Сообщества планктонных организмов оз. Раифское (Волжско-Камский государственный природный биосферный заповедник). II. Вертикальная гетерогенность пелагического планктоннного сообщества
. II. Вертикальная гетерогенность пелагического планктоннного сообщества")
Автор: Жариков В.В., Горбунов М.Ю., Уманская М.В., Быкова С.В., Тарасова Н.Г., Унковская Е.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Водные экосистемы
Статья в выпуске: 1-1 т.13, 2011 года.
Бесплатный доступ
В статье приведены результаты исследования планктонных сообществ оз. Раифское. Показано, что количественные и качественные показатели бактерио-, зоо-, протозоо- и фитопланктона закономерно изменяются по вертикали. Рассмотрены вопросы взаимодействия отдельных компонентов планктона друг с другом в градиенте факторов среды.
Планктонные сообщества, вертикальная структура, трофические связи
Короткий адрес: https://sciup.org/148199577
IDR: 148199577 | УДК: 574.583+574.38
Plankton communities of the lake Raifskoje (Volga-Kama state biosphere reserve). II. Vertical heterogeneity of pelagic plankton community
The results of examination of plankton communities of the lake in summer of 2006 and 2007 are presented. Characteristics of studied components of plankton - bacteria, microalgae, Protozoa and metazooplankton are found to be highly variable with depth. The interactions of separate planctonic groups and its possible changes in a gradient of environmental factors are discussed.
Текст научной статьи Сообщества планктонных организмов оз. Раифское (Волжско-Камский государственный природный биосферный заповедник). II. Вертикальная гетерогенность пелагического планктоннного сообщества
О зеро Раифское — наибольший водоем Раиф-ского участка Волжско-Камского заповедника. Мониторинг гидрохимического режима и планктонных сообществ озера осуществляется с 1983 г. [11], но специальных исследований вертикального распределения организмов планктона при этом не проводилось. Вместе с тем, большая глубина озера, его стратификация в летний период и гидрохимические данные [12] указывают на возможность существенной вертикальной гетерогенности планктонного сообщества. Цель настоящей работы — исследование количественных и структурных изменений в составе пелагического планктона озера в вертикальном градиенте абиотических условий.
МАТЕРИАЛ И МЕТОДЫ
Общая характеристика озера, данные о его гидрологии и гидрохимии представлены ранее [11, 12]. Данные по фито- и зоопланктону озера приведены в [2, 9, 11]. Наши исследования проводились в июле 2006 г. и в августе 2007 г. Пробы для химического анализа и гидробиологических исследований отбирали в пелагиали в области максимальной глубины батометром Руттнера с дискретностью 0,5-2 м в зависимости от глубины слоя. Гидрохимический и гидробиологический
анализ проб проводили стандартными методами [4, 7, 8]. Размерную структуру бактериопланктона определяли, как описано ранее [10]. Учет численности инфузорий, раковинных амеб и метазойно-го микрозоопланктона проводили на глицериновых препаратах, приготовленных из одних и тех же проб (объемом 300 мл), сконцентрированных до 10 мл и фиксированных сулемой. Хотя для точной оценки видового разнообразия многоклеточных животных такой объем проб, возможно, недостаточен, мы полагаем, что полученные результаты позволяют установить общий характер распределения экологических комплексов, в т.ч. микрозоопланктона. Кроме того, подсчет организмов из одной пробы повышает точность проведенных наблюдений и выявленных закономерностей. Идентификацию раковинных амеб, инфузорий и коловраток проводили, соответственно, по [6, 19 и 5], ракообразных учитывали без определения до рода и вида.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
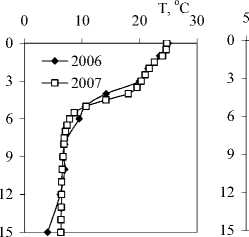
В моменты исследований озеро было стратифицировано, и в нем были сформированы выраженные вертикальные градиенты условий среды (рис. 1). Эпилимнион занимал глубины от поверхности до 3-3,5 м, металимнион — от 3,5 до 5 м, и гиполимнион — ниже 5 м. Температура в пределах эпилимниона снижалась более чем на 4°С, что указывает на его слабое перемешивание.
В 2006 г. в гиполимнионе озера наблюдался еще один, меньший градиент физико-химических характеристик (температура, электропроводность, Eh, концентрация фосфатов) на глубине 10-12 м. Концентрация кислорода начинала падать с глубины 2 или 3 м, и уже на глубине 4 м достигала по данным иодометрического титрования — почти 1 мг/л и аналитического нуля — при измерении кислородным электродом. Окислительно-восстановительный потенциал оставался высоким в 2007 г. во всем столбе воды, а в 2006 г. — до глу- бины 12 м, причем в придонном слое обнаруживались сульфиды в концентрации менее 1 мг/л.
В соответствии с изменяющимися условиями наблюдались закономерные изменения в распределении численности исследованных групп организмов: бактерий, фитопланктона, метазойного
зоопланктона, инфузорий, раковинных амеб, а также хлорофилла " а " (рис. 2), сопровождающиеся структурной перестройкой этих компонентов пелагического планктонного сообщества ("таксоценозов"), вызванной изменением как абиотических, так и биологических факторов.
pH Eh, мВ
9 0 200 400
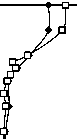
Fe, N-NO2, мг/л
О2, мг/л
P-PO 4 , N-NH 4 , мг/л

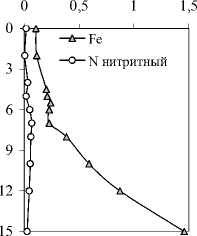
Рис.1. Вертикальное распределение физико-химических параметров и некоторых биогенных элементов в оз. Раифское 12.07.2006 г. и 01.08.2007 г. По оси Y — глубина, м.
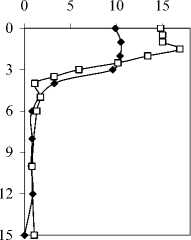
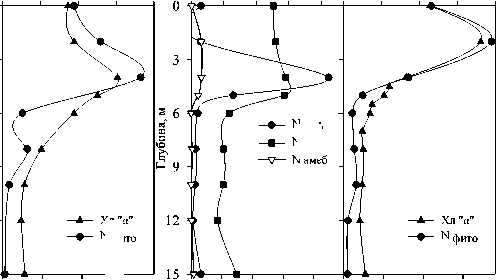
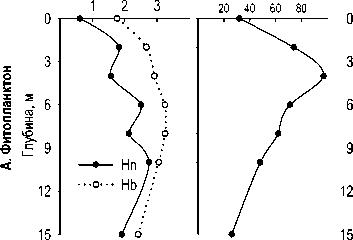
Максимальная концентрация хлорофилла 'а' ', численность и биомасса фитопланктона были зарегистрированы на глубине 4 м в 2006 г. и 2 м в 2007 г. В гиполимнионе численность фитопланктона была низкой (рис. 2). Характерны небольшие пики численности на глубине 8-10 м, обусловленные, видимо, концентрированием оседающего фитопланктона в градиенте плотности.
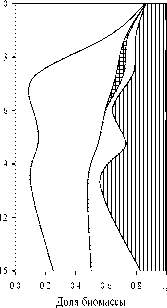
В оба года в фитопланктоне озера по численности доминировали цианобактерии. Их вклад в общую численность на всех горизонтах был более 69%, а на поверхности составлял более 98%. В 2006 г. до глубины 8 м преобладал Microcystis pulverea (Wood) Forti, ниже — Planktolyngbya lim-netica (Lemm.) Komarkova-Legnerova et Cronberg. В 2007 г. M. pulverea развивался, только начиная с глубины 4 м, в слоях 0-2 м доминировали Ana-baena (= Dolichospermium ?) scheremetievii Elenk. и Aphanizomenon flos-aqua (L.) Ralfs, а роль субдоминанта на всех горизонтах перешла от Pl. lim-netica к Pseudanabaena limnetica (Lemm.) Komarek .
По биомассе в эпилимнионе озера доминировали гетероцистные нитчатые цианобактерии с газовыми вакуолями, Aph. flos-aqua и представители р. Anabaena, A. flos-aqua ВгйЬ. ex Bornet et Flauhault (= Dolichospermium flos-aqua (Breb.) Wacklin, Hoffmann et Komarek) в 2006 и A. scheremetievii в 2007 г. В мета- и гиполимнионе вклад цианобактерий в биомассу фитопланктона снижался, особенно сильно в 2006 г., когда на глубине 4-6 м доминировали по биомассе диато мовые р. Aulacozeira (рис. 3). В 2007 г. развитие диатомовых было более слабым, и доминирующей по биомассе группой в мета- и гиполимнионе остались цианобактерии, а второй по биомассе группой — зеленые водоросли.
Вертикальное распределение бактерий в 2006 г. имело два пика численности. Первый из них находился на границе мета- и гиполимниона, ниже максимума фитопланктона, при минимальной численности зоопланктона. Второй максимум, в средней части гиполимниона, совпадал с максимумом численности инфузорий (рис. 2).
Морфологическая и размерная структура бактериопланктона закономерно изменялась по вертикали. В эпилимнионе около половины всех клеток было представлено кокками, 44,5-50,5% общей численности бактерий (ОЧБ), а в мета- и гиполимнионе их вклад снижался до 23,2-37,2% ОЧБ. Поскольку средний объем клеток определяется соотношением числа клеток различных размерных групп, то важно проследить вертикальные изменения размерной структуры бактериопланктона (рис. 4). В эпилимнионе доля клеток, наиболее устойчивых к выеданию простейшими и мик-розооопланктоном (очень мелкие клетки (L < 1 мкм, V < 0,1 мкм3) и очень крупные (L > 5 мкм, V > 1,6 мкм3)), колеблется в пределах 45,6-60,0% ОЧБ, в гиполимнионе, при уменьшении пресса хищников (практически исчезают коловратки, ракообразные и амебы, рис. 2), она составляет только 24,2-35,7% ОЧБ.
В планктоне озера в относительно небольших количествах (1,7-5,7% общей численности бактерий) обнаружены аноксигенные фототрофные бактерии сем. Chromatiaceae (рр. Thiocapsa и Thiopedia ) и Chloroflexaceae (Chloronema giganteum
Dubinina and Gorlenko 1975). Анализ вертикального распределения фототрофных прокариот в целом, включая цианобактерий, демонстрирует выраженное чередование представителей различных порядков (рис. 5).
А.
N инф, 103 экз/л N фито, 106 экз/л
0 5 10 15 20 0 20 40 60 80
Б.
N инф, 103 экз/л N фито, 106 экз/л
3 6 9 12 0 20 40 60 80
Хл " а
N инф
N зоо
N амеб
N амеб
N бакт N
0 1 2 3 0 5 10 15 0,0 0,2 0,4 0,6 0,8 0 5 10 15
Nбакт, 106 кл/мл; Хл " а ", мкг/л Nзоо, Nамеб, 103 экз/л Хл " а ", мкг/л
N3oo, NaMe6, 103 экз/л
Рис.2. Вертикальное распределение общей численности бактерий, фитопланктона, зоопланктона, инфузорий, раковинных амеб и хлорофилла " а " в оз. Раифское 12.07.2006 г. (А.) и 01.08.2007 г. (Б.)
У инфузорий в 2006 г. наблюдалось двухпиковое распределение с максимумом в гиполимнионе, в 2007 г. — однопиковое (рис. 2). При этом и верхний пик в 2006 г., и максимум в 2007 г. регистрировались на глубине 4 м, т.е. в зоне термо- и оксиклина (рис. 1, 2).
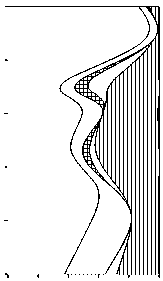
Прочие Зеленые Эвгленовые Динофлагелляты Диатомовые Цианобактерии
0,0 0,2 0,4 0,6 0,8 1,0
Доля биомассы
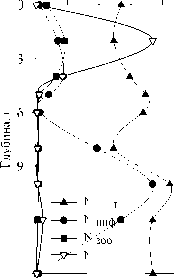
ssp . viridis Ehr. 1831; под оксиклином — Codonella cratera (Leidy, 1877) Imhof, 1885 и скутикоцилиа-тида Cyclidium glaucoma (Muller, 1773) Muller, 1786, а в придонных слоях — не идентифицированные нами до рода мелкие инфузории . Единственным видом в доминирующем комплексе сообщества инфузорий на всех горизонтах был мик-сотроф С. hirtus viridis (вклад по численности колеблется от 12% до 51%).
Размерная структура
Рис 3 . Вертикальное изменение таксономической структуры фитопланктона в оз. Раифское 12.07.2006 г. и 01.08.2007 г.
Морфологическая структура
А



В поверхностных слоях отмечены обычные для пресных водоемов виды из кл. Spirotrichea: Halteria grandinella (Muller, 1773) Dujardin, 1841, Limnostrombidium pelagicum (Kahl, 1932) Krainer, 1995 и представители р. Tintinnopsis, а также сво-бодноживущие (колониальный вид Epistylis pro-cumbens Zacharias, 1897) и эпифитные (представители р. Vorticella, развивающиеся на цианобактериях р. Anabaena и Microcystis ) виды п/кл. Peritrichia класса Oligohymenophorea (табл., рис. 6). В переходной зоне, на границе аэробной и анаэробной зон доминирует простоматида Coleps hirtus
Б
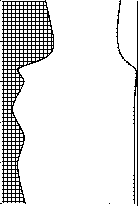
0 20 40 60 80
Доля, %
Нити
Палочки
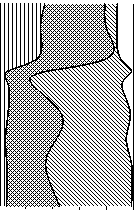
100 0 20 40 60 80
Доля, %
>5 мкм
2-5
1-2

100 0 20 40 60 80 100
Доля, %
>1,6 мкм3
Кокки
<1

0,4-1,6
0,1-0,4
<0,1
Рис. 4. Изменения структуры бактериопланктона оз. Раифское по вертикали в 12.07.2006 г.
Состав комплекса доминирующих видов инфузорий и характер их вертикального распределения в толще воды однотипны в 2006 и 2007 гг. На рис. 6 показана приуроченность максимумов численности отдельных структурообразующих видов к определенным зонам в толще воды.
Численность, млн кл./л
0 20 40 60 80 100 120
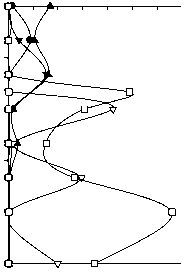
Биомасса, мг/л
0,0 0,5 1,0 1,5 2,0 2,5 3,0
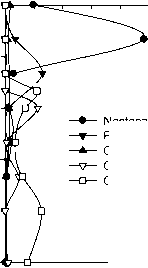
Nostocales
Pseudanabaenales
Chroococcales+Synechococcales
Chromatiales
Chloroflexales
Рис. 5. Вертикальное распределение порядков фототрофных прокариот (цианобактерий и аноксигенных фототрофных бактерий) в оз. Раифское 12.07.2006 г.
В трофической структуре (по данным 2006 г.) сообщества инфузорий планктона во всей толще, за исключением термоклина, оксиклина, доминируют бактериодетритофаги. Лишь на глубине 5-6 м преобладают миксотрофы ( C. hirtus viridis, Spathidium sp.). Наиболее разнообразна структура на поверхности: она представлена бактериодетри-тофагами, альгофагами, и неселективными все-ядами. В гиполимнионе развиваются исключительно бактериофаги.
В 2006 г. в эпилимнионе озера в значительных количествах (до 3 тыс. экз./л) обнаруживались раковинные амебы отр. Arcellinida, в основном Diffugia limnetica (Levander, 1900) Penard, 1902. Максимум их развития располагался на глубине 2 м. В 2007 г. численность арцеллинид снизилась по сравнению с предыдущим годом почти на порядок (рис. 2).
Наибольшее развитие метазойного зоопланктона также наблюдалось в эпилимнионе, причем если в 2006 г. он практически отсутствовал в гиполимнионе, то в 2007 г. его численность оставалась достаточно высокой, более 30% максимальной численности, до самого дна (рис. 2).
Таблица. Виды-доминанты и их вклады (%) по горизонтам в толще воды оз. Раифское 01.08.07
в общую численность сообщества инфузорий
|
Виды |
Горизонт |
||||||||
|
0 м |
2 м |
4 м |
5 м |
6 м |
8 м |
10 м |
12 м |
15 м |
|
|
Vorticella spp. |
15.2 |
15.3 |
0.1 |
1.3 |
0.5 |
6.3 |
2.7 |
1.9 |
0.7 |
|
Limnostrombidium pelagicum |
23.1 |
1.1 |
- |
- |
- |
- |
- |
- |
- |
|
Halteria grandinella |
14.1 |
12.2 |
0.1 |
0.2 |
7.4 |
1.6 |
- |
- |
0,4 |
|
Uronema spp. |
- |
- |
13.5 |
16.6 |
- |
5.5 |
- |
- |
3.0 |
|
Coleps hirtus viridis |
11.6 |
23.3 |
50.8 |
28.7 |
32.4 |
17.3 |
15.5 |
28.3 |
20.1 |
|
Codonella cratera |
1.4 |
7.6 |
0.3 |
2.8 |
3.2 |
22.8 |
14.5 |
18.9 |
9.2 |
|
Cyclidium glaucoma |
- |
- |
45.4 |
60.2 |
41.0 |
18.1 |
30.0 |
11.3 |
7.0 |
|
Мелкий неопределенный вид |
- |
- |
- |
0.3 |
1.1 |
18.9 |
17.3 |
28.3 |
50.9 |
Примечание: виды расположены в порядке смещения максимумов численности от поверхностных горизонтов к придонным; жирным шрифтом выделены вклады по численности > 10%. “- “ вид отсутствует.
2006 2007
Рис. 6. Вертикальное распределение численности доминирующих видов инфузорий в озере Раифское 12.07.06 и 01.08.07.
2006 г.
H n , H b n
n
20 40 60 80
2007 г.
H n , H b 123
n
H n , H b
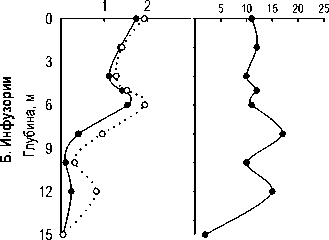
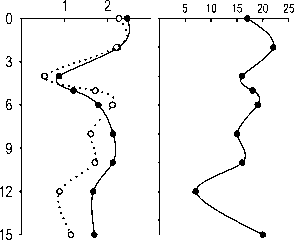
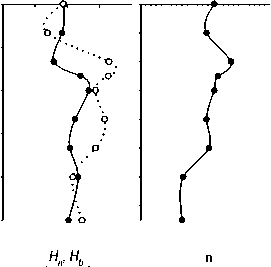
Рис. 7. Вертикальное изменение параметров разнообразия ассоциаций фитопланктона и инфузорий планктона 12.07.06 и 1.08.07: количества видов (и), индекса видового разнообразия Шеннона (по численности - Hn , по биомассе - Нь)
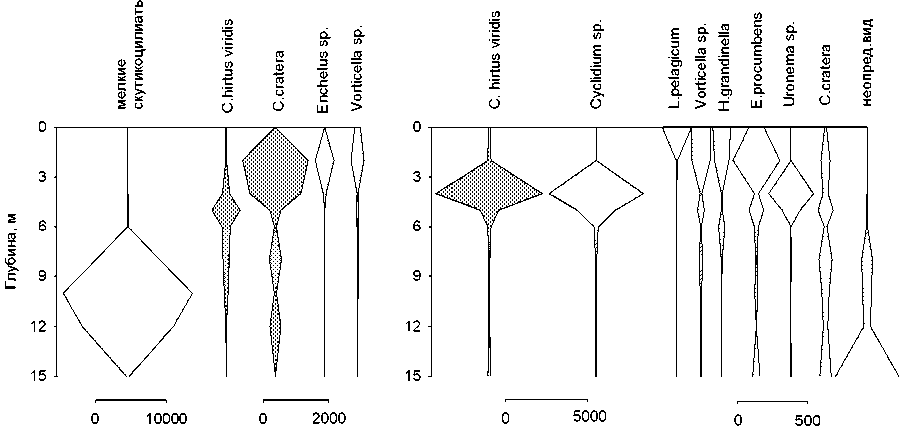
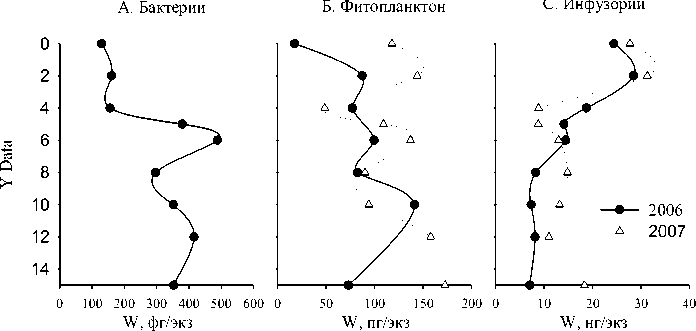
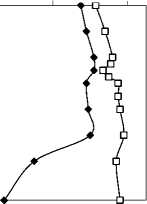
Рис. 8. Вертикальное изменение средней индивидуальной массы клетки бактерий, фитопланктона и инфузорий.
Имеющиеся данные позволяют оценить видовое разнообразие только фитопланктона и инфузорий (рис. 7). Индексы видового разнообразия Шеннона водорослей, рассчитанные по численностям ( Hn ) и биомассам ( Hb ) в оба года достигали максимальных величин в средней части водного столба — в металимнионе или верхней части гиполимниона. В эпилимнионе они были ниже, особенно в 2006 г., из-за сильного доминирования. Максимальное число видов (97) было обнаружено в 2006 г в пробе с глубины 4 м; в поверхностном и особенно придонном слое было зарегистрировано меньшее число видов (рис. 7 А).
Индекс Шеннона инфузорий (по численности) в 2006 г. был довольно стабильным в эпилимнио не, и резко снижался с глубины 8 м (рис.7Б). В 2007 г. наблюдался резкий минимум разнообразия в металимнионе, на глубине 4 м, обусловленный доминированием двух видов — C. hirtus viridis и Cyclidium cf. glaucoma. Число видов инфузорий менялось с глубиной без определенной закономерности (рис. 7Б).
Средняя индивидуальная масса особи инфузорий снижается от поверхности к верхней границе гиполимниона (рис. 8). У водорослей, и особенно у бактерий, наблюдается обратная тенденция. Низкие значения средней массы клетки фитопланктона приурочены к зонам доминирования мелкоклеточных цианобактерий, в первую очередь, M. pulverea. У бактерий наблюдается макси- мум объема клетки на глубине 6 м, видимо, связанный с развитием крупных фото- и хемотроф-ных бактерий в этой зоне.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Вертикальная стратификация водной толщи озер, обусловленная в первую очередь ее термическим расслоением, приводит к формированию многих вторичных градиентов физикохимических условий, в том числе ключевых для развития планктонных организмов [24]. Разнонаправленные градиенты условий приводят к вертикальному расслоению различных компонентов пелагического планктонного сообщества в соответствии с оптимумами условий среды, взаимодействиями с другими биотическими воздействиями (выеданием, синтрофными взаимодействиями и т.д.) и разнообразными компромиссами между ними.
Наиболее явно вертикальное расслоение планктонных сообществ проявляется в озерах с сульфид-содержащим гиполимнионом, и особенно в меромиктических озерах с аномально высокими градиентами окислительно-восстановительного потенциала и биогенных элементов в зоне хемоклина [3, 13, 18, 20, 21, 23, 26 и многие др.]. В таких озерах планктонные организмы, в массовых количествах развивающиеся в узких градиентных зонах, оказывают значительное влияние непосредственно на среду их обитания в отдельных слоях, влияя на световой режим, потребляя различные химические компоненты и выделяя кислород.
Хотя по классификации озер, принятой в Рамочной европейской водной директиве [17], оз. Раифское является малым (<5 км2), оно является одним из наибольших по площади, глубине и объему воды среди озер бассейна Средней и Нижней Волги. Озеро относительно малопродуктивно, с малым содержанием сульфатов в воде, поэтому в нем не может сформироваться сульфид-содержащий гиполимнион с высоковосстановительными условиями [3]. Однако ниже слоя 4 м в воде озера регистрируются только следы кислорода или его полное отсутствие.
Озеро питается двумя притоками, приносящими воду с контрастными химическими характеристиками: р. Сумка несет светлую воду с низким содержанием железа, а р. Сер-Булак — хотя и менее минерализованную, но намного более гумоз-ную (около 500 0Pt) воду с высоким содержанием железа. Смешение этих вод создает сложную гидродинамическую картину перемешивания озера и, как мы предполагаем, ответственно за вторичный градиент физико-химических условий в озере на глубинах 8-12 м. Однако в 2007 г. этот режим был нарушен, так как р. Сер-Булак выше впадения в озеро и р. Сумка ниже истока из озера, были перегорожены бобровыми плотинами. В результате питание озера летом 2007 г. осуществлялось только за счет вод р. Сумка, а его уровень в результате подпруживания увеличился по сравнению с предыдущим годом на ~0.3-0.5 м. Не смотря на это, термический режим озера не претерпел существенных изменений. Эпилимнион с небольшим градиентом температуры охватывал глубины от поверхности до 3-3.5 м, а металимнион — в диапазоне от 3.5 до 5 м.
Тем не менее, с 2006 по 2007 г. в составе планктонного биоценоза, судя по полученным данным (рис. 2, 3, 5 и 6), произошли существенные изменения, которые затрудняют выделение устойчивых стабильных комплексов планктонных видов, приуроченных к различным условиям обитания. Особенно динамичным оказалось сообщество фитопланктона, в котором произошли наиболее существенные перестройки, однако неодинаковым в 2006 и 2007 гг. было и распределение раковинных амеб, инфузорий и зоопланктона.
Гиполимнион в оз. Раифское имел окислительные условия и не содержал сероводорода (за исключением придонных слоев), поэтому в нем не наблюдался слой с доминированием сульфид-толерантных видов фитопланктона и инфузорий. Облигатно анаэробные инфузории ( Metopus sp.) регистрировались только в придонном слое, а крупные бентосные мигранты, характерные для хемоклина озер с сульфидным гиполимнионом [15], вообще не вносили сколь либо значительного вклада в состав инфузорий. Хотя криптофито-вые и эвгленовые водоросли, как и в озерах с сульфидным гиполимнионом [20, 27] формировали максимумы в области термоклина, в оз. Раифское они составляли пренебрежимо малую часть фитопланктона, особенно по численности. Цианобактерии сем. Pseudanabaenaceae, толерантные к сульфидам [16], развивались во всем столбе воды без ярко выраженных максимумов, а представители р. Planktothrix , ряд видов которого также тяготеют к хемоклину озер [21, 22], вообще не были обнаружены. Видимо, в условиях, сложившихся в озере, эти виды не могли конкурировать в области термоклина с видами, развивавшимися в эпилимнионе, а в зоне вторичного хемоклина (8-12 м), при прозрачности 0.8-1.65 м, не могли развиваться из-за отсутствия света. Следует отметить, что развитие перечисленных видов в металимнионе было большим в 2006 г., когда градиенты физико-химических условий были более выражены.
Характерной особенностью зоны металимниона было присутствие максимума фототрофных бактерий на глубине 5 м. В отличие от сульфидсодержащих озер, этот максимум был сформирован нитчатыми аноксигенными фототрофами (Chloronena giganteum) и физиологически -гибкими пурпурными бактериями, способными развиваться в отсутствие сероводорода в окружающей водной массе [14]. За исключением максимума, обусловленного развитием этих групп бактерий и, возможно, видов, восстанавливающих соединения азота и отвечающих за максимум нитритов в этой зоне, общая численность бактерий заметно увеличивалась с глубиной. Еще одной чертой, сближающей металимнион оз. Раиф-ского со стратифицированными сульфид- содержащими озерами, был максимум развития миксотрофной инфузории С. hirtus viridis, который в оба года располагался именно в области металимниона (хемоклина).
Очевидно, развитие характерного металимни-ческого комплекса видов планктона в стратифицированных озерах определяется в первую очередь противоположно направленными градиентами освещенности и окислительновосстановительных условий [21]. В тех случаях, когда второй градиент не выражен (как в исследованном озере), виды, адаптированные к условиям хемоклина, не получают преимуществ в развитии и, за немногими исключениями, проигрывают конкуренцию типичным обитателям эпилимниона или видам с широким экологическим диапазоном.
Важно отметить, что, судя по взаимному положению максимумов биомасс и численностей различных групп организмов, характер их трофических взаимодействий изменяется с глубиной. Так, противофазный характер развития инфузорий и бактериопланктона в эпилимнионе свидетельствует, очевидно, о контроле численности бактерий инфузориями, т.е. о "top-down" контроле. Наоборот, в гиполимнионе численность этих групп организмов меняется синфазно, что указывает на преобладание процессов "bottom-up" контроля, т.е. регуляцию развития инфузорий доступными ресурсами (бактериопланктоном).
ЗАКЛЮЧЕНИЕ
Оз. Раифское по своим гидрологическим и гидрохимическим параметрам относится к группе стратифицированных озер с невысокой трофно-стью и низким содержанием сульфатов в воде. Особенностью таких озер является высокий окислительно-восстановительный потенциал во всей водной толще и отсутствие сильных восстановителей (в первую очередь, сульфидов) до или почти до дна [3]. Однако градиент освещенности, а также формирующиеся в период стратификации градиенты температуры, содержания кислорода и биогенных элементов обусловливают сложный характер вертикального распределения планктонных организмов и их трофических взаимодействий.
Проведенное исследование позволило дать характеристику вертикальной структуры пелагического планктонного сообщества в оз. Раифское в годы с контрастным гидрологическим режимом. Несмотря на сильные межгодовые различия, удалось выделить некоторые общие особенности вертикальной структуры пелагического планктонного сообщества. В отличие от озер с сульфидсодержащим анаэробным гиполимнионом, в оз. Раифское перестройка планктонного сообщества, выражающаяся в смене размерной структуры, видового состава и ряда количественных показателей, охватывает более широкий диапазон глубин и имеет более сложный характер. В трофической структуре планктонного сообщества эпилимниона преобладают процессы "top-down" кон троля, тогда как с увеличением глубины и соответственным изменением условий, их роль, видимо, снижается, и в гиполимнионе основная роль уже принадлежит процессам "bottom-up" контроля. Тем не менее, для более полного выявления закономерностей вертикальной структуры планктонного сообщества в этом и подобных озерах необходим не только анализ исследованных групп организмов планктона, но и изучение других групп пелагических организмов (вплоть до высшего трофического звена — рыб), а также бентосных сообществ и их взаимодействия.
Авторы выражают признательность д.б.н. Ю.А. Мазею за помощь в определении раковинных амеб.
Список литературы Сообщества планктонных организмов оз. Раифское (Волжско-Камский государственный природный биосферный заповедник). II. Вертикальная гетерогенность пелагического планктоннного сообщества
- Горбунов М.Ю. Вертикальное распределение бактериохлорофиллов в гумозных озерах Волжско-Камского заповедника (респ. Татарстан)//Поволжск. экол. журн. 2011. (в печати).
- Деревенская О.Ю., Унковская Е.Н., Мингазова Н.М., Павлова Л.Р. Структура сообществ зоопланктона озер Раифского участка Волжско-Камского заповедника и его охранной зоны//Тр. Волжско-Камского гос. природного заповедника. 2002. Вып. 5. С. 53-70.
- Кузнецов С.И. Микрофлора озер и ее геохимическая деятельность. Л.: Наука, 1970. 440 с.
- Кузнецов С.И., Дубинина Г.А. Методы изучения водных микроорганизмов. М.: Наука, 1979. 288 с.
- Кутикова Л. А. Коловратки фауны СССР. Л.: Наука, 1970. 744 с.
- Мазей Ю.А., Цыганов А.Н. Пресноводные раковинные амебы. Москва: КМК. 2006. 300 с.
- Методика изучения биогеоценозов внутренних водоемов. М., 1975. 240 с.
- Новиков Ю.В., Ласточкина К.О., Болдина З.Н. Методы исследования качества воды водоемов. М.: Медицина, 1990. 400 с.
- Палагушкина О.В., Бариева Ф.Ф., Унковская Е.Н. Видовой состав, биомасса и продуктивность фитопланктона озер Раифского участка Волжско-Камского заповедника и его охранной зоны//Тр. Волжско-Камского гос. природного заповедника. 2002. Вып. 5. С. 37-52
- Уманская М.В. Экологические особенности развития бактериопланктона малых эвтрофных озер Самарской луки. Дисс. … канд.биол.наук. Тольятти, 2004. 148 с.
- Унковская Е.Н., Жариков В.В., Быкова С.В. Горбунов М.Ю., Уманская М.В., Тарасова Н.Г., Мухортова О.В., Палагушкина О.В. Деревенская О.Ю. Сообщества планктонных организмов оз. Раифское (Волжско-камский государственный природный биосферный заповедник). I. Биоразнообразие планктонных сообществ различных биотопов озера Раифское//Изв. Сам.НЦ РАН. 2010. Т.12, № 1(5). С. 1453-1460.
- Унковская Е.Н., Мингазова Н.М., Павлова Л.Р. Гидрологическая и гидрохимическая характеристика водоемов Раифы//Тр. Волжско-Камского гос. природного заповедника. 2002. Вып. 5. С. 9-36.
- Abella C.A., Garcia-Gil L.J. Microbial ecology of planktonic filamentous phototrophic bacteria in holomictic freshwater lakes//Hydrobiologia http://www.springerlink.com/content/100271/?p=8d0322d3032c49c7b542f944f7dee6ee&pi=0>. 1992. P. http://www.springerlink.com/content/r3867065n50v/?p=8d0322d3032c49c7b542f944f7dee6ee&pi=0> 79-86
- Arvola L. Vertical distribution of primary production and phytoplankton in two small lakes with different humus concentration in southern Finland//Holarctic Ecol. 1984. V.7. P. 390-398.
- Bark A.W. Studies on ciliated protozoa in eutrophic lakes: I. Seasonal distribution in relation to thermal stratification and hypolimnetic anoxia//Hydrobiologia. 1985. V.124, No. 2. P.167-176.
- Cohen Y., Jorgensen B.B., Pandan E., Shilo M. Sulphide-dependent anoxygenic photosynthesis in the cyanobacterium Oscillatoria limnetica//Nature. 1975. V.257, No.5526. P. 489-492.
- Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 establishing a framework for Community action in the field of water policy//Official J. EC. 2000. L 327, 22-12-2000. 72 p.
- Finlay B.J., Fenchel T. Ecology: Role of Ciliates in the Natural Environment/Ciliates: Cells as organisms/K. Hausmann and P.C. Bradbury (eds.)//Stuttgart: Gustav Fischer Verlag, 1996. P. 117-140.
- Foissner W., Berger H., Schaumdurg J. Identification and ecology of limnetic plankton ciliates. Informationsberichte des Bayer. Landesamtes für Wasserwirtschaft. 1999. Hf. 3. 793 p.
- Gervais F. Ecology of cryptophytes coexisting near a freshwater chemocline//Freshwater Biol. 1998. V. 39, No. 1. P.61-78.
- Gervais F., Siedel U., Heilmann B., Weithoff G., Heisig-Gunkel G., Nicklisch A. Small-scale vertical distribution of phytoplankton, nutrients and sulfide below the oxycline of a mesotrophic lake//J.Plankton Res. 2003. V.25, No.3. P.273-278.
- Komárek J. Planktic oscillatorialean cyanoprokaryotes (short review according to combined phenotype and molecular aspects)//Hydrobiologia 2003. V. 502, No. 1-3. P. 367-382.
- Miracle M.R., Vicente M., Pedrós-Alió C.Biological studies of Spanish meromictic and stratified karstic lakes//Limnetica. 1992. V. 8. P.59-77.
- Padisak J., Reynolds C. Shallow lakes -the absolute, the relative, the functional and the pragmatic//Hydrobiologia. 2003. V. 506-509. P.1-11.
- Wojciechowska W., Poniewozik M., Pasztaleniec A. Vertical distribution of dominant cyanobacteria species in three lakes -evidence for tolerance to different turbulence and oxygen conditions//Pol. J. Ecol. 2004. V.52, No.3. P.347-351.
- Zingel P., Ott I. Vertical distribution of planktonic ciliates in strongly stratified temperate lakes. Hydrobiologia. 2000. V. 435, No. 1-3. P. 19-26.
- Žvikas A. Structure of microorganism communities and pecularities of their activities in gypsum karst lakes of Northern Kithuania. Summary of doctoral dissertation. Vilnius, 2005. 37 p.
