (LG-01-22) в составе комплекса макрофауны преобладают хищники – 40% и фитофаги – 33% (суммарно более 70%). Сапрофаги составляют только 10%. В темногумусовой зоотурбированной почве (LG-0520) на заброшенной 45 лет назад карте сапрофаги составляют уже 72% (из которых 51% приходится на дождевых червей), доля хищников снижается до 17%, а количество фитофагов не превышает 3%. Это оказывает большое влияние на переорганизацию почвенной массы и трансформацию органического вещества, спо- собствуя процессам гумификации, что подтверждается морфоаналитическими исследованиями и дополняет имеющиеся в литературе материалы.
Таблица 4. Доминантный комплекс макрофауны исследованных участков
Table 4. Dominant macrofauna complex in the studied areas
|
Состав макрофауны |
LG-01-22 |
LG-05-20 |
LG-04-22 |
|
Lumbricidae |
8 |
51 |
30 |
|
Aranea |
5 |
7 |
н. о. |
|
Gastropoda |
33 |
н. о. |
н. о. |
|
Oniscoidea |
5 |
1 |
н. о. |
|
Carabidae (i) |
9 |
1 |
4 |
|
Staphylinidae (i) |
26 |
2 |
4 |
|
Geophilidae |
н. о. |
7 |
4 |
|
Diplopoda |
2 |
5 |
н. о. |
|
Diptera (l) |
н. о. |
6 |
11 |
|
Enchytraeidae |
н. о. |
11 |
22 |
|
Elateridae (i) |
н. о. |
н. о. |
7 |
|
Elateridae (l) |
н. о. |
н. о. |
11 |
Примечание. н. о. – не обнаружены.
Note. н. о. – not found.
В агропочве более 52% численности составляют сапрофаги (прежде всего дождевые черви, а также энхитреиды) и фитофаги – 18%. На долю хищников приходится около 12%, что значительно ниже доли хищников в почвах карт.
Биомасса. По обнаруженным значениям биомассы макрофауны участки карт ПФ в 3–5 раз превышают биомассу на участке пашни и составляют от 20 до 40 грамм на м 2 (рис. 5). Основной вклад в этот показатель на всех участках вносит биомасса дождевых червей. Биомасса макрофауны карт ПФ сопоставима с биомассой макрофауны в черноземе под луговой степью (Гиляров, Чернов, 1975), однако в 2–3 раза ниже, чем для серых лесных почв и черноземов под дубравами.
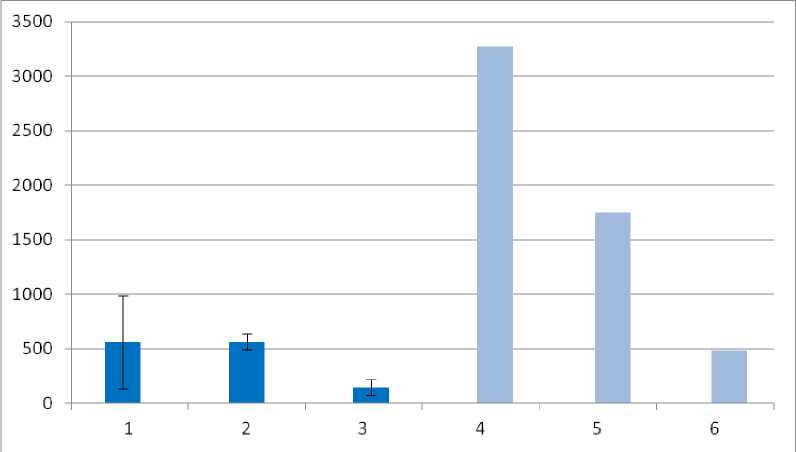
Рис. 5. Биомасса почвенной макрофауны (г/м2). Пробные участки: 1 – LG-01-22, 2 – LG-05-20, 3 – LG-04-22, 4 – дубрава на серой лесной почве, 5 – дубрава на черноземе, 6 –луговая степь на черноземе (участки 4, 5, 6 – Гиляров, Чернов, 1975, по Мордкович, Любечанский, 2020).
Fig. 5. Biomass of soil macrofauna (g/m2). Test plots: 1 – LG-01-22, 2 – LG-05-20, 3 – LG-04-22, 4 – oak grove on gray forest soil, 5 – oak grove on chernozem, 6 – meadow steppe on chernozem (plots 4, 5, 6 – Gilyarov, Chernov, 1975, after Mordkovich, Lyubechansky, 2020).
Микроорганизмы в почвах. В результате микробиологических исследований почв днищ карт выяснилось нетипичное распределение бактерий по профилю по сравнению с фоновыми черноземами Курской биосферной станции (Mikheev et al., 2021). В поверхностных горизонтах – органогенном и темногумусовом (0– 10 см) – техногенной арти-стратифицированной почвы в карте, не используемой в течение 2 лет (LG-01-22) и покрытой в момент описания эхиноцистисом дольчатым ( Echinocystis labata ), установлены близкие высокие величины численности бактерий, обладающих двумя типами стратегий: быстрорастущие сапрофитные и олиготрофные (способные расти в почве с низким содержанием органических веществ) (рис. 6). Вниз по профилю их численность постепенно снижается, и с глубины 30 см (гор. ТСН3lc,rr) начинают преобладать не требовательные к элементам питания олиготрофные бактерии. В органогенном горизонте, состоящем из сухих растительных остатков, преимущественно фрагментов тростника обыкновенного с примесью карбонатного мелкозема, выявлено большое количество споровых форм. Как отмечалось, образцы, взятые для исследования, были отобраны в начале сентября, и возможной причиной увеличения численности споровых форм явилось летнее иссушение верхнего слоя.
Плесневые грибы – микромицеты обнаружены лишь до глубины 30 см от поверхности (рис. 7), что может быть связано как с водным режимом – большим увлажнением нижних горизонтов, так и с локализацией на поверхности почвы опада тростника обыкновенного. Особенностью изученного профиля является максимальная численность актиномицетов в погребенном трансформированном сточными водами абраземе аккумулятивно-карбонатном на глубине 55–100 см. Здесь они, по-видимому, включаются в трансформацию органического вещества погребенного горизонта естественной почвы (рис. 1), уже подвергшегося деструкции в период эксплуатации карты. Как известно, такое распределение не типично для природных почв, в которых вниз по профилю происходит значительное уменьшение количества микроорганизмов.
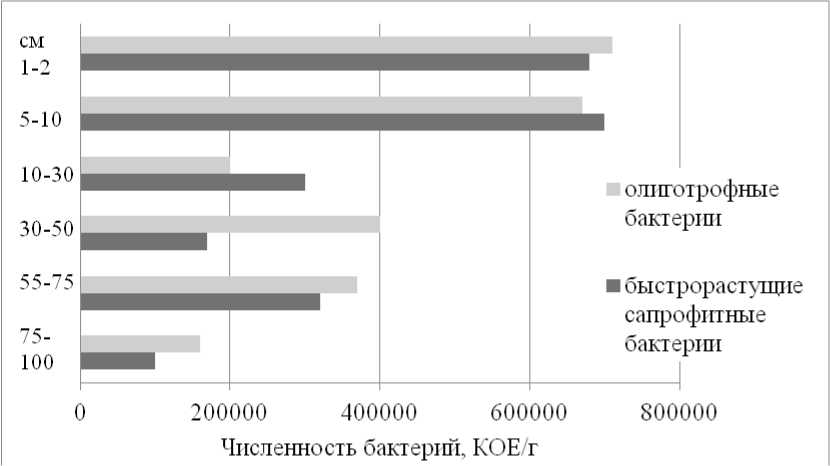
Рис. 6. Численность бактерий в темногумусовой техногенной арти-стратифицированной почве (LG-01-22).
Fig. 6. Number of bacteria in dark humus technogenic artistratified soil (LG-01-22).
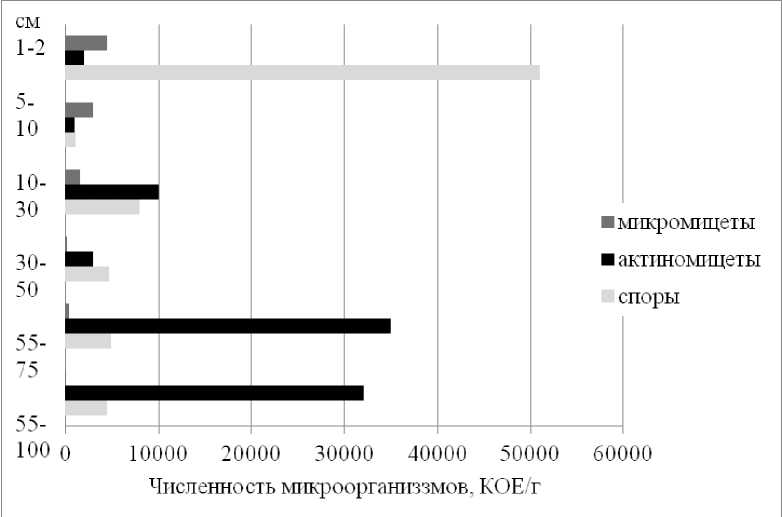
Рис. 7. Численность спор аэробных бактерий, актиномицетов и плесневых грибов в техногенной арти-стратифицированной почве (LG-01-22).
Fig. 7. Number of spores of aerobic bacteria, actinomycetes and mold fungi in technogenic artistratified soil (LG-01-22).
Использование метода MALDI-TOF MS позволило идентифицировать в почве Bacillus marisflavi, Pseudomonas putida и Oceanobacillus kimchii . Присутствовали непатогенные стафилококки в количестве 8 × 10 2 КОЕ/г, непатогенные псевдомонады ( Pseudomonas libanensis, Pseudomonas synxantha ), представители семейства энтеробактерий ( Buttiauxella agrestis, Serratia liquefa-ciens ), а также Aeromonas eucrenophila и энтерококки в количестве 5.0 × 10 2 КОЕ/г.
Изучение распределения численности бактерий по горизонтам профиля (LG-05-20) темногумусовой почвы карты ПФ, которая последний раз заливалась производственными сточными водами 45 лет назад, показало преобладание олиготрофов в верхнем темногумусовом горизонте (рис. 8).
В темногумусовом горизонте обнаружены олиготрофы и споровые формы бактерий (рис. 8, 9), что свидетельствует о не вполне благоприятных условиях для развития микроорганизмов. В переходных подгумусовых горизонтах (AUB) преобладают быстрорастущие сапрофитные бактерии (рис. 8), что связано с высоким содержанием на глубине 10–40 см легко разлагаемого (доступного) органического вещества (С орг = 1.75%). Во всех горизонтах присутствуют плесневые грибы, которые, являясь основными деструкторами сложных органических соединений (лигнин, хитин, дубильные вещества, целлюлоза), делают возможным дальнейшее их использование другими микроорганизмами. Обращает внимание высокая численность актиномицетов во всех горизонтах профиля (рис. 9), что не характерно для природных почв. В верхней части они разрушают растительный опад, а на глубине 30–40 см, по-видимому, выступают деструкторами труднодоступных полимеров в составе органических веществ, поступивших в почву со сточными водами и дефекатом во время использования карты.
Среди видов бактерий присуствовали Stenotrophomonas maltophilia, Achromobacter xylosoxidans, Achromobacter piechaudii, Pseudomonas putida, Acinetobacter calcoaceticus . На глубине 30– 40 см определены Serratia liquefaciens, Buttiauxella sp., Leclercia adecarboxylata . Идентифицированы непатогенные стафилококки, их численность составила до 10 3 КОЕ/г.
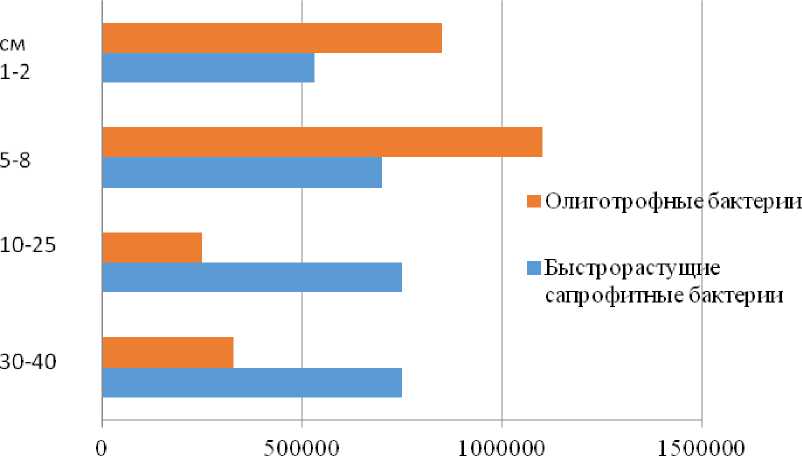
Численность бактерий, КОЕ. г
Рис. 8. Численность бактерий в различных горизонтах темногумусовой почвы (LG 05-20).
Fig. 8. N umber of bacteria in different horizons of dark humus soil (LG 05-20).
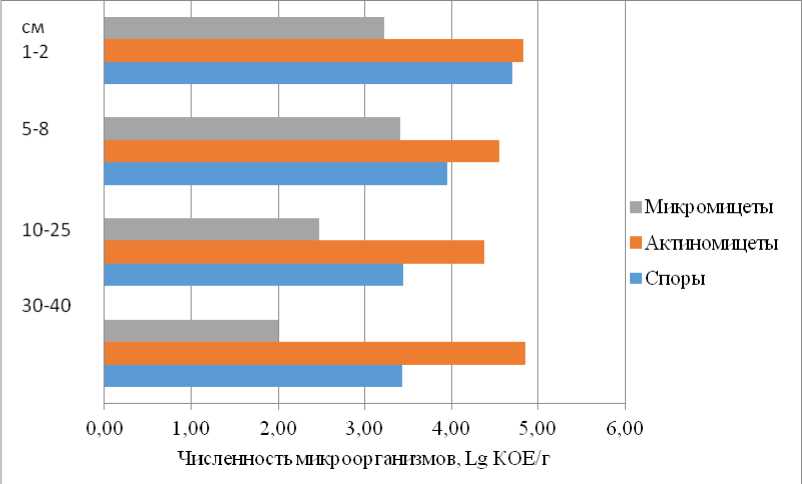
Рис. 9. Численность спор аэробных бактерий, актиномицетов и плесневых грибов в темногумусовой зоотурбированной почве (LG 05-20).
Fig. 9. Number of spores of aerobic bacteria, actinomycetes and mold fungi in dark humus zooturbated soil (LG 05-20).
ЗАКЛЮЧЕНИЕ
Почвы ПФ формируются в специфических природнотехногенных условиях и по своим биологическим свойствам отличаются от природных почв. Значительное влияние на микробиологические сообщества и макрофауну оказывают влажность почв, а также обилие и разнообразие органических и минеральных веществ, поступающих на ПФ со сточными водами.
Все группы макрофауны в почвах ПФ характерны для черноземных почв лесостепных регионов. Однако полученные значения существенно ниже численности макрофауны черноземных почв под дубравами Центрально-Черноземного заповедника, но сопоставимы с биомассой беспозвоночных в черноземе под луговой степью. Этот показатель в значительной степени определяется количеством энхитреид, учет которых в дальнейшем из-за их малых размеров следует продолжить, с использованием специальной методики.
Биомасса макрофауны в техногенной арти-стратифициро-ванной почве периодически действующей карты в 1.5 раза ниже, чем в темногумусовой зоотурбированной почве неиспользуемой карты. Несмотря на схожесть по набору таксонов (43%), у изученных почв ПФ есть ряд принципиальных различий. В почве карты, последний раз заливаемой 2 года назад, в составе комплекса макрофауны преобладают хищники и фитофаги (более 70%) при незначительном количестве сапрофагов. По нашему мнению, в основном это связано с поступлением техногенных стоков, обогащенных органическими веществами и биофильными элементами, что формирует особые условия существования почвенной биоты, не характерные для региональных природных местообитаний. Напротив, в почве давно неиспользуемой карты (45 лет) значительно выше численность сапрофагов (72%), среди них 51% приходится на дождевых червей. О результате активной деятельности последних свидетельствуют многочисленные и разнообразные признаки биогенной проработки материала в профиле на всех морфологических уровнях: разложение опада, биогенное острук-туривание, зоотурбации с формированием червороин, обилие ко-пролитов различного размера, зоогенный перенос материала меж- ду горизонтами.
Микрофлора в почвах ПФ и естественных черноземах ми-грационно-мицелярных Курской биосферной станции идентичны по составу физиологических групп, но различаются по их количеству и распределению в профиле. Особенностью техногенной ар-ти-стратифицированной почвы периодически заливаемой карты является отсутствие плесневых грибов в нижней части профиля, что может быть связано как с водным режимом – большим увлажнением нижних горизонтов, так и с локализацией на поверхности почвы опада тростника обыкновенного. Напротив, во всех горизонтах темногумусовой зоотурбированной почвы в карте, заброшенной 45 лет назад, обнаружены плесневые грибы. Кроме того, здесь наблюдается высокая численность актиномицетов, которые являются деструкторами органических веществ, поступивших в почву со сточными водами и дефекатом еще в период использования карты.
Для сравнения, судя по данным, в окружающих ПФ агрочерноземах дисперсно-карбонатных численность, биомасса и число таксонов беспозвоночных значительно меньше, что связано с неблагоприятными физическими свойствами пахотных почв (глы-бистость, высокая плотность, сегрегация железа). Имеющиеся публикации по составу макрофауны в осваиваемых по традиционной технологии черноземах и почвах других типов подтверждают данную закономерность (Tsiafouli et al., 2015; Булгакова, 2017).
Список литературы Сообщества беспозвоночных и микроорганизмов в почвах полей фильтрации Льговского сахарного завода (Курская область)
- Афанасьева Е.А. Черноземы Средне-Русской возвышенности. М.: Наука, 1966. 227 с.
- Андрусевич Е.В., Жуков А.В., Кунах О.Н. Экоморфическая организация сообществ мезопедобионтов как основа зоологической диагностики антропогенных почв // Вісник Харківського національного університету імені В.Н. Каразіна. № 1126. Серія: біологія. Вип. 22. 2014. C. 86-97.
- Аринушкина Е.В. Руководство по химическому анализу почв. М.: Издво Моск. ун-та, 1970. 487 с.
- Базыкина Г.С., Овечкин С.В. Миграционно-мицелярные черноземы Курской области в климатических и биосферных циклах // Бюллетень Почвенного института имени В.В. Докучаева. 2012. Вып. 70. С. 3-17. https://doi.org/10.19047/0136-1694-2012-70-3-18.
- Балабко П.Н., Хуснетдинова Т.И., Карпова Д.В., Славянский А.А. Влияние отходов свеклосахарного производства на плодородие дерновоподзолистой почвы при выращивании картофеля // Агрохимический вестник. 2014. № 6. С. 22-25.
- Баранцевич Е.П., Баранцевич Н.Е. Применение MALDI-TOF массспектрометрии в клинической микробиологии // Трансляционная медицина. 2014. № 6. С. 23-28.
- Белобров В.П., Юдин С.А., Ермолаев Н.Р., Айдиев А.Я. Структура почвенного покрова и трансформация морфометрических параметров типичных черноземов в технологии прямого посева // Бюллетень Почвенного института имени В.В. Докучаева. 2023. Вып. 115. С. 5-31. https://doi.org/10.19047/0136-1694-2023-115-5-31.
- Булгакова М.А. Состав и численность дождевых червей как косвенный показатель физических свойств почв степных агроценозов Предуралья // Современные проблемы науки и образования. 2017. № 2. С. 227.
- Булышева А.М., Хохлова О.С., Бакунович Н.О., Русаков А.В., Мякшина Т.Н. Изменение свойств почв залежного ряда Курской области и тренды восстановления постагрогенных почв лесостепной и степной зон // Почвоведение. 2021. № 8. C. 983-998.
- Вадюнина А.Ф., Корчагина З.А. Методы исследования физических свойств почв. М.: Агропромиздат, 1986. 416 с.
- Возняковская Ю.М., Курдюков Ю.Ф., Попова Ж.П., Лощинина Л.П. Оценка биологического состояния чернозема под различными севооборотами // Почвоведение. 1996. № 9. C. 1107-1111.
- Воробьева Л.А. Химический анализ почв. М.: Изд-во Моск. ун-та, 1998. 272 с.
- Всеволодова-Перель Т.С. Дождевые черви фауны России. Кадастр и определитель. М.: Наука, 1997. 102 с.
- Гиляров М.С., Чернов Ю.И. Почвенные беспозвоночные в составе сообществ умеренного пояса // Ресурсы биосферы. Л.: Наука. 1975. Т. 1. С. 218-240.
- Гиляров М.С., Криволуцкий Д.А. Жизнь в почве. М.: Мол. гвардия, 1985. 191 с.
- Гурин А.Г., Гнеушева В.В. Изменение агрохимических свойств почвы и ее биологической активности при использовании отходов сахарного производства на посевах яровой пшеницы // Вестник аграрной науки. 2018. № 1(70). С. 3-7.
- Железова А.Д., Тхакахова А.К., Ярославцева Н.В., Гарбуз С.А., Лазарев В.И., Когут Б.М., Кутовая О.В., Холодов В.А. Микробиологические показатели агрегатов типичных черноземов в многолетних полевых опытах // Почвоведение. 2017. № 6. С. 711-717.
- Замотаев И.В., Белобров В.П. Эколого-генетические проблемы землепользования на Среднерусской возвышенности (на примере Льговского района Курской области) // Идеи В.В. Докучаева и современные проблемы сельской местности. Ч. 1. Москва-Смоленск, 2001. С. 18-24.
- Замотаев И.В., Белобров В.П., Курбатова А.Н., Белоброва Д.В. Агрогенная и постагрогенная трансформация почв Льговского района Курской области // Бюллетень Почвенного института имени В.В. Докучаева. 2016. Вып. 85. С. 97-114. https://doi.org/10.19047/0136-1694-2016-85-97-114.
- Замотаев И.В., Белобров В.П., Юдин С.А., Белоброва Д.В. Оценка почвенных свойств при мониторинге земель на регионально-локальном уровне // Научные ведомости БелГУ. Серия Естественные науки. 2018. Т. 42. № 2. С. 172-180.
- Замотаев И.В., Грачева Р.Г., Конопляникова Ю.В., Долгих А.В., Карелин Д.В., Белоновская Е.А., Добрянский А.С., Михеев П.В. Почвообразование на отходах сахарной промышленности в Центральном Черноземье // Почвоведение. 2023. № 11. C. 1450-1471.
- Замотаев И.В., Грачева Р.Г., Михеев П.В., Конопляникова Ю.В. Формирование и трансформация почв в районах размещения отходов сахарной индустрии (обзор) // Почвоведение. 2022. № 8. C. 949-961.
- Караваева Н.А., Денисенко Е.А. Постагрогенные миграционномицелярные черноземы разновозрастных залежей южной лесостепи ЕТР // Почвоведение. 2009. № 10. С. 1165-1176.
- Злотин Р.И., Ходашова К.С. Роль животных в биологическом круговороте лесостепных экосистем. М.: Наука, 1974. 220 с.
- Иванов А.Н. Влияние дефеката на свойства, плодородие серых лесных почв Бие-Чумышского междуречья и урожайность сельскохозяйственных культур. Автореф. дис. … к. с./х. н. Барнаул, 2004. 18 с.
- Карелин Д.В., Суховеева О.Э., Глаголев М.В., Добрянский А.С., Сабреков А.Ф., Замотаев И.В. Годовой бюджет углеродсодержащих биогенных парниковых газов при смешанном землепользовании: Льговский район как модельный объект Центрального Черноземья // Почвоведение. 2023. № 8. С. 911-924.
- Классификация и диагностика почв России. Смоленск: Ойкумена, 2004. 342 с.
- Козеев К.Ш., Стрелкова В.И., Тищенко С.А. Влияние переувлажнения на биоту и свойства почв Юга России. Ростов-на-Дону: Ростиздат, 2006. 143 с.
- Коноплина Е.А. Оценка воздействия вторичных ресурсов сахарного производства на биоресурсы агроэкосистем. Автореф. дис. … к. с./х. наук. Воронеж, 2011. 20 с.
- Кутовая О.В., Никитин Д.А., Гераськина А.П. Технология no-till как фактор активности почвенных беспозвоночных в агрочерноземах Ставропольского края // Сельскохозяйственная биология. 2021. Т. 56. № 1. С. 199-210.
- Кутовая О.В., Тхакахова А.К., Чевердин Ю.И. Влияние поверхностного переувлажнения на биологические свойства луговочерноземных почв Каменной степи // Бюллетень Почвенного института имени В.В. Докучаева. 2016. Вып. 82. С. 56-70. https://doi.org/10.19047/0136-1694-2016-82-56-70.
- Мамонтов В.Г., Артемьева З.С., Лазарев В.И., Родионова Л.П., Крылов В.А., Ахмедзянова Р.Р. Сравнительная характеристика свойств целинного, пахотного и залежного чернозема типичного Курской области // Бюллетень Почвенного института имени В.В. Докучаева. 2020. Вып. 101. С. 182-201. https://doi.org/10.19047/0136-1694-2020-101-182-201.
- Методы почвенно-зоологических исследований / Отв. ред. акад. М.С. Гиляров. М.: Наука, 1975. 279 с.
- Микроорганизмы и охрана почв. М.: Изд-во МГУ, 1989. 206 с.
- Минникова Т.В., Русева А.С., Колесников С.И., Труфанов Д.А. Численность бактерий в нефтезагрязненном черноземе обыкновенном после ремедиации // Вестник Московского Университета. Серия 17. Почвоведение. 2023. Т. 78. № 3. С. 65-75.
- Мониторинг природных экосистем Центрально-Черноземного заповедника // Труды Центрально-Черноземного государственного заповедника. 2021. Вып. 20. 411 с.
- Мордкович В.Г. Беспозвоночные животные и диагностика элементарных почвенных процессов // Почвоведение. 1991. № 10. С. 92- 99.
- Мордкович В.Г. Зоологическая диагностика почв: императивы, предназначение и место в составе почвенной зоологии и почвоведения // Журнал общей биологии. 2013. Т. 74. № 6. С. 463-471.
- Мордкович В.Г., Любечанский И.И. Почвенные беспозвоночные русского лесостепья (обзор публикаций XX-XXI вв. и ревизия результатов) // Почвы и окружающая среда. 2020. № 4. С. 1-35.
- Никитин Д.А., Иванова Е.А., Железова А.Д., Семенов М.В., Гаджиумаров Р.Г., Тхакахова А.К., Чернов Т.И., Ксенофонтова Н.А., Кутовая О.В. Оценка влияния технологии no-till и вспашки на микробиом южных агрочерноземов // Почвоведение. 2020. № 12. С. 1508-1520.
- Полевой определитель почв России. М.: Почвенный ин-т им. В.В. Докучаева, 2008. 182 с.
- Прокофьева Т.В., Герасимова М.И., Безуглова О.С., Бахматова К.А., Гольева А.А., Горбов С.Н., Жарикова Е.А., Матинян Н.Н., Наквасина Е.Н., Сивцева Н.Е. Введение почв и почвоподобных образований городских территорий в классификацию почв России // Почвоведение. 2014. № 10. C. 1155-1164.
- Пузанова Л.Н. Агроэкологическая оценка и сельскохозяйственное использование субстрата очистных сооружений свеклосахарных заводов: на примере ОАО “Сахарный комбинат “Льговский””: Автореф. дис. … к. с-х. наук. Курск, 2009. 19 с.
- Пухова Н.Ю., Верховцева Н.В., Ларина Г.Е. Структура микробного сообщества чернозема выщелоченного в зависимости от антропогенной нагрузки // Проблемы агрохимии и экологии. 2011. № 4. С. 42-47.
- Рахлеева А.А. Роль зоогенных структур крупных почвенных беспозвоночных в создании и поддержании неоднородности почвенных свойств (обзор) // Вестник Московского университета. Серия 17: Почвоведение. 2022. № 3. С. 30-35.
- Снегин Э.А., Артемчук О.Ю. Оценка степени устойчивости популяций виноградной улитки (Helix pomatia) к действию генотоксичных компонентов городской среды // Системы контроля окружающей среды. Севастополь: ИПТС. 2018. Вып. 13(33). С. 110-114.
- Щеглов Д.И. Черноземы центра Русской равнины и их эволюция под влиянием естественных и антропогенных факторов. М.: Наука, 1999. 211 с.
- Щеглов Д.И., Чаплыгин Д.Ю., Говоров В.В. Почвенная мезофауна в черноземах Каменной степи // Вестник ВГУ. Серия: Химия. Биология. Фармация. 2006. № 2. С. 184-189.
- Coggan N.V., Hayward M.W., Gibb H. A global database and “state of the field” review of research into ecosystem engineering by land animals // J. Anim. Ecol. 2018. Vol. 87. P. 974-994. https://doi.org/10.1111/1365-2656.12819.
- Giebel R., Worden C., Rust S.M. et al. Microbial fingerprinting using matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS) applications and challenges // Adv. Appl. Microbiol. 2010. Vol. 71. P. 149-184.
- Holland R.D., Wilkes H.G., Rafii F. et al. Rapid identification of intact whole bacteria based on spectral patterns using matrix-assisted laser desorption/ionization with time-of-flight mass spectrometry // Rapid Commun. Mass Spectrom. 1996. Vol. 10. P. 1227-1232.
- IUSS Working Group WRB 2015 World Reference Base for Soil Resources 2014, update 2015 International soil classification system for naming soils and creating legends for soil maps World Soil Resources Reports 106 (Rome: FAO). 192 p.
- Lavelle P., Spain A., Blouin M., Brown G, Decaëns T., Grimaldi M., Jiménez J.J., McKey D., Mathieu J., Velasquez E., Zangerlé A. Ecosystem Engineers in a Self-organized Soil: A Review of Concepts and Future Research Questions // Soil Sci. 2016. Vol. 181. Iss. 3/4. P. 91-109. https://doi.org/10.1097/SS.0000000000000155.
- Le Bayon R.-C., Bullinger-Weber G., Schomburg A., Turberg P., Schlaepfer R., Guenat C. Earthworms as Ecosystem Engineers: A Review. In Earthworms: Types, Roles and Research. New York: NOVA Science Publishers, 2017. P. 129-178. URL: https://www.dora.lib4ri.ch/wsl/islandora/object/wsl:27067.
- Mikheev P., Zamotaev I., Telnova N. Microbial Features in Newly Formed Soils of Disposal Fields from Sugar Refineries // E3S Web of Conferences. Actual Problems of Ecology and Environmental Management (APEEM) 265.03009. 2021. Vol. 265. P. 1-5.
- Tsiafouli M.A., Thébault E., Sgardelis S.P., De Ruiter P.C., Van Der Putten, W.H., Birkhofer K., Hemerik L., de Vries F.T., Bardgett R.D., Brady M.V., Bjornlund L., Jørgensen H.B., Christensen S., Hertefeldt T.D., Hotes S., Gera Hol W.H., Frouz J., Liiri M., Mortimer S.R., Setälä H., Tzanopoulos J., Uteseny K., Pižl V., Stary J., Wolters V., Hedlund K. Intensive agriculture reduces soil biodiversity across Europe. Global Change Biology. 2015. Vol. 21(2). P. 973-985. https://doi.org/10.1111/gcb.12752.
