Сообщества почвенных нематод в условиях интродукции древесных растений на территории Ботанического сада Петрозаводского государственного университета

Автор: Сущук Анна Алексеевна, Калинкина Дарья Сергеевна, Платонова Елена Анатольевна
Журнал: Hortus Botanicus @hortbot
Рубрика: Сохранение, мобилизация и изучение генетических ресурсов растений
Статья в выпуске: 11, 2016 года.
Бесплатный доступ
В статье рассмотрены особенности сообществ нематод в корнеобитаемом слое почвы подкронового пространства интродуцированных древесных растений на территории Ботанического сада Петрозаводского государственного университета (Республика Карелия). В качестве оценочных параметров использованы таксономическое разнообразие, плотность популяций нематод, эколого-трофическая структура сообществ и эколого-популяционные индексы, основанные на анализе фауны нематод. В результате исследования был обнаружен 51 род нематод, среди которых 6 – паразиты растений. Выявлено, что в почве под хвойными культурами доминирующей группой являются бактериотрофы, субдоминантами в большинстве случаев выступают микотрофы; в почве с посадками лиственных пород снижается доля нематод-бактериотрофов и увеличивается доля паразитов растений. Анализ эколого-популяционных индексов сообществ нематод показал, что почвенная трофическая сеть в местах посадок лиственных интродуцентов характеризуется как многокомпонентная и стабильная, а под хвойными интродуцентами – как деградированная (базальная).
Древесные интродуценты, ботанический сад, почвенные нематоды, плотность популяций, структура сообществ, нематоды-паразиты растений
Короткий адрес: https://sciup.org/14748480
IDR: 14748480 | УДК: 631.467.2:630*271
Soil nematode community under the non-native trees in the Botanic Garden of Petrozavodsk State University
The particularities of soil nematode communities of the rhizosphere of non-native trees were studied in the Botanic Garden of Petrozavodsk State University (Republic of Karelia). Taxonomic diversity, abundance, community structure and ecological indices derived from nematode fauna analysis were used as the evaluation parameters. Nematode fauna included 51 genera, 6 of them were plant parasitic. The dominant eco-trophic group in the nematode community structure of coniferous trees was bacterial feeders; fungal feeders in most cases were observed in the second numbers. The contribution of bacterial feeders was decreased and plant parasites were increased in eco-trophic structure of nematode communities of deciduous trees in compared with coniferous trees. Analysis of ecological indices showed that the state of soil nematode communities reflects complex, structured (stable) soil food web in the biocenoses with deciduous trees, and degraded (basal) food web – under coniferous trees.
Текст научной статьи Сообщества почвенных нематод в условиях интродукции древесных растений на территории Ботанического сада Петрозаводского государственного университета
Нематоды (круглые черви) – одна из наиболее многочисленных и разнообразных групп организмов, обитающих в почве. Свободноживущие нематоды в качестве вторичных консументов играют ключевую роль в процессах, происходящих в почве, регулируя бактериальные и грибные популяции, изменяя круговорот питательных веществ, главным образом путем стимуляции роста микробов, участвуют, таким образом, вместе с другими организмами в процессе минерализации веществ и создании почвенного плодородия. Часть видов нематод являются паразитами растений: питаясь за счет живых растений, они влияют на их рост, развитие, продуктивность. Изучение нематод-паразитов растений важно с практической точки зрения: некоторые виды относятся к карантинным объектам, опасным вредителям сельскохозяйственных культур, снижают урожай, являются переносчиками вирусов растений. Кроме того, использование нематод в качестве биологических индикаторов для оценки состояния почвенных экосистем в настоящее время является признанным фактом (Bongers, 1990; Ferris et al., 2001).
Растительное сообщество значительно влияет на педофауну, включая группы нематод, трофически связанные с растениями (Wardle et al., 2003); в этой связи занос чужеродных для региона видов растений может косвенно способствовать изменению численности и разнообразия фитопаразитов, более широкому их расселению. В современной нематологической литературе имеются данные о воздействии планомерной интродукции и выращиваемых новых видов растений на особенности фауны почвенных нематод в целом и группы фитопаразитов, в частности, на примере питомников древесных пород (Skwiercz, 2012; Chalanska, Labanowski, 2014), агроценозов с плодовоягодными культурами (Tabolin et al., 2010; Pokharel et al., 2015), рекультивируемых ландшафтов с посадками лиственных и хвойных деревьев (Hanel, 2008) и условий закрытого грунта в ботанических садах (Gubin, Sigareva, 2014). Однако детальные работы, посвященные изучению сообществ почвенных нематод в сфере влияния интродуцируемых древесных растений, культивируемых в эстетических целях и для сохранения/увеличения биоразнообразия в экосистемах Севера, отсутствуют. Подобные исследования важны для разработки методов защиты растений в ботанических садах, а также учета и сохранения биоразнообразия растительного и животного компонентов в садово-парковых экосистемах урбанизированных территорий.
Цель работы – изучение фауны почвенных нематод, в том числе паразитов растений, корнеобитаемого слоя древесных интродуцированных растений на территории Ботанического сада Петрозаводского государственного университета (ПетрГУ).
Объекты и методы исследований
Отбор почвенных образцов проводился в июле 2013 г. на территории Ботанического сада Петрозаводского государственного университета (61°50'36,94" с.ш., 34°23'19,27" в.д.) в верхнем корнеобитаемом слое почвы в пределах подкронового пространства 10 видов деревьев (рис. 1): сосна Веймутова ( Pinus strobus L.), ель канадская, или сизая ( Picea glauca (Moench) Voss), пихта бальзамическая ( Abies balsamea (L.) Mill.), пихта сибирская ( Abies sibirica Ledeb.), лиственница сибирская ( Larix sibirica Ledeb.), сосна сибирская кедровая ( Pinus sibirica Du Tour), дуб черешчатый ( Quercus robur L.), липа сердцевидная ( Tilia cordata Mill.), вяз гладкий ( Ulmus laevis Pall.), клен остролистный ( Acer platanoides L.). Почвенные образцы отбирались на глубину 10 см в 9 повторностях. Почвы в местах произрастания интродуцентов – супесчаные с большим количеством камней и валунов. Местами встречаются небольшие песчаные наносы и суглинки. Нередки выходы скальных пород (базальтов). По уровню рН солевой вытяжки почва в корнеобитаемом слое древесных растений сильно кислая (рН сол.=3.8–4.8).
Фитоценологические исследования включали описание растительности на пробных площадях (25 кв. м) в подкроновом пространстве исследуемых видов деревьев. Травяно-кустарничковый ярус представлен в основном аборигенными луговыми и лесными травами с небольшим участием бореальных кустарничков, встречается возобновление ряда древесных растений (как местной, так и инорайонной флоры). Видовой состав и проективное покрытие травяно-кустарничкового яруса под разными видами деревьев варьируют в широких пределах (табл. 1).
Нематод выделяли из почвы (навеска 30 г) модифицированным методом Бермана. Фиксацию материала производили при помощи ТАФ (триэтаноламин, формалин и вода в соотношении 2:7:91) (van Bezooijen, 2006). Идентификацию нематод осуществляли на временных глицериновых препаратах. Каждый таксон согласно классификации Йейтса с соавторами относили к одной из шести экологотрофических групп: бактериотрофам (Б), микотрофам (М), политрофам (П), хищникам (Х), нематодам, ассоциированным с растениями (Аср) и паразитам растений (Пр) (Yeates et al., 1993).
Для оценки состояния сообществ нематод использованы следующие параметры: таксономическое разнообразие (количество родов), индекс Шеннона H' , плотность популяций (численность) нематод (кол-во экз./100 г сырой почвы), эколого-трофическая структура. Также были проанализированы экологопопуляционные индексы сообществ нематод: индекс зрелости сообществ (ΣMI) (Bongers, 1990) и индексы, характеризующие почвенную трофическую сеть (индексы структурирования (SI) и обогащения (EI) почвенной трофической сети, индекс преобладающего пути разложения органического вещества в почве (CI)) (Ferris et al., 2001).
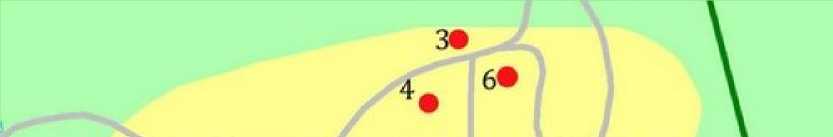
Сектор восточ но азиатской флоры
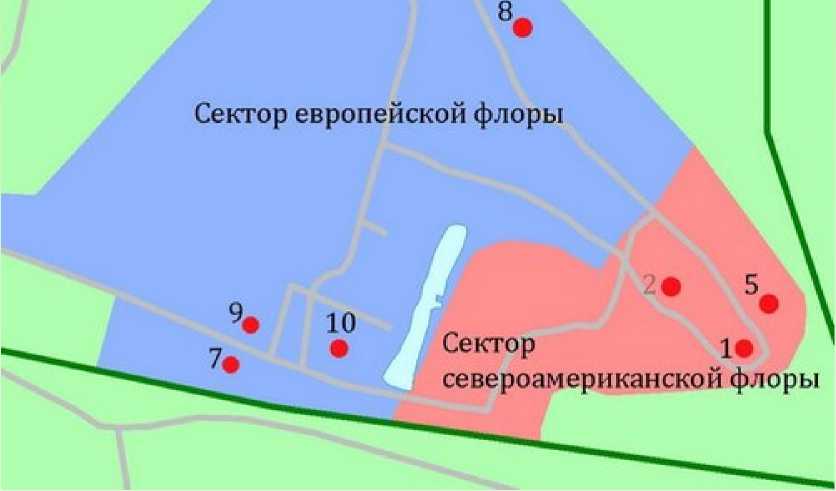
Рис. 1. Расположение пробных площадей на территории арборетума Ботанического сада ПетрГУ.
1 – сосна Веймутова, 2 – ель канадская, 3 – лиственница сибирская, 4 – сосна сибирская кедровая, 5 – пихта бальзамическая, 6 – пихта сибирская, 7 – липа сердцевидная, 8 – клен остролистный, 9 – вяз гладкий, 10 – дуб черешчатый.
Fig. 1. The location of sample plots on the arboretum of the Botanic Garden of PetrSU.
1 – Pinus strobus , 2 – Picea glauca , 3 – Larix sibirica , 4 – Pinus sibirica , 5 – Abies balsamea , 6 – Abies sibirica , 7 – Tilia cordata , 8 – Acer platanoides , 9 – Ulmus laevis , 10 – Quercus robur .
Индекс ΣMI представляет собой полуколичественную оценку состояния экосистемы на основе состава и соотношения таксонов нематод с различными экологическими преференциями, связанными с их морфологией, биологией и экологией, выраженных в значениях, присвоенных каждому таксону по специальной с-р-шкале Бонгерса: от колонизаторов со значением 1, устойчивых к неблагоприятным условиям существования, до персистеров со значением 5, чувствительных к факторам окружающей среды. Индекс обогащения почвенной трофической сети (enrichment index, EI) прямо коррелирует с обогащением почвы органикой или уровнем плодородия, индекс структурирования трофической сети
(structure index, SI) отражает сложность и стабильность почвенной экосистемы. CI (channel index) показывает преобладающий путь разложения органического вещества в почвенной экосистеме – с участием бактерий или почвенных грибов – и рассчитывается на основе численности нематод-бактериотрофов и микотрофов в сообществе. Низкие значения CI свидетельствуют об активном участии бактерий в разложении органики, высокие – о преобладании грибов в данном процессе (Ferris et al., 2001).
Вследствие малого числа повторностей был использован H-критерий Краскела–Уоллиса. Различия между группами считали достоверными при р < 0.05. Числовые данные представлены в форме М ± SE (среднее значение ± стандартная ошибка среднего). Расчеты выполнены при помощи программы PAST 1.68. (Hammer et al., 2001).
Для выполнения исследования использовано оборудование центра коллективного пользования ИБ КарНЦ РАН «Комплексные фундаментальные и прикладные исследования особенностей функционирования живых систем в условиях Севера».
Таблица 1. Состав травяно-кустарничкового яруса под пологом исследуемых древесных растений на пробных площадях 25 м².
Table 1. The composition of the understory vegetation in the 25 m² sample plots under the canopy of investigated woody plants.
|
Параметры |
1* |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Сомкнутость крон |
0.6 |
0.5 |
0.5 |
0.3 |
0.9 |
0.7 |
0.8 |
0.7 |
0.9 |
0.6 |
|
Покрытие травянокустарничкового яруса, % |
30 |
60 |
90 |
85 |
2 |
65 |
20 |
50 |
30 |
35 |
|
Количество видов в травянокустарничковом ярусе |
17 |
21** |
23 |
26 |
10 |
30 |
14 |
23 |
6 |
12** |
|
Травяно-кустарничковый ярус, Проективное покрытие, % |
||||||||||
|
Achillea millefolium L. |
3 |
+ |
+ |
|||||||
|
Aegopodium podagraria L. |
5 |
5 |
5 |
30 |
1 |
|||||
|
Agrostis capillaris L. |
3 |
5 |
7 |
+ |
||||||
|
Alchemilla vulgaris auct. |
3 |
15 |
20 |
+ |
+ |
|||||
|
Angelica sylvestris L. |
+ |
1 |
+ |
1 |
1 |
|||||
|
Anthoxanthum odoratum L. |
3 |
r |
||||||||
|
Anthriscus sylvestris Hoffm. |
3 |
20 |
3 |
3 |
+ |
3 |
+ |
1 |
||
|
Campanula glomerata L. |
+ |
|||||||||
|
Campanula latifolia L. |
+ |
+ |
30 |
+ |
||||||
|
Campanula rotundifolia L. |
1 |
+ |
||||||||
|
Carex digitata L. |
+ |
+ |
||||||||
|
Carex pallescens L. |
+ |
|||||||||
|
Centaurea jacea L. |
+ |
r |
5 |
|||||||
|
Chamaenerion angustifolium (L.) Scop. |
+ |
|||||||||
|
Clinopodium vulgare L. |
1 |
|||||||||
|
Convallaria majalis L. |
+ |
+ |
||||||||
|
Corydalis solida (L.) Clairv. |
+ |
7 |
||||||||
|
Dactylis glomerata L. |
3 |
8 |
5 |
+ |
||||||
|
Deschampsia cespitosa (L.) P. Beauv. |
2 |
+ |
||||||||
|
Deschampsia flexuosa (L.) Trin. |
25 |
10 |
3 |
+ |
||||||
|
Dianthus deltoides L. |
3 |
|||||||||
|
Dryopteris carthusiana (Vill.) H. P. Fuchs |
+ |
|||||||||
|
Epilobium montanum L. |
+ |
|||||||||
|
Festuca ovina L. |
2 |
|||||||||
|
Festuca pratensis Huds. |
5 |
|||||||||
|
Fragaria vesca L. |
1 |
4 |
5 |
10 |
+ |
|||||
|
Galium mollugo L. |
1 |
2 |
15 |
2 |
+ |
+ |
+ |
|||
|
Geranium sylvaticum L. |
7 |
1 |
10 |
15 |
5 |
1 |
||||
|
Geum urbanum L. |
+ |
+ |
+ |
|||||||
|
Glechoma hederacea L. |
3 |
|||||||||
|
Heracleum sibiricum L. |
1 |
|||||||||
|
Hieracium penduliforme (Dahlst.) Johanss. |
+ |
|||||||||
|
Hieracium pratense Tausch |
2 |
|||||||||
|
Hieracium umbellatum L. |
+ |
+ |
||||||||
|
Hieracium vulgatum Fries |
+ |
1 |
||||||||
|
Hypericum maculatum Crantz |
+ |
2 |
8 |
3 |
+ |
+ |
||||
|
Knautia arvensis (L.) Coult. |
+ |
3 |
5 |
1 |
+ |
|||||
|
Lathyrus pratensis L. |
+ |
+ |
+ |
|||||||
|
Leucanthemum vulgare (Vaill.) Lam. |
+ |
5 |
||||||||
|
Luzula pilosa (L.) Willd. |
+ |
+ |
1 |
+ |
||||||
|
Maianthemum bifolium (L.) F. W. Schmidt |
+ |
10 |
||||||||
|
Melampyrum nemorosum L. |
+ |
+ |
||||||||
|
Melampyrum sylvaticum L. |
+ |
+ |
+ |
|||||||
|
Myosotis sylvatica Hoffm. |
3 |
+ |
+ |
|||||||
|
Orthilia secunda House |
+ |
|||||||||
|
Oxalis acetosella L. |
1 |
+ |
||||||||
|
Paris quadrifolia L. |
1 |
+ |
||||||||
|
Phleum pratense L. |
3 |
5 |
||||||||
|
Pimpinella saxifraga L. |
1 |
+ |
||||||||
|
Poa nemoralis L. |
1 |
|||||||||
|
Potentilla thuringiaca Bernh. |
+ |
|||||||||
|
Prunella vulgaris L. |
1 |
+ |
||||||||
|
Ranunculus acris L. |
+ |
|||||||||
|
Ranunculus auricomus L. |
+ |
+ |
+ |
|||||||
|
Ranunculus repens L. |
+ |
|||||||||
|
Rumex acetosa L. |
+ |
|||||||||
|
Silene viscaria (L.) Jess. |
1 |
1 |
+ |
|||||||
|
Solidago virgaurea L. |
+ |
+ |
||||||||
|
Stellaria holostea L. |
20 |
|||||||||
|
Taraxacum officinale F. H. Wigg. |
+ |
r |
+ |
+ |
||||||
|
Trifolium medium L. |
+ |
|||||||||
|
Trifolium pratense L. |
+ |
|||||||||
|
Trifolium spadiceum L. |
1 |
|||||||||
|
Urtica dioica L. |
+ |
+ |
+ |
|||||||
|
Vaccinium myrtillus L. |
+ |
+ |
5 |
1 |
||||||
|
Vaccinium vitis-idaea L. |
+ |
|||||||||
|
Veronica chamaedrys L. |
2 |
8 |
20 |
+ |
1 |
3 |
5 |
|||
|
Veronica officinalis L. |
+ |
|||||||||
|
Vicia cracca L. |
+ |
2 |
1 |
+ |
||||||
|
Vicia sepium L. |
+ |
+ |
1 |
+ |
+ |
|||||
|
Viola canina L. |
+ |
+ |
||||||||
|
Виды сем. Poaceae (скошены) |
10 |
20 |
||||||||
|
Возобновление древесных растений |
||||||||||
|
Abies balsamea (L.) Mill. |
+ |
+ |
||||||||
|
Acer platanoides L. |
+ |
3 |
3 |
|||||||
|
Cotoneaster sp. |
r |
+ |
||||||||
|
Crataegus sp. |
r |
|||||||||
|
Daphne mezereum L. |
+ |
r |
||||||||
|
Lonicera xylosteum L. |
+ |
+ |
+ |
r |
||||||
|
Prunus padus L. |
+ |
1 |
+ |
+ |
||||||
|
Quercus robur L. |
+ |
|||||||||
|
Ribes spicatum Robson |
+ |
|||||||||
|
Rubus idaeus L. |
+ |
+ |
||||||||
|
Sorbus aucuparia L. |
+ |
+ |
+ |
+ |
+ |
2 |
+ |
|||
Примечания:
-
* в соответствии с планом на рис. 1.: 1 - Pinus strobus; 2 – Picea glauca; 3 – Larix sibirica; 4 – Pinus sibirica; 5 – Abies balsamea; 6 – Abies sibirica; 7 – Tilia cordata; 8 – Acer platanoides; 9 – Ulmus laevis; 10 – Quercus robur.
-
* *В данном случае количество видов приводится без учета некоторых видов сем. Poaceae (скошены). Названия растений приводятся по The Plant List, 2013.
«+» – проективное покрытие вида менее 1%, «r» – единично.
Notes:
-
* In accordance with the plan in Fig. 1.: 1 - Pinus strobus ; 2 – Picea glauca ; 3 – Larix sibirica ; 4 – Pinus sibirica ; 5 – Abies balsamea ; 6 – Abies sibirica ; 7 – Tilia cordata ; 8 – Acer platanoides ; 9 – Ulmus laevis ; 10 – Quercus robur .
-
* * In this case, the number of species is given without regard to certain species of the family Poaceae (were mowed down). Plant names are given by The Plant List, 2013.
-
" +" – cover of the species is less than 1%, «r» – unique.
Результаты и обсуждение плотность популяций нематод в условиях
Таксономическое разнообразие и дендроинтродукции
Фауна почвенных нематод в корнеобитаемом слое растений-интродуцентов Ботанического сада представлена 51 таксоном нематод, 10 из которых являются общими всех исследованных биотопов. Это бактериотрофы родов Panagrolaimus, Rhabditis, Acrobeloides, Cervidellus, Plectus, микотрофы Aphelenchoides и Ditylenchus , политроф Eudorylaimus , нематоды, ассоциированные с растениями Filenchus , Malenchus . В группу нематод-паразитов растений входят 6 родов: полигостальные (т. е. широкоспецифичные в отношении растений-хозяев) корневые эктопаразиты Cephalenchus , Helicotylenchus , Nagelus , Tylenchorhynchus , Paratylenchus и мигрирующий эндопаразит Pratylenchus (табл. 2). Наиболее встречаемыми таксонами фитопаразитов под деревьями-интродуцентами оказались Paratylenchus , Pratylenchus , Tylenchorhynchus , что характерно, главным образом, для луговых биоценозов Карелии (Сущук и др., 2012). В местах посадок древесных растений найдены редкие для региона виды нематод ( Cephalenchus leptus , Nagelus leptus ), которые ранее были единично обнаружены в парках г. Петрозаводска, на о-ве Большой Соловецкий (Белое море) и о-ве Валаам (Ладожское озеро), и, таким образом, приурочены к центрам дендроинтродукции (Диева, Сущук, 2014; Калинкина и др., 2014; Kalinkina et al., 2015). Карантинные виды на территории Ботанического сада не выявлены. Обнаружение редких для региона видов служит в пользу гипотезы о проникновении новых и распространении редких видов фитопаразитических нематод при интродукции растений в экосистемы Севера.
Таблица 2. Список таксонов нематод, выявленных в верхнем корнеобитаемом слое почвы в местах посадок деревьев-интродуцентов на территории Ботанического сада ПетрГУ.
Table 2. Check-list of nematode taxa in upper soil layer of investigated biotopes in the Botanic Garden of Petrozavodsk State University.
Присутствие/отсутствие таксона в биоценозе
|
Таксон |
ТГ * |
СР ** |
1 *** |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
Acrobeloides |
Б |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Achromadora |
Б |
2 |
+ |
+ |
+ |
+ |
— |
+ |
— |
— |
— |
|
|
Alaimus |
Б |
4 |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
|
Anaplectus |
Б |
2 |
- |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
- |
|
Bastiania |
Б |
3 |
— |
— |
— |
— |
+ |
|||||
|
Cephalobus |
Б |
2 |
+ |
+ |
- |
+ |
+ |
- |
- |
+ |
- |
+ |
|
Ceratoplectus |
Б |
2 |
- |
+ |
- |
- |
- |
- |
- |
+ |
- |
- |
|
Cervidellus |
Б |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Chiloplacus |
Б |
2 |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Cylindrolaimus |
Б |
3 |
- |
- |
- |
- |
+ |
- |
- |
+ |
- |
+ |
|
Eucephalobus |
Б |
2 |
+ |
+ |
+ |
+ |
— |
+ |
+ |
+ |
+ |
|
|
Eumonhystera |
Б |
2 |
+ |
+ |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
|
Heterocephalobus |
Б |
2 |
+ |
- |
- |
- |
- |
+ |
- |
+ |
+ |
+ |
|
Macrolaimus |
Б |
2 |
— |
+ |
— |
— |
— |
— |
||||
|
Mesorhabditis |
Б |
1 |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
|
Metateratocephalus |
Б |
3 |
- |
- |
- |
- |
- |
- |
+ |
+ |
- |
- |
|
Monhystrella |
Б |
2 |
— |
— |
— |
— |
+ |
— |
— |
|||
|
Panagrolaimus |
Б |
1 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Plectus |
Б |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Prismatolaimus |
Б |
3 |
— |
+ |
— |
+ |
— |
+ |
— |
+ |
||
|
Rhabditis |
Б |
1 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Teratocephalus |
Б |
3 |
- |
- |
- |
- |
+ |
- |
- |
- |
+ |
- |
|
Theristus |
Б |
2 |
— |
— |
— |
+ |
— |
— |
— |
|||
|
Tylocephalus |
Б |
2 |
- |
+ |
+ |
- |
+ |
+ |
- |
- |
- |
- |
|
Wilsonema |
Б |
2 |
- |
+ |
- |
+ |
+ |
+ |
+ |
- |
- |
+ |
|
Cephalobidae |
Б |
2 |
— |
— |
— |
— |
+ |
— |
— |
— |
— |
— |
|
Aphelenchoides |
М |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Aphelenchus |
М |
2 |
+ |
+ |
- |
- |
- |
- |
- |
+ |
+ |
+ |
|
Ditylenchus |
М |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Diphtherophora |
М |
3 |
- |
- |
- |
+ |
- |
- |
+ |
+ |
+ |
- |
|
Tylencholaimus |
М |
4 |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
+ |
- |
|
Aporcelaimellus |
П |
5 |
— |
— |
— |
— |
+ |
— |
+ |
+ |
+ |
— |
|
Eudorylaimus |
П |
5 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Mesodorylaimus |
П |
5 |
- |
- |
+ |
+ |
+ |
- |
+ |
- |
+ |
- |
|
Clarkus |
Х |
4 |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
|
Mylonchulus |
Х |
4 |
- |
- |
- |
- |
- |
- |
+ |
- |
+ |
+ |
|
Trischistoma |
Х |
3 |
- |
- |
+ |
- |
+ |
+ |
+ |
- |
- |
- |
|
Aglenchus |
Аср |
2 |
+ |
+ |
- |
- |
- |
+ |
- |
+ |
+ |
+ |
|
Coslenchus |
Аср |
2 |
- |
+ |
- |
- |
+ |
- |
- |
+ |
+ |
+ |
|
Filenchus |
Аср |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Lelenchus |
Аср |
2 |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
- |
+ |
|
Malenchus |
Аср |
2 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
Tylenchus |
Аср |
2 |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
|
Tylenchidae |
Аср |
2 |
- |
+ |
+ |
- |
+ |
- |
- |
+ |
- |
- |
|
Steinernema |
Пб |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
|
Cephalenchus |
Пр |
3 |
- |
- |
- |
- |
- |
- |
- |
+ |
- |
- |
|
Helicotylenchus |
Пр |
3 |
- |
- |
+ |
+ |
- |
- |
- |
+ |
- |
+ |
|
Nagelus |
Пр |
3 |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
|
Paratylenchus |
Пр |
3 |
+ |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
|
Pratylenchus |
Пр |
3 |
+ |
+ |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
|
Tylenchorhynchus |
Пр |
3 |
+ |
+ |
+ |
- |
+ |
- |
- |
- |
+ |
+ |
|
51 таксон |
24 |
32 |
22 |
25 |
31 |
25 |
29 |
33 |
28 |
30 |
Эколого-трофическая структура сообществ почвенных нематод в условиях дендроинтродукции
Исследование эколого-трофической структуры сообществ нематод показало, что бактериотрофы, микотрофы и паразиты растений являются преобладающими группами (табл. 4). В почве под хвойными культурами доминируют бактериотрофы, субдоминантами в большинстве случаев выступают микотрофы. В структуре сообществ под посадками лиственных пород снижается доля нематод-бактериотрофов (от 43.6 % до 27.1 %, в среднем) и увеличивается доля паразитов растений (от 13.8 % до 29.2 %) по сравнению с хвойными породами.
Корнеобитаемый слой почвы большинства исследованных дендроинтродуцентов ботанического сада выделяется высокой долей нематод-паразитов растений в сообществе (табл. 4), что не характерно для естественных лесных биоценозов региона, в которых показатель варьирует на уровне 0.1–3.6 % от фауны (Груздева и др., 2011). Результаты согласуются с данными, полученными для лесных питомников Польши, где фитопаразитические нематоды составляли основу нематодных комплексов (Skwiercz, 2012; Chalanska, Labanowski, 2014). В нашем исследовании больше всего корневых паразитов растений выявлено под лиственными культурами, особенно в посадках дуба черешчатого, где паразитические нематоды доминируют в сообществе; среди хвойных – в почве биотопа с сосной Веймутова. Подобное явление было отмечено при изучении фауны почвенных нематод на рекультивируемых промышленных отвалах, где численность нематод-паразитов растений в почве посадок лиственных деревьев была выше по сравнению с хвойными породами (Hanel, 2008).
Таблица 4. Эколого-трофическая структура сообществ почвенных нематод в верхнем корнеобитаемом слое почвы (n=9) в местах посадок деревьев-интродуцентов на территории Ботанического сада ПетрГУ.
Table 4. Eco-trophic nematode community structure in upper soil layer of investigated biotopes (n=9) in the Botanic Garden of Petrozavodsk State University.
|
Биотопы |
Эколого-трофическая группа нематод, % |
|||||
|
Б |
М |
П |
Х |
Аср |
Пр |
|
|
A. Хвойные породы деревьев |
||||||
|
1 |
41.31±5.01ab |
12.61±3.07a |
3.42±0.84ab |
0.68±0.40a |
13.58±1.69ac |
28.41±5.18a |
|
2 |
37.65±3.47ab |
27.88±4.37b |
1.87±0.89ab |
1.65±0.60a |
17.92±2.83ab |
13.03±3.16b |
|
3 |
41.51±3.65ab |
22.82±3.01b |
2.91±0.73ab |
1.94±0.97a |
9.71±1.66c |
21.10±2.91ab |
|
4 |
35.25±4.71a |
12.72±1.15a |
2.59±0.86ab |
1.48±1.01a |
28.81±5.54b |
19.16±1.51ab |
|
5 |
45.54±2.34b |
40.56±2.34c |
3.43±0.57a |
0.74±0.49a |
8.74±1.45c |
0.99±0.28c |
|
6 |
60.32±3.69c |
32.53±4.09bc |
1.46±0.67b |
1.88±0.74a |
3.68±0.78d |
0.13±0.13d |
|
Б. Лиственные породы деревьев |
||||||
|
7 |
40.26±3.78a |
14.39±1.90a |
10.41±1.74a |
9.74±2.19a |
6.94±1.73a |
18.26±3.90a |
|
8 |
26.90±2.24b |
14.79±2,54a |
10.08±1.94a |
1.06±0.51b |
22.05±2.67b |
25.13±1.64a |
|
9 |
23.10±3.66bc |
30.86±1.60b |
3.0±1.15b |
2.31±0.86b |
12.73±1.60c |
27.98±2.66a |
|
10 |
18.19±2.67c |
13.81±2.02a |
4.49±1.10b |
2.04±0.93b |
16.10±2.94bc |
45.35±2.99b |
Примечания:
n – объем выборки; Б – бактериотрофы, М – микотрофы, П – политрофы, Х – хищники, Аср – нематоды, ассоциированные с растениями, Пр – нематоды-паразиты растений. Значения с разными буквенными обозначениями в строках внутри разделов А и Б статистически различаются (p < 0.05).
Notes:
n is the number of replications; B – bacterial feeders, F – fungal feeders, Om – omnivores, Pr – predators, Asp – nematodes associated with plants, Pp – plant parasites. Values with different letters in rows within sections A and B are significantly different at p < 0.05.
Особое положение занимают места посадок пихты сибирской и бальзамической за счет высокого процента в структуре сообщества нематод-бактериотрофов и микотрофов и низкого – паразитов растений. Такое соотношение эколого-трофических групп в сообществах почвенных нематод характерно для лесных экосистем Республики Карелия в целом (Груздева, 2001; Груздева и др., 2006, 2011). Сильное затенение и обильный опад из неразложившейся хвои вокруг стволов привел к формированию мертвопокровных участков. Крайне низкое развитие травянистой растительности – основного источника питания фитопаразитов – обусловило низкие значения разнообразия и относительного обилия нематод данной группы.
Эколого-популяционные индексы, характеризующие сообщества почвенных нематод в условиях дендроинтродукции
Данные по фауне нематод и количественные показатели их сообществ были проанализированы с использованием индексов, общепринятых в экологической нематологии. Полученные результаты показали, что степень зрелости сообществ нематод ΣMI имеет наименьшие значения для биотопов с посадками некоторых хвойных деревьев (табл. 5). Существует точка зрения, что при неблагоприятных условиях среды в сообществах нематод начинают преобладать менее специализированные и более устойчивые виды, имеющие короткие жизненные циклы (Bongers, 1990; Yeates, 1994). На основании ΣMI значений можно предположить, что условия обитания нематод в почве корневой зоны сосны Веймутова, ели канадской и пихты сибирской неблагоприятны для почвенных организмов, в частности нематод. Под лиственными интродуцентами сообщества почвообитающих нематод более разнообразны и стабильны, т. к. в них существуют виды с различной устойчивостью к условиям среды, и расширяется спектр таксонов с высокими значениями по с-р-шкале (К-стратегов).
Это подтверждается и значениями эколого-популяционных индексов, характеризующих состояние почвенной трофической сети и степень нарушенности почвенной экосистемы (Ferris et al., 2001). Выявлено, что наиболее стабильные, сложные почвенные трофические сети с высокими значениями индекса структурирования SI свойственны ценозам с лиственными интродуцентами. Кроме того, значения индекса SI выше 50 для данных биотопов характеризуют почвенную экосистему как ненарушенную (табл. 5).
Посадки большинства хвойных культур по соотношению индексов SI (ниже 50) и EI (ниже 50) рассматриваются как нарушенные экосистемы с деградированной почвенной трофической сетью (табл. 5). Известно, что упрощенная трофическая сеть приурочена к почвенным экосистемам, испытывающим воздействие неблагоприятных факторов среды (климатических или антропогенных). Авторами концепции – Г. Феррисом с соавторами – был введен термин «базальная трофическая сеть» для обозначения примитивной сети, представленной нематодами с широкой экологической пластичностью, главным образом бактериотрофами и микотрофами. Простоту сети авторы связывают с фактором стресса, включая ограничение пищевых ресурсов, неблагоприятные условия среды или загрязнение (Ferris et al., 2001). По литературным данным для естественных хвойных лесов характерен низкий уровень обогащения почв органикой (значения EI не превышают 50), что согласуется с полученными нами результатами, но почвенные трофические сети сохраняют многокомпонентность и стабильность (высокие значения SI) (Ferris et al., 2001; Матвеева и др., 2008; Cerevkova, Renco, 2009).
Таблица 5. Эколого-популяционные индексы, рассчитанные для сообществ почвенных нематод верхнего корнеобитаемого слоя почвы (n=9) в местах посадок деревьев-интродуцентов на территории Ботанического сада ПетрГУ.
Table 5. Ecological indices of nematode communities in upper soil layer of investigated biotopes (n=9) in the Botanic Garden of Petrozavodsk State University.
|
Биотопы |
ΣMI |
SI |
EI |
CI |
|
A. Хвойные породы деревьев |
||||
|
1 |
2.35±0.04ab |
32.82±6.79ab |
39.77±3.99a |
44.32±9.74abc |
|
2 |
2.30±0.05a |
26.20±7.10ad |
53.35±3.47b |
43.90±6.43ab |
|
3 |
2.61±0.04c |
28.04±5.28a |
32.81±2.76a |
78.12±10.61c |
|
4 |
2.56±0.04bc |
52.41±7.57bc |
65.66±6.52c |
25.75±10.45a |
|
5 |
2.50±0.02ab |
50.41±5.06c |
39.52±2.61a |
59.81±4.33bc |
|
6 |
2.28±0.07ab |
12.84±4.27d |
34.22±3.02a |
70.87±7.11c |
|
Б. Лиственные породы деревьев |
||||
|
7 |
2.66±0.05a |
74.22±2.90a |
38.93±4.58a |
47.54±8.43a |
|
8 |
2.47±0.03b |
69.94±5.09ab |
66.35±5.21b |
30.00±8.23a |
|
9 |
2.70±0.06a |
79.02±4.20a |
70.03±5.30b |
28.37±10.16a |
|
10 |
2.45±0.03b |
55.42±6.46b |
56.41±4.13b |
43.49±6.69a |
Примечания:
n – объем выборки; ΣMI – индекс зрелости сообществ нематод, SI – индекс структурирования, EI – индекс обогащения почвенной трофической сети, CI – индекс преобладающего пути разложения органического вещества в почве. Значения с разными буквенными обозначениями в строках внутри разделов А и Б статистически различаются (p < 0.05).
Notes:
n is the number of replications; ΣMI – maturity index; SI, EI, CI – structure, enrichment and channel indices of soil food web. Values with different letters in rows within sections A and B are significantly different at p < 0.05.
Индекс CI, описывающий процесс разложения органики, показал варьирование значений независимо от породы интродуцентов с тенденцией увеличения значений в почве под хвойными породами (табл. 5). Самые высокие значения (60–78) определены в местах произрастания Larix sibirica , Abies balsamea и Abies sibirica , что позволяет сделать вывод о значительной активности почвенных грибов в деструкции органики в данных биотопах. Причиной может быть накопление в подстилке трудноразлагаемых растительных остатков, трансформация которых осуществляется в основном грибами (Звягинцев и др., 2005). Под остальными интродуцентами индекс имел как низкие
( Pinus sibirica , Acer platanoides , Ulmus laevis ), так и средние ( Pinus strobus , Picea glauca , Tilia cordata , Quercus robur ) значения, т. е. разложение органики происходит при доминировании бактериальной компоненты или сбалансированном участии двух групп.
Заключение
Результаты исследования показали, что разнообразие фауны нематод в почве под широколиственными породами выше, чем в местах произрастания хвойных деревьев. На территории Ботанического сада найдены виды нематод-фитопаразитов ( Cephalenchus leptus , Nagelus leptus ), ранее единично встреченные в регионе, обнаружение которых служит в пользу гипотезы о проникновении новых и распространении редких видов фитопаразитических нематод при интродукции растений в экосистемы Севера.
В почве под всеми исследованными хвойными культурами доминирующей группой являются бактериотрофы, субдоминантами в большинстве случаев выступают микотрофы. В посадках лиственных пород снижается доля нематод-бактериотрофов и увеличивается доля паразитов растений по сравнению с хвойными. Количественное преобладание корневых паразитов растений в фауне выявлено под дубом черешчатым, что может быть связано с произрастанием растений за пределами северной границы естественного ареала и малопригодными для вида почвенными условиями. В целом, высокий процент паразитов растений является отличительной особенностью структуры сообществ почвенных нематод корнеобитаемого слоя в местах посадок растений-интродуцентов, тогда как естественные лесные биоценозы Республики Карелия характеризуются низким вкладом фитопаразитов в фауну. Данный факт вызывает опасения причинения вреда растениям и требует дальнейшего мониторинга нематологической ситуации в Ботанических садах.
На основе анализа фауны нематод и расчета эколого-популяционных индексов можно охарактеризовать почвенную трофическую сеть под посадками лиственных интродуцентов как структурированную, а почвенную экосистему – как ненарушенную. Под большинством хвойных культур почвенная трофическая сеть определена как упрощенная, функционирующая в условиях действия неблагоприятных факторов среды, а почвенная экосистема – как высоко нарушенная. Особенности биотопов с хвойными интродуцентами могут быть обусловлены локальными условиями местообитаний (близкое залегание коренных пород в сочетании с малой глубиной плодородного слоя и др.), антропогенной нагрузкой или комплексом факторов.
Благодарности
Авторы выражают признательность студентам эколого-биологического и агротехнического факультетов ПетрГУ (А. В. Марковскому, Н. А. Камаевой) за предоставление данных об уровне кислотности почв. Измерения выполнены в рамках курсовых работ и летней практики (руководитель к.с.-х.н. Л. А. Кузнецова).
Исследования выполнены в рамках государственного задания (темы № 0221-2014-0004, 0221-2015-0006) при частичной финансовой поддержке РФФИ (грант 15-04-07675_а) и Программы стратегического развития ПетрГУ на 2016 год.
