Соподчиненность почвенных условий и растительных сообществ в сосняках как следствие действия комплекса факторов

Автор: Надпорожская Марина Алексеевна, Зубкова Елена Владимировна, Фролов Павел Владимирович, Быховец Сергей Станиславович, Чертов Олег Георгиевич
Журнал: Вестник Тверского государственного университета. Серия: Биология и экология @bio-tversu
Рубрика: Ботаника
Статья в выпуске: 2, 2018 года.
Бесплатный доступ
Изучены условия формирования соподчиненных пространственных единиц почвенно-растительного покрова в сосняке кустарничковом Отрадинского лесничества Московской области. Мозаика почв представлена подзолами иллювиально-железистыми с торфяно-подзолами иллювиально-гумусовыми. Живой напочвенный покров состоит из синузий бруснично-зеленомошных, бруснично-чернично-зеленомошных и чернично-сфагновых. Комплексность почв и нижнего растительного яруса определяют постоянно действующие факторы: дюнно-бугристый микрорельеф и поверхностные четвертичные отложения (кварцевые пески на водоупорных породах). Приведены количественные показатели для лесной подстилки: мощность, зольность и отношение валовых C/N. Внешние факторы: рекреация (тропиночная сеть), низовые пожары и повреждение лесной подстилки кабанами и лосями пока не оказывают существенного влияния на структуру почвенно-растительного покрова. Практическое применение результатов: 1) валидация математической модели динамики живого напочвенного покрова CAMPUS-S; 2) мониторинг и нормирование рекреационной нагрузки в сосновых лесах Подмосковья.
Сосновый лес, подзол, лесная подстилка, живой напочвенный покров, брусника, черника
Короткий адрес: https://sciup.org/146279500
IDR: 146279500 | УДК: 574.42
Factors of soil and ground vegetation formation in pine forests
The analysis of a complex combination of natural and anthropogenic factors affecting the formation and functioning of ground vegetation and soils in the Scots pine ( Pinus sylvestris L.) forest of the experimental site “Otradinsky forestry” in the Moscow Region is presented. The mosaic of soils is represented by podzols illuvial-ferruginous (Albic Podzols (Arenic) and peat-podzols illuvial-humus (Albic Folic Gleyic Podzols (Arenic)). The ground vegetation cover is represented by associations of cowberry-green mosses, cowberry-blueberry-green-mosses and blueberry-sphagnum. The aim of the work was to determine the leading factors in the formation of this complex soil-vegetation cover. A regular set of 0.25x0.25 m sample plots and soil pits 0.3 m deep were made at the study area. Permanent and random factors are identified. As permanent factors determining the complexity of soils and the understorey vegetation, there are: micro-relief (dune-bumpy) and the structure of surface Quaternary deposits (quartz sands are underlain by water-resistant materials). Random factors include: recreation (a path network), ground fires and damage of the forest floor by wild boars. Analysis of the materials of the geobotany field study showed that random factors do not yet have a significant effect on the soil-vegetation cover structure.
Текст научной статьи Соподчиненность почвенных условий и растительных сообществ в сосняках как следствие действия комплекса факторов
Введение. Данная статья продолжает серию публикаций по созданию комплекса математических моделей EFIMOD (Komarov et al., 2003; Моделирование..., 2007) и CAMPUS (Комаров и др., 2015; Фролов и др., 2015) для оценки последствий разных режимов лесопользования, начатых А.С. Комаровым и ОТ. Чертовым (Komarov et al., 2003; Жукова и др., 2016) с сотрудниками лаборатории моделирования экосистем института физико-химических и биологических проблем почвоведения. В лесной экосистеме наиболее сложен для математической формализации живой напочвенный покров. Травяно-кустарничковая растительность быстрее реагирует на смену экологических условий (Копцик и др., 2001). Мозаичность является следствием неоднородности исходных условий среды, различий биологических свойств совместно встречающихся видов-доминантов (Раменский, 1924) и сукцессионных процессов. Например, во многих сосновых лесах Московской области из-за формирования второго древесного яруса из березы и ели начал изменяться видовой состав растений напочвенного покрова (Сукцессионные процессы, 1999; Рысин, 2009).
Вопросы взаимообусловленного пространственно-временного изменения почв и растительности являются актуальной научной проблемой, разработка которой необходима для прогноза динамики лесных экосистем (Иванов, Шадриков, 2010). Доля напочвенного покрова в общих биомассе и опаде леса играет важную роль в биологическом круговороте отдельных элементов, дефицитных в лесных экосистемах. Например, в сосновых лесах на кварцевых песках доля напочвенного покрова в общем опаде составляет около 10%, но при этом с опадом кустарничков и трав может поступать более 30% азота (Родин, Базилевич, 1965; Emmer, 1995).
Для получения более реалистичных прогнозов нужна математическая имитация взаимодействия всех элементов экосистемы, в частности и живого напочвенного покрова. Работы в этом направлении активно проводятся (Смирнова, Чумаченко, 2012; Логофет, Маслов, 2018).
Разработана модель динамики травяно-кустарничкового яруса CAMPUS-S (Фролов, Зубкова, 2017), выполнена ее верификация, проанализировано влияние на развитие кустарничков (черники и брусники) разных факторов их среды обитания (Зубкова и др., 2017). Для валидации программы CAMPUS-S требуется детальная документация связи пространственной локализации сообществ кустарничков и эдафических условий среды. Цель работы - анализ некоторых условий произрастания кустарничков черники и брусники в сосновом лесу.
Методика. Полевые работы проведены в 2015-2018 годах. В сосновом лесу с комплексным напочвенным растительным покровом (чередование участков доминирования черники, брусники, и со-доминирования черники и брусники) на пробных площадях выполнено детальное изучение растительности и почв. В местах с доминированием кустарничков заложены ключевые участки в трехкратной повторности на ненарушенных местах вдали от тропинок. Для определения изменения биомассы кустарничков по градиенту изменения условий влажности вдоль одной из пробных площадей с интервалом в 1 метр было взято 26 площадок 0.25x0.25 м. Глубина взятия монолитов на площадках 0,3 м. Надземные и подземные органы исследуемых видов кустарничков были разделены по фракциям биомассы, отмыты, высушены до абсолютно сухого состояния и взвешены.
Базовые почвенные разрезы заложены на основных элементах катены дюнно-бугристо-западинного комплекса рядом с ключевыми участками исследований. Дополнительно прикопками (по 30 шт. около каждого разреза) уточнены мощности поверхностных горизонтов. Морфология минеральных горизонтов почв описана по стандартным процедурам (Розанов, 2004). Лесную подстилку разделяли на подгоризонты, различающиеся по степени трансформации опада, используя международные индексы: L - слаборазложенный, F -ферментированный и Н - гумифицированный. Эти индексы соответствуют О’, О” и О’” в Классификации почв России В отобранных почвенных образцах для каждого из 9 участков определены: зольность (прокаливание при 900 °C), а также содержание органического углерода (С) и валового азота (N) на анализаторе CHNS-932 фирмы Leco (США).
Статистическую обработку данных проводили в программах Microsoft Excel 2010 и IBM SPSS Statistics. На диаграммах приведены средние значения и стандартное отклонение среднего (± SD, отмечены барами).
Объекты исследования. Общая характеристика ключевого участка. Ключевой участок выбран в сосновом лесу Отрадинского участкового лесничества (ОУЛ) опытного лесного хозяйства «Русский лес» (географические координаты трех участков исследований: 54°54Т0"с.ш., 37°39'56"в.д.; 54°54,09"с.ш„ 37°39'57" в.д.; 54°54'09" с.ш., 37°40'02"в.д.). Здесь террасы Оки, сложенные плотными известняками каменноугольного периода, перекрыты днепровской мореной и аллювиально-флювиогляциальными кварцевыми песками. Мощность песчаных отложений от 0,75 м до 2 м, рельеф слабо волнистый (Лидов, 1949, цит. по: Ханина, Бобровский, 2004). Почвы, расположенного рядом, Приокско-Террасного заповедника были подробно описаны В.М. Фридландом (1955). Комплексность почвенно-
Вестник ТвГУ. Серия "Биология и экология". 2018. № 2. растительного покрова определяют особенности микрорельефа. Важным абиогенным фактором функционирования этих ландшафтов является различная дренированность, обусловленная расчлененностью рельефа, уклоном поверхности и относительно небольшой мощностью песков (Иванов, Шадриков, 2010).
Лимитируют развитие почвенно-растительного покрова этих местообитаний - дефицит элементов минерального питания растений почвообразующих песков и контрастный режим увлажнения бугристодюнного рельефа с сухими повышенными элементами мезорельефа и периодическим избыточным увлажнением в понижениях. Грубогумусные лесные подстилки, формирующиеся под преобладающими здесь сосновыми лесами, эвтрофируют почвы на кварцевых песках и сглаживают режим увлажнения. Накопление лесной подстилки и ее качество можно считать главными эдафическими факторами, от которых зависит развитие самого лабильного компонента экосистемы сосновых лесов, живого напочвенного покрова. В настоящей работе мы оценивали приоритетные показатели, характеризующие процессы аккумуляции и минерализации органического вещества этих почв: мощность органогенных горизонтов, а также отношение валовых С и N и зольность подстилок.
Растительность ключевого участка. Растительность ключевого участка представлена сосняком кустарничковым. Древесный ярус (9С1Б) образован сосной обыкновенной (Pinus sylvestris L.) 80-85 лет с примесью березы повислой (Betula pendula Roth) и березы пушистой феШ1а pubescens Ehrh.,); изредка встречаются более старые сосны 100-120 лет, оставленные для осеменения территории во время рубок в 40-50-х годах. Второй ярус отсутствует. Подрост представлен единичными елями (Picea abies (L.) Karst.) от 0,2 м до 5-6 м, березами до 7 м (Betula pubescens Ehrh., Betula pendula Roth); рябиной (Sorbus aucuparia L.) до 2-3 м, сосной от 0,2 м до 1 м, торчками дуба (Quercus robur L.) 0,5-1,5 м высотой. Из кустарников на пониженных, более влажных участках встречается ива пепельная (Salix cinerea L.). Напочвенный покров состоит преимущественно из кустарничков и мхов. На повышениях доминирует брусника (Vaccinium vitis-idaea L.). Есть отдельные участки с вереском (Calluna vulgaris (L.) Hull) и лишайниками (Cetraria islandica (L.) Ach., Cladonia rangiferina (L.) Weber ex F.H.Wigg., Cladonia furcata (Huds.) Schrad. и др.). Из мхов преобладают Polytrichum juniperinum Hedw., Dicranum polysetum Sw. Pleurozium schreberi (Brid.) Mitt. На склонах бугров, где со-доминируют черника и брусника, встречается папоротник орляк (Pteridium aquilinum (L.) Kuhn ex Decken), марьянник луговой (Melampyrum pratense L.), единично вейник лесной (Calamagrostis arundinacea (L.) Roth). Понижения занимают около 5% от общей площади ключевого участка. Здесь древесный ярус образует окна, по контуру занятые подростом березы и ивой козьей.
Напочвенный покров таких участков представлен черникой (Vaccinium vitis-idaea L.) с участием голубики обыкновенной (Уасстшт uliginosum L.), изредка встречаются кусты багульника болотного (Ledum palustre L.). На дне понижений, затапливаемом весной талыми водами на одну-три недели, черника отсутствует. Понижения заняты мхами (Sphagnum angustifolium (C.E.O.Jensen ex Russow) C.E.O.Jensen и др.) и редкими кочками (20-25 см в диаметре) пушицы влагалищной (Eriophorum vaginatum L.). Возобновление сосны, появляющееся в летний период на этих участках весной, при затоплении верховодкой, погибает. Граница вымочек маркируется отмершими парциальными кустами черники. Названия растений даны по справочнику С.К.Черепанова (1995).
Почвы ключевого участка. На ключевом участке Отрадинского лесничества в изученном сосняке кустарничковом почвенный покров представлен сочетанием подзолов иллювиально-железистых (O-AEh-E-Bf-BC-C) с торфяно-подзолами иллювиально-гумусовыми глееватыми (O(T)-AEh-E-Bf-BHF-BF-BG-C) на аллювиально-флювиогляциальных песчано-супесчаных и щебнистых отложениях. Ниже приведено описание наиболее характерных почвенных разрезов. Дата проведения полевых работ: Об.VI 2017.
Площадка ПП9 («Брусника») расположена на верхней части плакора. Микрорельеф микрозападинный. Местами порой кабанов (лесная подстилка нарушена до минерального горизонта неровными полосами шириной 1-2 м и длиной 3-4 м. Хозяйственное использование: рекреация. Доминирующий вид травяно-кустарничкового яруса (ТКЯ): брусника. Мертвый покров занимает 60% поверхности.
О’ - 0-2,5 см - бурый, влажный, рыхлый, слаборазложившийся опад - хвоя и кора сосны, веточки, листья брусники, остатки мха.
О” - 2,5-6,5 см - темно-бурый, влажный, состоит из средне разложенных остатков мха, листьев, веточек, уплотнен, обильно — живые и мертвые корни; мицелий белого и желтого цвета.
О’” - 6,5-9 см - бурый, влажный, хорошо разложившийся; живых корней меньше, чем в О”, много отмерших корней.
АН pir - 9-10 см - черная прослойка древесных угольков неравномерной мощности (0.5-2 см).
АЕ - 10-19 см - палево-белесый сероватый бесструктурный песок.
Е - 19-33 см - белесо-желтовато-розоватый мелкий песок. Bf- 33-39 см - желтый свежий песок.
BF- 39-80 см - ржаво-желтый влажный песок с ортштейнами. Название почвы
Классификация почв СССР (1977): Подзолистая на песчаносупесчаных и щебнистых отложениях.
Классификация почв России (2004): Подзол иллювиально- железистый.
Форма гумуса (Чертов, 1981): грубогумусная (постпирогенный грубый гумус).
World Reference Base (2014): Albie (Arenic) Podzol.
Площадка ПП8 («Брусника-черника»). Общий характер рельефа: площадка расположена на плакоре с понижением примерно 10°. Положение в рельефе: верхняя часть склона. Микрорельеф: микрозападины и приствольные повышения. Влияние животных: порой кабанов. Хозяйственное использование: рекреация.
Доминирующий вид ТКЯ: брусника и черника. Мертвый покров: выражен (20%).
Описание почвенного профиля:
О’ - 0-2 см - бурый влажный рыхлый слаборазложившийся опад — хвоя и кора сосны, веточки, листья брусники, остатки мха
О” - 2-6 см - темно-бурый, влажный, состоит из средне разложенных остатков мха, листьев, веточек, уплотнен, обильно -живые и мертвые корни.
О’” -6-8 см - бурый влажный хорошо разложившийся опад, живых корней меньше, много отмерших корней.
АН pir - 8-9 см - черная прослойка мелких древесных угольков с отдельными вкраплениями крупных древесных углей, неравномерной мощности (0.5-2 см).
АЕ - 9-20 см - светло-серый свежий песок, с черными пятнами древесных углей, переход в следующий горизонт постепенный.
Е - 20-38 см - светло-палевый песчаный плотноватый связанный с редкими корнями.
BF] - 38-60 см - охристый свежий песок, толстые корни.
BF2 - 60-110 см - палево-желтый влажный песок с орштейнами, корней мало. Уровень грунтовых вод ПО см.
Название почвы.
Классификация почв СССР (1977): Подзолистая.
Классификация почв России (2004): Подзол иллювиальножелезистый. Форма гумуса (Чертов, 1981): грубогумусная (постпирогенный грубый гумус).
World Reference Base (2014): Albie Podzol (Arenic).
Площадка ПП7 («Черника»). Положение в мезорельефе: нижняя часть склона в понижении между буграми. Микрорельеф: западинный. Влияние животных: не отмечено. Хозяйственное использование: рекреация. К западу от разреза находится микро-понижение с погибшими парциальными кустами черники. Доминирующий вид ТКЯ: черника. Мертвый покров: выражен (5%).
О’ (Т’) - 0-3 см - светло-бурый, мокрый, рыхлый, состоит из слабо разложенных остатков мхов (сфагнумы и зеленые мхи), встречаются листья, веточки.
О” (Т”) - 3-8 см - бурый, мокрый, состоит из средне разложенных остатков мхов (сфагнумы и зеленые мхи), листьев, веточек, уплотнен, обильно - живые и мертвые корни.
О’” (Т”) - 8-15 см коричнево-серый хорошо разложенный торф (перегной), сырой, корни мертвые и живые, переход резкий.
Е - 15-23 см - палево-белесый влажный бесструктурный песок, уплотнен, неравномерный по мощности, корни редкие, переход резкий, неровный, затеками.
ВН - 23-60 см - кофейно-коричневый влажный непрочнокомковатый песок, плотный, переход постепенный.
BCG - с 60 см - желтый с сизым оттенком влажный бесструктурный песок, плотный.
Уровень грунтовых вод в разрезе 50 см.
Название почвы.
Классификация почв СССР (1977): Торфянисто-подзолистая грунтово-оглеенная.
Классификация почв России (2004): Торфяно-подзол иллювиально-гумусовый глеевый.
Форма гумуса (Чертов, 1981): Торфянисто-перегнойная.
World Reference Base (2014): Albie Folic Gleyic Podzol (Arenic).
Результаты и обсуждение. Анализ комплексности почвенного покрова. На изученной пробной площади соснового леса Отрадинского лесничества подзолы иллювиально-железистые приурочены к плоским вершинам и пологим склонам повышенных элементов бугристодюнного рельефа (около 95% площади). В понижениях между буграми-дюнами находятся почвы с оторфованными лесными подстилками, мощность которых немногим более 10 см. Перепад высот между вершинами бугров и днищами понижений составляет 1,0 — 1,5 м. Поверхностные подподстилочные минеральные горизонты содержат древесные угольки, маркирующие прошедшие лесные пожары. Бугристо-дюнный мезорельеф препятствовал распахиванию этой территории, поэтому эти лесные массивы подвергались только действию рубок, пожаров и рекреации. Почвы изученного ключевого участка ОУ Л не имеют следов окультуривания в виде остаточного пахотного горизонта и не были в сельскохозяйственном использовании.
Мощность лесной подстилки зависит от возраста соснового леса (в нашем случае - времени, прошедшего после верхового пожара), количества и качества поступающего опада и условий его трансформации (режимы температуры и влажности). Средообразующая роль лесной подстилки в бореальных лесах определяется ее высокой водоудерживающей способностью и аккумуляцией зольных элементов и азота (Wilde, 1958; Чертов, 1981; Телеснина, 2018).
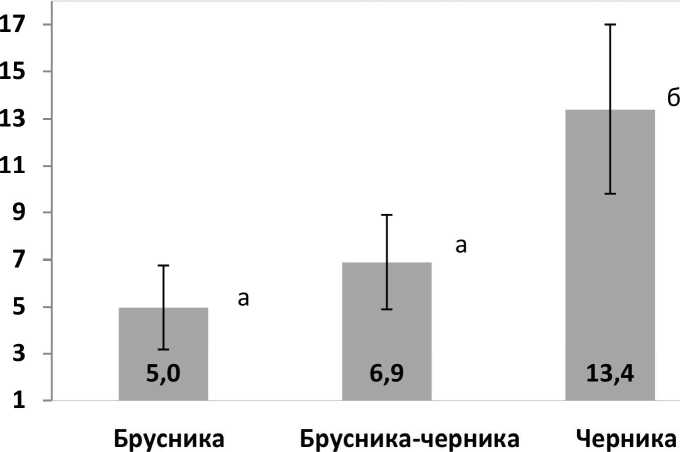
В сосняках зеленомошных дренированных местообитаний тайги равновесная мощность лесных подстилок восстанавливается через 125 лет после верхового пожара и составляет 7,5-8,5 см (Горшков и др., 2005). Соответственно, в 80-летнем сосновом лесу мощность подстилки может составлять 6,3-7,4 см. Средняя мощность лесных подстилок (рис. 1а) площадки брусники 5,0 см, что на 26-48% меньше. Мощность подстилки площадки брусника-черника 6,9 см соответствует среднему значению в предполагаемом диапазоне 6,3-7,4 см. Вероятно, что верхние части микрорельефа подвергаются действию какого-то фактора, тормозящего аккумуляцию подстилочных горизонтов. Это могут быть локальные низовые пожары, следы которых были отмечены на стволах деревьев и в почве неподалеку от ключевого участка. Либо это воздействие рекреации, когда вытаптывание приводит к уплотнению подстилки и уменьшению ее мощности. Порой животных также могут изменять нормальный ход аккумуляции подстилок, поскольку рыхление и перемешивание с минеральным горизонтом ускоряет минерализацию органического вещества. Пока действие этих факторов нельзя признать существенным из-за варьирования показателей — дисперсионный анализ показал отсутствие достоверных различий по мощности лесных подстилок на площадках с доминированием брусники и со доминированием брусники и черники.
Средняя мощность оторфованных подстилок под синузиями черники согласно дисперсионному анализу достоверно больше (13,4 см), формируется она за счет сфагновых мхов и повышенной влажности.
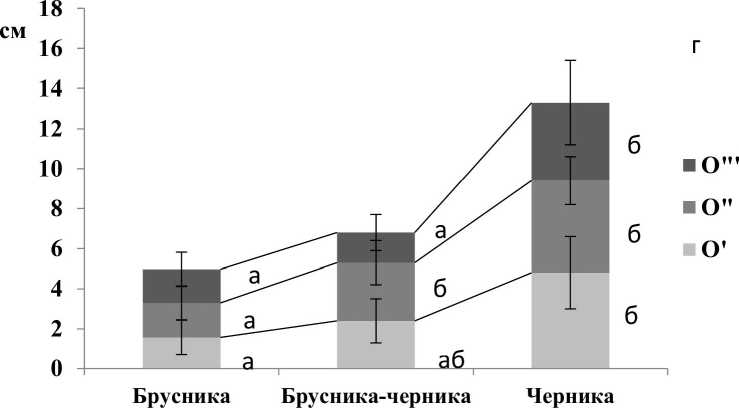
Подстилки всех трех участков разделяются на три слоя, соответствующих стадиям трансформации растительного материала. Можно отметить тенденцию увеличения мощности слоев подстилки от вершины бугра к понижению (рис. 2а). Достоверность различий представлена на рисунке 26.
Зольность и отношение C/N являются показателями интенсивности процессов минерализации лесной подстилки. В подстилках площадок брусника-черника и черника зольность максимальна в подгоризонтах О’” (рис. 2) и соответствует степени разложения растительного материала. Зольность всех слоев подстилки площадки брусника несколько повышена, по сравнению с другими двумя площадками. Кроме этого, в этой подстилке нет возрастания зольности от О’ к О’”. Вероятно, это последствия роющей активности животных. Отмеченные различия статистически недостоверны.
см
а
б
Рис. 1 . Мощность лесной подстилки на участках с доминированием кустарничков: а) общая мощность, б) подгоризонтов лесной подстилки (цифры показывают среднее из 28 повторностей): идентичные буквы обозначают отсутствие достоверных различий между средними значениями (критерий Стьюдента-Ньюмена-Келса, Р< 0,05).
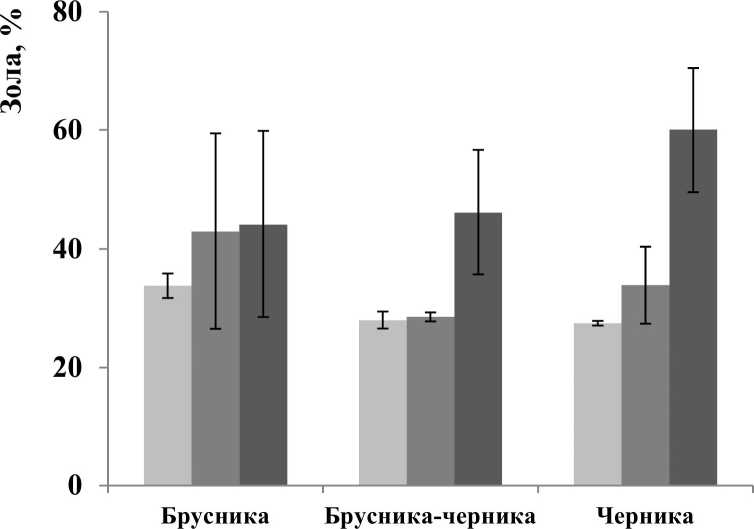
Рис. 2. Зольность по подгоризонтам лесных подстилок (обозначения как на рис. 1)
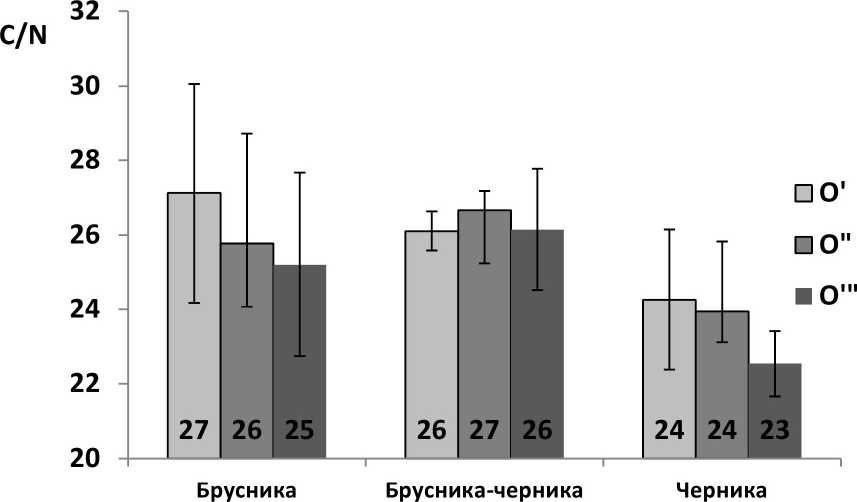
Рис. 3 . Отношения валовых C/N по подгоризонтам лесной подстилки
Отношения валовых С и N по подгоризонтам лесной подстилки (рис. 3) колеблется вблизи нижнего диагностического показателя грубого гумуса (C/N 25, по: Чертов, 1981). Различия по участкам и подгоризонтам по показателю C/N недостоверны.
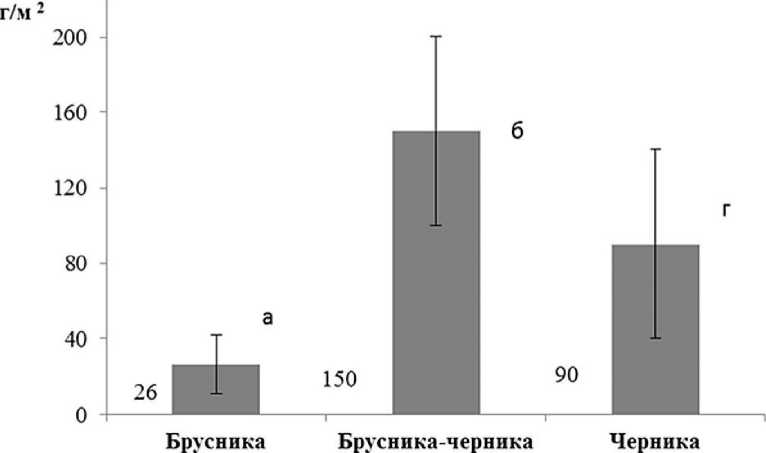
Биомасса живого напочвенного покрова. Участки с доминированием брусники, расположенные на верхних элементах рельефа, характеризуются минимальным накоплением биомассы кустарничков; участки с доминированием черники в нижних частях склонов имеют запасы биомассы почти в два раза больше; запасы биомассы кустарничков максимальные на промежуточной территории, где два вида со-доминируют (рис. 4 а).
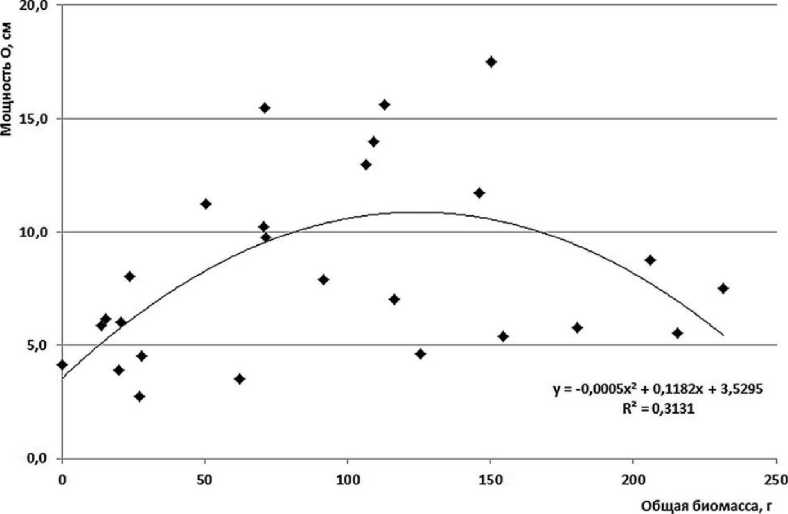
Корреляционный анализ показывает, что связи между показателями биомассы и мощностью лесной подстилки нет (рис. 4 б). Изученные показатели качества лесной подстилки (отношения C/N и зольность) тоже не имеют достоверных отличий. Таким образом, эффект максимальной суммарной биомассы кустарничков при их со-доминировании является следствием разного пространственного размещения органов растений: подземные побеги (корневища) и корни брусники чаще располагаются в минеральном горизонте и на границе подстилки с ним, в то время, как у черники, подземные органы расположены выше и занимают преимущественно ферментативный и гумусовый слои подстилки. Парциальные побеги черники образуют ярус на высоте 15-20 см, где и расположена основная фотосинтезирующая масса листьев. Черника относится к листопадным кустарничкам с симподиально ветвящимися побегами, дающим листья на молодых ежегодно отрастающих веточках. В отличие от нее побеги брусники высотой 10-15 см более компактны и более равномерно покрыты листьями; брусника не сбрасывает листья на зиму, длительность их жизни составляет 3-4 года. Найденные проявления динамики продуктивности кустарничкового яруса имеют важное биологическое значение. Потребности брусники и черники к условиям среды, выявленные по экологическим шкалам (Цыганов, 1983; Зубкова и др., 2008) в целом довольно похожи, но имеют и некоторые отличия. Были проведены подробные исследования, которые будут детально рассмотрены в отдельных публикациях.
Итак, ведущими внутренними экосистемными факторами, приводящими к формированию комплексного почвенного покрова из подзолов иллювиально-железистых и торфяно-подзолов иллювиальногумусовых глеевых на территории опытного участка Отрадинского лесничества Московской области являются бугристо-дюнный рельеф и комплекс почвообразующих и подстилающих пород. Сочетание этих факторов приводит к формированию неравномерного увлажнения элементов микрорельефа, что в свою очередь вызывает дифференциацию поверхностных горизонтов почв и дифференциацию напочвенного покрова: синузии зеленых мхов и брусники приурочены к повышенным элементам, а сфагновые мхи и черника занимают понижения. Промежуточные элементы микрорельефа заняты зелеными мхами, черника и брусника доминируют при незначительном участии трав. При этом по изученным характеристикам (зольности и отношению C/N) лесные подстилки дюнно-бугристого комплекса достоверных различий не имеют.
а
б
Рис. 4. Биомасса кустарничков (а) и корреляция биомассы кустарничков и мощности лесной подстилки (б).
Анализ действия внешних факторов на почвенно-растительный покров. Как уже отмечено выше, комплексность почвеннорастительного покрова, обусловленная эдафическими факторами, усложняется влиянием внешних факторов природного и антропогенного происхождения. Во-первых, это низовые пожары разной мощности, чаще затрагивающие вершины бугров. Во всех изученных подзолах отмечено наличие древесных углей под лесной подстилкой. В торфяно-подзолах подподстилочный слой древесных угольков отмечен на одной площадке из трех. Понижения между буграми чаще повреждаются вымочками из-за застаивающихся талых вод. Причина нарушения лесной подстилки кабанами и лосями -произрастание здесь трюфеля зернистого (Elaphomyces granulatus Fr.). Кабанами подстилка «пропахивается» полосами шириной около 1 -2 м и длиной до 3-4 метров, что создает открытые участки почвы, благоприятные для возобновления древесных пород и других растений. В отличие от низовых пожаров, когда с продуктами горения теряется большая доля биогенных элементов, и трофический статус почвы понижается, животные являются только факторами биогенной педотурбации.
Важным внешним фактором может быть рекреация. Пробная площадь соснового леса в ОУЛ находится в буферной зоне Приокско-Террасного заповедника, весь летний сезон здесь идет активный сбор грибов и ягод. Чрезмерная рекреационная нагрузка может приводить к деградации лесных биогеоценозов, обеднению биоразнообразия и нарушению почвенного покрова. Особенно высока чувствительность к рекреации у поверхностных почвенных горизонтов. Согласно «Отраслевым стандартам...» (ОСТ 56-100-95) на обследованной нами территории Отрадинского лесничества уровень рекреационной нагрузки соответствует первой-второй стадиям дигрессии: доля нарушенной тропинками площади около 3-5%. Пока тропиночная сеть не оказывает значительного влияния на экосистему соснового леса, хотя несколько уменьшает увлажненность повышенных элементов микрорельефа. Территория, вследствие ее близости к городским агломерациям, нуждается в регулярном контроле ее экологического состояния.
Заключение. В настоящее время основное влияние на формирование мохово-кустарничкового яруса и почв ключевого участка ОУ Л оказывают постоянно действующие экосистемные факторы: микрорельеф (бугристо-дюнный) и комплекс почвообразующих и подстилающих пород (аллювиальнофлювиогляциальные пески на днепровской морене). Разная мощность песчаных отложений создает дифференциацию режимов влажности почв, что влияет на видовой состав растений нижних ярусов леса и скорость разложения опада. Все внешние факторы: вымокание понижений при весеннем застаивании вод, пожары, действие животных и рекреация, пока не оказывают достоверно определяемое влияние на растительность.
Проведенное исследование позволило представить картину ведущих факторов образования изученного комплексного почвеннорастительного покрова, дать количественную оценку ключевой площади по биомассе кустарничков и параметрам подстилки. Полученные данные будут использованы для создания алгоритмов моделирования участков леса с разными экологическими условиями роста растений, а также для задач мониторинга состояния и прогноза перспектив рационального природопользования.
Авторы благодарят руководство ГКУМО «Русский лес» за возможность проводить исследования; Н.В. Филиппову (ФГБОУ ВО «ЮГУ») за помощь в определении грибов, а также в.н.с., к.б.н. А.М.Ермолаева, м.н.с. Г.Г. Фролову и Е.П.Стадник за помощь в этапе полевых работ. Почвенные анализы выполнены в ЦКП ИФХиБПП РАН.
■institute of Physicochemical and Biological Problems in Soil RAS, Puschino 3University of Applied Sciences Bingen, Bingen (Germany)
The analysis of a complex combination of natural and anthropogenic factors affecting the formation and functioning of ground vegetation and soils in the Scots pine (Pinus sylvestris L.) forest of the experimental site “Otradinsky forestry” in the Moscow Region is presented. The mosaic of soils is represented by podzols illuvial-ferruginous (Albie Podzols (Arenic) and peat-podzols illuvial- humus (Albie Folic Gleyic Podzols (Arenic)). The ground vegetation cover is represented by associations of cowberry-green mosses, cowberry-blueberry-green-mosses and blueberry-sphagnum. The aim of the work was to determine the leading factors in the formation of this complex soil-vegetation cover. A regular set of 0.25x0.25 m sample plots and soil pits 0.3 m deep were made at the study area. Permanent and random factors are identified. As permanent factors determining the complexity of soils and the understorey vegetation, there are: micro-relief (dune-bumpy) and the structure of surface Quaternary deposits (quartz sands are underlain by water-resistant materials). Random factors include: recreation (a path network), ground fires and damage of the forest floor by wild boars. Analysis of the materials of the geobotany field study showed that random factors do not yet have a significant effect on the soil-vegetation cover structure. Keywords: pine forest, podzol, ground vegetative cover, cowberries, blueberries.
Об авторах:
БЫХОВЕЦ Сергей Станиславович - кандидат географических наук, ведущий научный сотрудник лаборатории моделирования экосистем, ФГБУН Институт физико-химических и биологических проблем почвоведения РАН, 142290, Московская область, Пущино, ул. Институтская, д. 2; e-mail: sbykhovets @r amb ler.ru
Надпорожская M.A. Соподчиненность почвенных условий и растительных сообществ в сосняках как следствие действия комплекса факторов / М.А. Надпорожская, Е.В. Зубкова, П.В, Фролов, С.С. Быховец, ОТ. Чертов // Вести. ТвГУ. Сер. Биология и экология. 2018.№ 2. С. 122-138.
Список литературы Соподчиненность почвенных условий и растительных сообществ в сосняках как следствие действия комплекса факторов
- Горшков В.В., Ставрова Н.И., Баккал И.Ю. 2005. Динамика восстановления лесной подстилки в бореальных сосновых лесах после пожаров//Лесоведение. № 3. С. 37-45.
- Жукова Л.А., Нотов А.А., Зубкова Е.В, Паленова М.М. 2016. Роль А.С. Комарова в развитии популяционно-онтогенетического направления//Вестник ТвГУ. Серия «Биология и экология». № 4. С. 279-328.
- Зубкова Е.В., Ханина Л.Г., Грохлина Т.И., Дорогова Ю.А. 2008. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoScaleWin: Учебное пособие./Мар. гос. ун-т, Пущинский гос. ун-т. -Йошкар-Ола: МарГУ. 96 с.
- Зубкова Е.В., Фролов П.В., Лянгузова И.В., Шанин В.Н., Быховец С.С. 2017. Экосистемные функции кустарничков: модельный подход к их оценке // Математическое моделирование в экологии // Материалы Пятой Национальной научной конференции с международным участием (Пущино, 16-20 октября 2017 года). Пущино, ИФХиБПП РАН. С. 91-94.
- Иванов И.В., Шадриков И.Г. 2010. Коэволюция почв и растительного покрова в южной тайге (на примере Приокско-Террасного заповедника)//Почвоведение. № 11. С. 1326-1333.
- Классификация и диагностика почв России. 2004/Авторы и составители: Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. М.: Почвенный институт им. В.В. Докучаева. 342 с.
- Классификация и диагностика почв СССР. 1977/Егоров В. В., Иванова Е. Н., Фридланд В. М. М.: Колос. 225 с.
- Комаров А.С., Зубкова Е.В., Фролов В.П. 2015. Клеточно-автоматная модель динамики популяций и сообществ кустарничков//Сибирский лесной журнал. Вып. 3. С. 57-69
- Копцик Г.Н., Багдасарова Т.В., Горленко О.В. 2001. Взаимосвязи видового разнообразия растений и свойств почв в экосистемах южной тайги//Бюл. МОИП. Отд. биол. Т. 106. Вып. 2. С. 31-38.
- Кузнецов В.А., Рыжова И.М., Стома Г.В. 2017. Изменение свойств почв лесопарков Москвы при высоком уровне рекреационной нагрузки//Почвоведение. № 10. С. 1270-1280.
- Моделирование динамики органического вещества в лесных экосистемах. 2007/отв. ред. В.Н. Кудеяров. М.: Наука. 280 с.
- ОСТ 56-100-95. 1995. Методы и единицы рекреационных нагрузок на лесные природные комплексы. М. 14 с.
- Раменский Л.Г. 1924. Основные закономерности растительного покрова и методы их изучения//Вестник опытного дела Средне-Черноземной области. Январь-февраль. Воронеж: Областная редакция издательского комитета Народного комиссариата земледелия, 1924 С. 37-73: по изд. Избранные работы. Проблемы и методы изучения растительного покрова. Л.: Наука, 1971. С. 5-33.
- Родин Л. Е., Базилевич Н.И. 1965. Динамика органического вещества и биологический круговорот в основных типах растительности. М.-Л.: Наука. 253 с.
- Розанов Б.Г. 2004. Морфология почв. М.: Академический проект. 432 с.
- Рысин Л.П. 2009. Сукцессионные процессы в лесах Центральной части Русской равнины//Успехи современной биологии. Т. 129. № 6. С. 578-587.
- Смирнова О.В., Чумаченко С.Н. 2012. Концептуальная модель динамики напочвенного покрова//Лесной вестник. №9. С. 94-102.
- Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия. 1999. Под ред. О.В. Смирновой, Е.С. Шапошникова/О. В. Смирнова, Л. Б. Заугольнова, О. И. Евстигнеев и др. Российское ботаническое общество СПб. 549 с.
- Телеснина В.М., Семенюк О.В., Богатырёв Л.Г., Бенедиктова А.И. 2018. Особенности напочвенного покрова и лесных подстилок в искусственных липовых насаждениях в зависимости от характера ухода//Вестник Московского университета. Серия 17: Почвоведение. № 2. С. 3-11.
- Фролов П.В., Зубкова Е.В. 2017. Моделирование динамики биомассы растений травяно-кустарничкового яруса лесных экосистем//Сохранение лесных экосистем: проблемы и пути их решения: материалы Всероссийской научно-практической конференции 15-19 мая 2017 г. Киров: Радуга-Пресс. С. 334-339.
- Фролов П.В., Зубкова Е.В., Комаров А.С. 2015. Клеточно-автоматная модель сообщества двух видов растений разных жизненных форм//Известия академии наук, сер. биологическая. № 4. С. 341-349.
- Ханина Л.Г., Бобровский М.В. 2004. Прогноз изменения растительного покрова в Приокско-Террасном заповеднике по лесотаксационным данным//Восточноевропейские леса: история в голоцене и современность в 2 кн./Российская академия наук, Центр по проблемам экологии и продуктивности лесов; отв. ред. О.В. Смирнова. Кн. 2. М.: Наука. С. 131-137.
- Фридланд В.М. 1955. Почвы Приокско-Террасного государственного заповедника//Материалы по географии и генезису почв лесной зоны Европейской территории СССР/Труды Почвенного ин-та им. В.В. Докучаева. М.: Изд-во АН СССР. Т. XLVI. С. 136-190.
- Фролов П.В., Зубкова Е.В., Комаров А.С. 2015. Клеточно-автоматная модель сообщества двух видов растений разных жизненных форм // Известия академии наук, сер. биологическая. №4. С. 341-349 / P. V. Frolov, E. V. Zubkova, and A. S. Komarov. 2015. A Cellular Automata Model for a Community Comprising Two Plant Species of Different Growth Forms // Biology Bulletin. Vol. 42. No. 4. Р. 279-286.
- Цыганов Д.Н. 1983. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука. 196 с.
- Черепанов С.К. 1995. Сосудистые растения России и сопредельных государств. СПб. 990 с.
- Чертов О.Г. 1981. Экология лесных земель (почвенно-экологическое исследование лесных местообитаний). Л.: Наука. 192 с.
- Emmer I.M. 1995. Humus form and soil development during a primary succession of monoculture Pinus sylvestris on poor sandy substrates. Amsterdam. 135 p.
- Komarov A.S., Chertov O.G., Zudin S.L., Nadporozhskaya M.A., Mikhailov A.V., Bykhovets S.S., Zudina E.V., Zoubkova E.V. 2003. EFIMOD 2 -A model of growth and elements cycling of boreal forest ecosystems//Ecological Modelling. V. 170. P. 373-392.
- Wilde S.A. 1958. Forest soils; their properties and relation to silviculture. New York, Ronald Press. 537 p.
- World Reference Base for Soil Resources. International soil classification system for naming soils and creating legends for soil maps. 2014. Rome: FAO. www.fao.org/3/i3794en/I3794en.pdf
