Сообщества мидий на литорали Кандалакшского залива Белого моря

Автор: Шкляревич Галина Андреевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 11 (105), 2009 года.
Бесплатный доступ
Двустворчатый моллюск мидия, сообщество, индекс разнообразия, литораль
Короткий адрес: https://sciup.org/14749522
IDR: 14749522
Текст статьи Сообщества мидий на литорали Кандалакшского залива Белого моря
Исследования Mytilus edulis L. и сообществ, образуемых этим моллюском, cоставляют в настоящее время одно из важных и приоритетных направлений в изучении естественных процессов, происходящих в экосистемах беломорских мелководий.
Мидия, являясь массовым двустворчатым моллюском-фильтратором, играет одну из самых значительных ролей в экологии Белого моря [3], [9], [1], поэтому существует огромное количество публикаций, посвященных биологии и экологии этого моллюска. Однако исследованиям литоральных поселений мидий посвящено относительно небольшое количество работ [5], [7], [8]. Межгодовые колебания количественных показателей Mytilus edulis также все еще остаются слабо исследованным разделом ее экологии . Понимание причин межгодовой динамики плотности и биомассы мидии в литоральных ее поселениях невозможно без исследования закономерностей феномена видового разнообразия сообществ, образующихся вокруг этого вида-эдификатора.
Цель нашей работы – выполнить описание сообществ литоральной мидии, проследить из
менчивость количественных показателей эди-фикатора и членов образуемых им сообществ в пространстве и времени.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
Работа выполнена в мелководной части акватории, осушаемой во время отливов, – на литорали Кандалакшского залива Белого моря, входящей в состав Кандалакшского государственного природного заповедника (рис. 1).
Исследования проводились по общепринятой в различных модификациях методике количественных гидробиологических работ [4], [12].
Жирной линией выделена материковая и островная территория, серым цветом – акватория Кандалакшского государственного природного заповедника.
Места для мониторинговых межгодовых исследований представляют собой плоские участки песчано-каменистого грунта разной степени за-иленности с более или менее равномерными плотными локальными поселениями Mytilus edulis
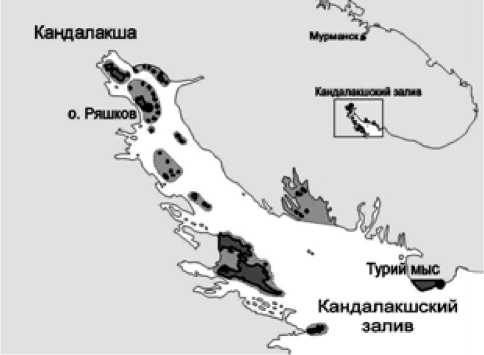
Рис. 1. Картосхема района исследований размером 100–400 м2 в нижнем горизонте литорали. Пробы собирались ежегодно в первой половине июля. Размер пробной площади составлял 0,1 м2. На каждом участке ежегодно бралось по 5 проб, место их сбора определялось случайным образом. Грунт с пробных площадей выбирался до глубины 15–20 см с целью наиболее полного учета всех, в том числе инфаунных организмов. С поверхности камней все животные осторожно и тщательно срезались скальпелем. В полевых условиях проводилась первичная обработка проб: грунтовая часть проб промывалась морской водой через сито с ячеей 1мм2 , все животные разбирались по видам, подсчитывались и после обсушивания на фильтровальной бумаге взвешивались. Животные с массой до 500 мг взвешивались на торсионных весах с точностью до 0,001 г, остальные беспозвоночные – на аптекарских весах с точностью до 0,01 г. Моллюски и усоногие раки-балянусы взвешивались вместе с раковиной и домиком. Большинство беспозвоночных были определены до вида, трудно идентифицируемые животные – до более высоких таксономических рангов.
Местообитания в обеих точках исследования (рис. 1) довольно значительно отличаются друг от друга по степени прибойности, гидродинамики во время приливо-отливных процессов, а также некоторыми зависящими от этого гидрохимическими параметрами водных масс и состава грунта.
Участок 1 расположен на литорали юговосточного мыса о. Ряшкова, который входит в состав Северного архипелага островов. Этот район относительно открытый и поэтому он подвержен довольно значительному гидродинамическому воздействию вод, так как расположен в отдалении от отдельных групп островов и с двух сторон от него находятся Восточная и Западная Ряшковские Салмы, ширина которых колеблется от 1 до более 2 км с глубинами до 25 м.
Участок 2 расположен на заповедной литорали материковой части Турьего мыса – самого глубоководного района, поблизости от побережья этого мыса находится максимальная глубина Бе- лого моря – 340 м. Участок 2 обращен к открытому морю и совсем не прикрыт ни островами, ни мелями, поэтому здесь наблюдается самая высокая степень гидродинамической активности.
В осенне-зимне-весенний период во время становления и разрушения ледового покрова исследуемые биотопы подвергаются его механическому воздействию (частично разрушающему путем прореживания мидиевые поселения и их сообщества) по возрастающей в следующем порядке местообитаний мидий: Ряшков, Турий мыс и в обратном порядке по степени распреснения при весенних процессах ледотаяния и материкового стока.
РЕЗУЛЬТАТЫ
Исследования показали, что сообщества мидий, обитающих на о. Ряшкове, состоят из 14, а на Турьем мысу – из 12 видов и групп беспозвоночных (табл. 1 и 2).
Таблица 1
Средние численность (N, экз./м2) и биомасса (B, г/м2) организмов макрозообентоса, составляющих сообщество Mytilus edulis на юго-восточном мысу о. Ряшкова
|
Вид |
Год |
|||||||
|
2005 |
2006 |
2007 |
2008 |
|||||
|
N |
B |
N |
B |
N |
B |
N |
B |
|
|
Turbellaria g. sp. |
20 |
0,2 |
0 |
0 |
0 |
0 |
0 |
0 |
|
Nemertini |
0 |
0 |
0 |
0 |
20 |
1 |
0 |
0 |
|
Lineus sp. |
0 |
0 |
40 |
0,9 |
0 |
0 |
50 |
2,4 |
|
Nematodes g. sp. |
320 |
0,3 |
0 |
0 |
20 |
0,1 |
475 |
0,5 |
|
Oligochaeta g. |
||||||||
|
sp. |
3580 |
19,1 |
1480 |
3,4 |
4000 |
15,2 |
1100 |
8,6 |
|
Semibalanus balanoides |
0 |
0 |
0 |
0 |
0 |
0 |
800 |
66,2 |
|
Yera albifrons |
120 |
0,3 |
0 |
0 |
0 |
0 |
175 |
2,7 |
|
Gammarus sp. |
1280 |
19,8 |
40 |
2,5 |
920 |
44,5 |
250 |
42,6 |
|
Littorina littorea |
0 |
0 |
0 |
0 |
0 |
0 |
25 |
5,2 |
|
Littorina obtusata |
20 |
2,7 |
300 |
50,6 |
400 |
44,9 |
1150 |
66,6 |
|
Littorina saxatilis |
760 |
86,3 |
20 |
2,4 |
700 |
61,8 |
525 |
66,4 |
|
Hydrobia ulvae |
140 |
1,3 |
1400 |
4,3 |
1440 |
12,1 |
0 |
0 |
|
Mytilus edulis |
13060 |
9108 |
12980 |
3528 |
18980 |
5530 |
16650 |
6163 |
|
Maccoma baltica |
860 |
101,4 |
100 |
37,6 |
220 |
31,3 |
100 |
7,7 |
Таблица 2
Средние численность (N, экз./м2) и биомасса (B, г/м2) организмов макрозообентоса, составляющих сообщество Mytilus edulis на Турьем мысу
|
Вид |
Год |
|||||||
|
2005 |
2006 |
2007 |
2008 |
|||||
|
N |
B |
N |
B |
N |
B |
N |
B |
|
|
Turbellaria |
0 |
0 |
0 |
0 |
0 |
0 |
120 |
0,4 |
|
g. sp. Lineus sp. |
0 |
0 |
0 |
0 |
40 |
0,5 |
20 |
0,1 |
|
Nematodes g. |
420 |
0,3 |
340 |
0,2 |
60 |
0,4 |
700 |
0,6 |
|
sp. Polydora |
0 |
0 |
200 |
0,8 |
0 |
0 |
820 |
2,5 |
|
quadrilobata Fabricia |
0 |
0 |
0 |
0 |
0 |
0 |
80 |
0,7 |
|
sabella Oligochaeta |
5720 |
19,7 |
1460 |
6,7 |
1240 |
6,3 |
3280 |
12,5 |
|
g. sp. Gammarus |
0 |
0 |
0 |
0 |
0 |
0 |
20 |
3,6 |
|
sp. Littorina |
0 |
0 |
140 |
8,3 |
180 |
3,6 |
760 |
51,5 |
|
obtusata Littorina saxatilis |
900 |
18,5 |
1540 |
34,9 |
900 |
23,7 |
500 |
20,7 |
|
Hydrobia |
0 |
0 |
0 |
0 |
0 |
0 |
60 |
0,2 |
|
ulvae Mytilus edulis |
18320 |
3075 |
13580 |
2780 |
21480 |
3619 |
32400 |
4828 |
|
Macoma |
0 |
0 |
20 |
1,8 |
0 |
0 |
20 |
0,1 |
|
baltica |
||||||||
Как видно из табл. 1 и 2, виды и группы макрозообентоса, входящего в состав исследованных сообществ, очень сильно различаются по своей значимости. Mytilus edulis является руководящим или доминантным. Его количественные показатели на порядок и более превосходят плотность и биомассу всех других членов сообществ. Субдоминантными видами сообщества Mytilus edulis на о. Ряшкове являются: Oligochaeta g. sp.; Gammarus sp. ; Littorina obtusata ; Littorina saxatilis; Hydrobia ulvae и Macoma baltica , на Турьем мысу – Nematodes g. sp.; Oligochaeta g. sp.; Littorina saxatilis. К числу второстепенных видов сообществ мидий на о. Ряшкове относятся: Lineus sp .; Nematodes g. sp.; Yera albifrons и Hydrobia ulvae , на Турьем мысу – Lineus sp. ; Polydora quadrilobata; Littorina obtusata , Macoma baltica. Остальные виды являются случайными. Приведенная иерархия видов и групп традиционна [11].
Значение отдельных видов в исследуемых сообществах кроме структурной иерархической значимости должно определяться их ролью в функционировании экосистемы или в ее главнейшей продукционной составляющей. Но при исследовании морских мелководных сообществ установить конкретную функциональную роль видов и отдельных трудно определяемых групп беспозвоночных нелегко, если об их значении судить только по численности и биомассе. Для этого необходимо выполнить анализ биоразно- образия, а также равномерности распределения биомассы и количества видов (J) в сообществе. Для этих целей подходит информационный индекс разнообразия (H) К. Шеннона [10].
Таблица 3
Информационный индекс разнообразия (H), индекс равномерности распределения биомассы (J) и количество видов (N) в исследованных сообществах
Mytilus edulis
|
Местообитание |
H |
J |
N |
|
О. Ряшков, юго-восточный мыс |
0,3 |
0,08 |
14 |
|
Турий мыс |
0,53 |
0,15 |
12 |
Таким образом, индекс разнообразия сообществ, обитающих в различных географических точках, расположенных примерно в 120 км друг от друга, неодинаков, он колеблется от 0,3 до 0,53 бит.
Сравним наши данные с литературными (усредненными по Кандалакшскому и Онежскому заливам), полученными А. И. Бабковым и А. Н. Голиковым [2] для сообществ Mytilus edulis , обитающих на глубине от 1 до 40 м. Число видов в этих сублиторальных сообществах (зообентоса + мейобентоса) составляет в среднем 30 ± 10 при относительно невысоком индексе видового разнообразия (0,9–1,9 бит).
Теперь посмотрим, как изменяются основные биономические показатели вида-эдификатора и всех остальных членов сообщества во времени (рис. 2). Как показано на рис. 2, характеристики средней биомассы Mytilus edulis и суммарной биомассы всех остальных беспозвоночных – членов сообществ довольно значительно варьируют во все годы наших исследований.
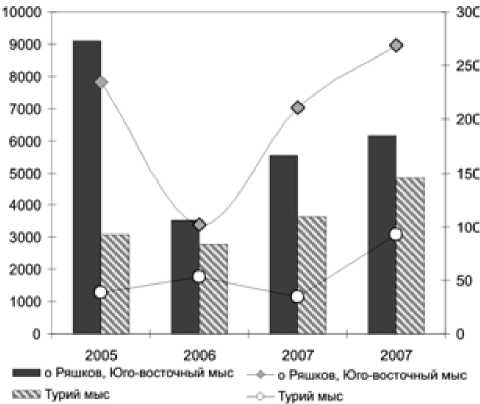
Рис. 2. Межгодовые колебания средней биомассы (г/м2) Myti-lus edulis (гистограмма) и суммарной биомассы всех остальных беспозвоночных – членов сообществ на исследованных участках (кружки и квадраты)
Большую роль в формировании межгодовых и хорологических колебаний количественных показателей мидий играют физические факторы ледового покрова (имеющие разреживающее значение для мидиевых банок). В исследуемых нами поселениях ледовый покров вырывает вмерзшие в нижнюю часть льда, ложащегося на литораль во время отливов, отдельные фрагменты сообществ Mytilus edulis и при подвижках в осенне-весенний период уносит их за пределы сообществ. При этом образуются пятна – окна «чистого» грунта (самого разного размера и формы), освободившегося от обрастания мидиями. Через эти окна происходит вымывание накопившихся илов с токсичным H 2 S, что является положительным процессом восстановления оптимальных условий биотопа мидиевого биоценоза [6]. По краям таких окон во второй половине июля мы ежегодно наблюдали агрегированные скопления спата в увеличивающемся количестве по возрастающей прореживающего действия фактора ледового покрова в следующем порядке местообитаний мидий: о. Ряшков, Турий мыс.
Таким образом, межгодовая изменчивость биомассы мидий в их плотных поселениях зависит от ряда абиотических факторов окружающей среды, а также от биотических внутри- и межвидовых отношений. Из абиотических факторов самое существенное влияние на биомассу Myti-lus edulis оказывает различная ледовая ситуация в осенне-зимне-весенние периоды и колеблющаяся термо-галинная составляющая гидрологического режима в разные годы и в разных участках Кандалакшского залива Белого моря.
В обоих исследованных местообитаниях на биомассу Mytilus edulis практически в одинаковой степени действует фактор хищничества морских звезд Asterias rubens. В одном из них – на о. Ряшкове – мидий кроме звезд активно и в больших количествах поедают различные многочисленные морские птицы [3], [9] и рыбы [1]. На Турьем мысу пресс хищников-птиц и рыб значительно слабее и мидиевые поселения разреживаются в большей степени льдом. В обоих участках восстановление элиминированной части поселений мидий и их сообществ происходит за счет регулярно оседающей из планктона молоди различных беспозвоночных.
Все исследуемые участки мидиевых поселений находятся в охраняемой акватории заповедника, где множество самых различных видов (от беспозвоночных до рыб, птиц и крупных млекопитающих) многочисленных животных образуют гильдию использующих плотные мидиевые поселения в качестве трофического компонента экологической ниши.
Хищническая деятельность всех членов «мидиевой» гильдии и прореживающее воздействие ледового покрова в течение 4 лет оказывали положительное омолаживающее влияние на исследованные мидиевые поселения и их сооб- щества. При отсутствии аналогичного влияния таких факторов естественные сукцессионные процессы приводят к старению мидий и, в конечном счете, к полной гибели этих сообществ.
Mytilus edulis – эдификатор, то есть вид с сильно выраженной средообразующей способностью. Можно сказать, что он кондиционирует условия непосредственно окружающей сообщество среды. Межгодовые колебания средней биомассы вида-эдификатора Mytilus edulis , естественно, сильно влияют на соответствующие временные флюктуации всех членов сообществ. Рис. 2 иллюстрирует довольно четко выраженную прямую зависимость суммарной биомассы всех беспозвоночных – членов сообществ от средней биомассы Mytilus edulis.
ЗАКЛЮЧЕНИЕ
Mytilus edulis , обитающая на литорали Кандалакшского залива Белого моря, часто образует сплошные плотные поселения. При этом вид является эдификатором с сильно выраженной средообразующей способностью к кондиционированию условий непосредственно окружающей свой биоценоз среды. Исследованные сообщества мидий состоят из 12–14 видов и групп беспозвоночных, индекс разнообразия сообществ, обитающих в различных географических точках, колеблется от 0,3 до 0,53 бит.
Основой выявленных нами межгодовых динамических процессов являлись закономерные изменяющиеся абиотические (климатические) факторы окружающей среды, а также внутри- и межвидовые взаимоотношения мидий и их сообществ на литорали Кандалакшского залива. Все закономерно изменяющиеся факторы среды в пространстве и во времени являются инициаторами развития явления биоразнообразия как в целом сообществе, его иерархической, структурной и функциональной характеристиках, так и в его составляющих – популяциях видов. Отдельные популяции видов (или их части – поселения) должны постоянно и непрерывно адаптироваться в двух направлениях. Во-первых, они должны, балансируя на грани толерантности и резистентности своих экологических возможностей, выживать в меняющейся среде, а также передавать информацию о все более и более успешных адаптациях очередному поколению. Во-вторых, они должны адаптироваться к более оптимальному взаимодействию с другими видами в сообществе. Обе группы адаптаций должны развиваться вместе, так как сообщества являются средой, в которой виды должны выживать и эволюционировать.
Работа выполнена при финансовой поддержке программы «Развитие научного потенциала высшей школы 2009–2010 гг.» (грант 3832).
Список литературы Сообщества мидий на литорали Кандалакшского залива Белого моря
- Азаров В. В. Питание рыб на литорали острова Ряшкова и Лодейного в Белом море//Труды Кандалакшского государственного заповедника. Воронеж: Книжное изд-во, 1963. Вып. 4. С. 35-53.
- Бабков А. И., Голиков А. Н. Гидробиокомплексы Белого моря. Л.: Изд-во Зоологического ин-та АН СССР, 1984. 103 с.
- Бианки В. В., Бойко Н. С., Нинбург Е. А., Шкляревич Г. А. Питание обыкновенной гаги Белого моря//Экология и морфология гаг в СССР. М.: Наука, 1979. С. 126-170.
- Кусакин О. Г. Население литорали//Океанология. Биология океана. Биологическая структура океана. Т. 1. М.: Наука, 1977. С. 174-178.
- Луканин В. В., Лангуев Н. К. Распределение и экология локального поселения мидий (Mytilus edulis L.) на беломорской литорали//Экологические исследования перспективных объектов марикультуры фауны Белого моря. Л.: Изд-во Зоологического ин-та АН СССР, 1982. С. 17-24.
- Луканин В. В., Наумов А. Д., Федяков В. В. Поселения мидий: постоянное непостоянство//Природа. 1990. № 11. С. 56-62.
- Максимович Н. В., Герасимова А. В. Долговременный мониторинг литоральных поселений двустворчатых моллюсков (Molluska, Bivalvia) в губе Чупа (Белое море)//Морские и пресноводные биосистемы севера Карелии. СПб.: Изд-во СПбГУ, 2004. С. 95-120.
- Максимович Н. В., Максимович А. Н., Герасимова А. В. Об организации поселений мидий Mytilus edulis L. в условиях литорали Белого моря//Вестник СПбГУ. Сер. 3. 2003. Вып. 4 (№ 27). С. 44-53.
- Перцов Н. А., Флинт В. Е. Питание гаги Кандалакшского заповедника и роль ее в динамике литоральной фауны//Труды Кандалакшского государственного заповедника. Воронеж: Книжное изд-во, 1963. Вып. 4. С. 7-28.
- Шеннон К. Математическая теория связи//Работы по теории информации и кибернетике. М.: Изд-во иностранной лит-ры, 1963. С. 243-332.
- Шитиков В. К., Розенберг Г. С., Зинченко Т. Д. Количественная гидроэкология: методы, критерии, решения. Кн. 1. М.: Наука, 2005. 281 с.
- Шкляревич Г. А. Межгодовая динамика массовых видов бентоса на литорали островов Кандалакшского залива Белого моря//Биология моря. 1980. № 5. С. 26-32.
