Сопряженное влияние кортизола и тестостерона на адаптационную стратегию поведения в условиях «открытого поля»

Автор: И.В. Червова, И.И. Шахматов, Ю.А. Бондарчук, П.С. Маршалкина
Журнал: Ульяновский медико-биологический журнал @medbio-ulsu
Рубрика: Биологические науки
Статья в выпуске: 4, 2024 года.
Бесплатный доступ
Цель. Изучение сопряженного влияния кортизола и тестостерона на адаптацию животных в условиях «открытого поля» с учетом их конституциональных особенностей. Материалы и методы. Изучена энтропия поведения 20 взрослых крыс-самцов линии «Вистар». Энтропия поведения рассчитывается с помощью специальной формулы на основе показателя, фиксируемого в ходе тестирования по методике И.Ю. Забродина, позволяющей оценить вероятностное присутствие тех или иных поведенческих актов, включаемых животным в поведенческий арсенал исходя из индивидуальной копинг-стратегии. В опыте осуществлялась регистрация актов дефекации и уринации, определяемых многими исследователями как внешнее проявление эмоционального напряжения. По завершении эксперимента был произведен забор крови в целях оценки концентрации кортизола и тестостерона, которая осуществлялась иммунофер-ментным методом на анализаторе Immelite DPS Cirrus Inc. Для количественной характеристики согласованности показателей был применен корреляционный анализ. Результаты. В ходе эксперимента определена конституционально заданная характеристика – энтропия поведения крыс. Комплексный характер адаптации в стрессовой обстановке проявляется через применение животными различных копинг-стратегий. Адаптация сопровождается напряжением эндокринной системы, различной интенсивностью проявления ряда вегетативных реакций. Кортизол принимает участие в развитии стрессовых реакций, вызывая повышение концентрации глюкозы в крови, определяя стратегию адаптацию организма к стрессогенным условиям. Тестостерон, являясь антагонистом кортизола, понижает эффективность его действия, выступает в роли одного из компонентов стресс-лимитирующей системы. Выводы. Концентрация кортизола взаимосвязана с проявлением отдельных поведенческих актов, которые характерны для двух разных копинг-стратегий – ограниченного применения поведенческих актов (низкий показатель энтропии поведения) и, напротив, активной перцепции при сохранении общей стратегии поисковой активности (высокий показатель энтропии поведения). Функциональный антагонизм кортизола и тестостерона на фоне вариативного рецепторного связывания дает возможность рассматривать соотношение этих гормонов как маркер эустресса.
Открытое поле, адаптация, кортизол, тестостерон, поведение, поведенческие акты, дефекация, уринация, гормоны, эустресс
Короткий адрес: https://sciup.org/14132314
IDR: 14132314 | DOI: 10.34014/2227-1848-2024-4-157-168
Combined effect of cortisol and testosterone on adaptative behaviour strategy in open field
Objective. The aim of the study is to examine the combined effect of cortisol and testosterone on animal adaptation in the open field, taking into account their constitutional characteristics. Materials and Methods. The authors examined behavioural entropy of 20 adult male Wistar rats. Behavioral entropy is calculated using a special formula based on the parameter fixed during testing according to I.Yu. Zbrodin methodology. It allows us to assess the probability of certain behavior acts used by ani-mal based on individual coping strategies. The experiment included recording defecation and urination rates, which are defined by many researchers as an external manifestation of emotional stress. Upon the experiment completion, blood was collected to assess the cortisol and testosterone levels. Blood test was carried out using the enzyme immunoassay on an Immelite DPS Cirrus Inc. analyzer. Correlation analysis was used for quantitative characteristics of agreement rates. Results. During the experiment, a constitutionally given characteristic, namely the entropy of rat behavior, was determined. The complex nature of adaptation in a stressful environment is manifested through various coping strategies used by animals. Adaptation is accompanied by tension in the endocrine system, various manifestation of vegetative reactions. Cortisol contributes to stress development, increasing the concentration of glucose in the blood, determining the strategy of body adaptation to stressful environment. Testosterone, being cortisol antagonist reduces its effectiveness, and acts as one of the components of the stress-limiting system. Conclusion. Cortisol concentration is connected with certain behaviour acts, which characterize two different coping strategies limited use of behaviour acts (low behavioral entropy) and, on the contrary, active perception under general strategy of search activity (high behavioral entropy). The functional antagonism of cortisol and testosterone against the background of variable receptor binding makes it possible to consider the ratio of these hormones as a marker of eustress.
Текст научной статьи Сопряженное влияние кортизола и тестостерона на адаптационную стратегию поведения в условиях «открытого поля»
Введение. Стрессогенные условия предполагают реактивное проявление комплекса поведенческих и вегетативных реакций, лежащих в основе адаптации к изменениям среды. Особенности проявления адаптации обусловлены конституциональными характеристиками организма и результатом эпигенетических процессов.
Напряжение эндокринной системы сопровождает сложный поведенческий сценарий, являясь и причиной, и маркером возникающих психоэмоциональных состояний.
Антагонизм кортизола и тестостерона основан на факте ингибирования анаболического действия тестостерона выделением кортизола в кровь и его воздействии на клетки-мишени через специфические клеточные рецепторы [1].
«Открытое поле» является инструментом, позволяющим в искусственных условиях воспроизвести тревожное эмоциональное состояние (спровоцированное подобное тревоге поведение), используя освещение, замкнутое
пространство и изоляцию животного от привычной обстановки содержания [2].
Цель исследования. Изучение сопряженного влияния кортизола и тестостерона на адаптацию животных в условиях «открытого поля» с учетом их конституциональных особенностей.
Материалы и методы. Исследование проводили на 20 взрослых крысах-самцах линии «Вистар», поведение которых изучали, фиксируя поведенческие акты и позы. Динамика поведенческой адаптации оценивалась ежедневно в утреннее время в течение 5 дней, экспериментальная сессия длилась 5 мин. Регистрация и обсчет данных проводились в соответствии с методикой И.Ю. Забродина [3] по формуле энтропии поведения:
Н=-Σ P I log ₂ P I ΣP ij log ₂ P ij Σ ijk log ₂ P ijk , где Н – показатель энтропии, Р I – вероятность появления i-го поведенческого акта; P ij – вероятность появления i-го поведенческого акта за
j-м состоянием; P ijk – вероятность следования трех поведенческих актов друг за другом.
Вероятность следования поведенческих актов «движение животного по замкнутому периметру поля», «обнюхивание», «вертикальная стойка», «груминг», «неподвижность», «движение на месте», «норковый рефлекс», «вертикальная стойка на стенку» составляла основу для создания индивидуального поведенческого профиля животного. В полученном массиве выделяли минимальное и максимальное значение энтропии поведения. Интервал от минимального показателя энтропии до максимального делили на три части. Животные, имевшие высокие значения энтропии, попадали в 1-ю группу (8 высокоэнтропийных животных), низкие значения показателя – в 3-ю (6 низкоэнтропийных животных), остальные – во 2-ю (6 животных), где средние значения показателя энтропии составляли 1,16133 (табл. 1).
Таблица 1
Table 1
Энтропия поведения крыс в открытом поле в течение экспериментальной серии Behavioral entropy in rats in the open field in experiments
По количеству актов дефекации и урина-ции оценивали эмоциональность животных. Интенсивность проявления тревожности, сопровождающейся вегетативным напряжением, выраженность которого имеет достоверные отличия у животных с разными показателями энтропии поведения, определяли по уровню кортизола и тестостерона в крови им-муноферментным методом на анализаторе Immulite (DPS Cirrus Inc., США) [4, 5].
Исследования проводились в соответствии с положениями Хельсинкской декларации об этических принципах медицинских исследований, утвержденной Всемирной медицинской ассоциацией в 1964 г.
Результаты. В сыворотке крови показана вариабельность концентраций кортизола и тестостерона у особей всей экспериментальной группы – уровень кортизола составил 67,33± ±33,32 нМ/л, тестостерона – 38,44±10,67 нМ/л (V=49 % и V=27 % соответственно). Выявлены коррелятивные отношения между содержанием кортизола и энтропией поведения как показателем меры неупорядоченности разнообразных форм проявления поведенческой адаптации в «открытом поле» (ОП) (r=-0,319). Безусловно, это подтверждает суждение о том, что энтропия поведения дает возможность определять копинг-стратегию животного в условиях обстановочной неопределенности.
|
Группа Group |
1-я группа, n=8 Group 1, n=8 |
2-я группа, n=6 Group 2, n=6 |
3-я группа, n=6 Group 3, n=6 |
|
Показатель энтропии (H) Entropy index (H) |
1,256±0,0709 |
1,16133±0,06426 |
0,85617±0,16369 |
|
Значение р при α=0,05 p (α=0.05) |
р<0,001 |
р<0,001 |
р<0,001 |
Таблица 2
Table 2
Связь концентрации свободного кортизола в крови с поведенческими актами у крыс
Correlation between free cortisol concentration in blood and rat behaviour
|
Поведенческий акт Behaviour act |
я s я О |
J « п 2 а СО р |
Я |
£ 5 s я с 2 я S и ф © |
е -2 © а щ а |
© я £ я я « « О й ? а |
|
Коэффициент корреляции Correlation coefficient |
r=-0,060* |
r=-0,036* |
r=-0,166* |
r=-0,099* |
r=-0,143* |
r=-0,581** |
Примечание. * – р<0,005, ** – р<0,001.
Note. * – p<0.005, ** – p<0.001.
Минуты эксперимента
Minutes of the experiment
0,25 0,216
Minutes of the experiment
А / A Б / B
Рис. 1. Динамика вероятности актов «неподвижность» (А) и «вертикальная стойка на стенку» (Б) в течение экспериментальных серий у высококортизоловых крыс
Fig. 1. Probability dynamics of “Immobility” (A) and “Upright posture on the wall” (B) in high-cortisol rats in the experiments
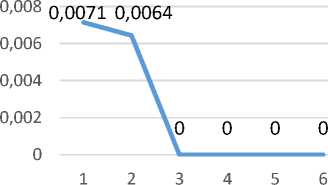
Поведенческий акт «вертикальная стойка на стенку» является своеобразным проявлением активного поисково-ориентировочного поведения в «открытом поле». Изучение динамики данного поведенческого акта показало, что его частота у высококортизоловых животных незначительно снижается, однако остается достаточной в течение всей экспериментальной серии (рис. 1 Б).
Нарастающее присутствие поведенче-
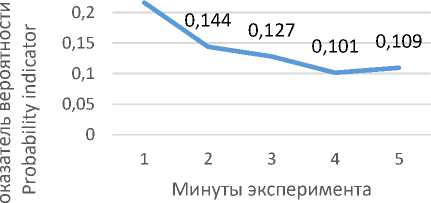
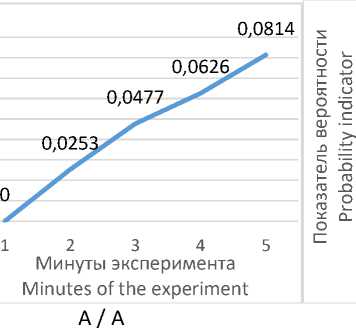
низкокортизоловых животных, что может служить своеобразным внешним маркером адаптации данной группы животных к условиям ОП (рис. 2 А). Вероятность проявления поведенческого акта «вертикальная стойка» резко возрастает в начале опыта (на вторую минуту) и далее снижается (рис. 2 Б), что показывает неустойчивый характер его включения в поведенческий адаптивный сценарий
ского акта «неподвижность» характерно для
низкокортизоловых крыс.
Либо животное активно осваивает пространство, проявляя поисковое поведение, либо оно эмоционально переживает стресс, сопровождаемый тревожностью и соответствующим напряжением гипоталамо-гипофизарной надпочечниковой оси (ГГН-контура). Насколько будет проявлена активация ГГН-контура в ответ на стрессорный фактор – характеристика, слабо соотносящаяся с конституциональными особенностями высшей нервной деятельности. Очевидно, активация ГГН-контура прово-
цирует комплекс адаптационных перестроек, в котором поведение и содержание гормонов являются не причиной, а следствием воспроизводимых эффекторных влияний ЦНС. Вместе с тем эмоциональное напряжение можно рассматривать не только через высокий уровень кортизола, но и через присутствие тех или иных сочетаний поведенческих актов в «открытом поле». Так, обнаружена взаимосвязь концентрации кортизола с отдельными поведенческими актами (табл. 2).
«Вертикальная стойка на стенку» – поведенческий акт, определяющий готовность животного сменить плоскость перемещения для оглядывания или преодоления препятствия «открытого поля», иными словами, это состояние готовности к смене условий вопреки стратегиям утаивания, замирания. Чем выше уровень кортизола, тем ниже показатель вероятности проявления акта «вертикальная стойка на стенку». Переход животного в иную плоскость (вниз – «норковый рефлекс» или вверх – «вертикальная стойка на стенку») является стратегией поисковой активности, характерной для низкокортизо-ловых животных. По мнению ряда авторов, по
горизонтальной и вертикальной двигательной активности можно оценить ориентировочно-исследовательскую реакцию крыс [8–10]. Этот вывод подтверждается обратной взаимозависимостью содержания кортизола и проявления поведенческого акта «норковый рефлекс» (табл. 2).
Немаловажной характеристикой, показывающей наличие взаимосвязи стратегии адаптации к «открытому полю» и содержания кортизола, является динамика вероятностного проявления того или иного поведенческого акта (рис. 1). Вы-сококортизоловые животные демонстрируют снижение вероятности проявления поведенческого акта «неподвижность» (рис. 1 А).
0,09
0,08
0,07
0,06
0,05
0,04
0,03
0,02
0,01
0,35
0,3
0,25
0,2
0,15
0,1
0,05
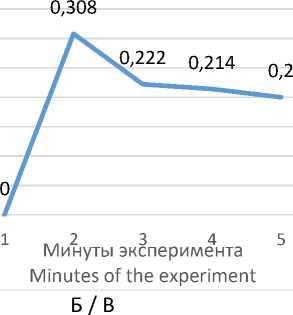
Рис. 2. Динамика вероятности актов «неподвижность» (А) и «вертикальная стойка» (Б) в течение экспериментальных серий у низкокортизоловых крыс
Fig. 2. Probability dynamics of “Immobility” (A) and “Upright posture” (B) in low-cortisol rats in experiments
Помимо взаимосвязи между содержанием кортизола и поведенческими актами, нами была выявлена корреляция уровней кортизола и тестостерона (r=-0,568), что подкрепляет
утверждение об антагонистическом характере их влияния на клеточные и субклеточные процессы. Вместе с тем взаимосвязь концентрации свободного кортизола и количества актов
уринации и дефекации может свидетельство- влиянии гормонов на работу желудочно-кишеч-вать о сложном, комплексном и нелинейном ного тракта и выделительной системы (табл. 3).
Таблица 3
Table 3
Связь концентрации свободного кортизола в крови с уровнем тестостерона в крови и количеством актов дефекации, уринации
Correlation between the concentration of free cortisol in blood and testosterone level in blood and defecation/urination rate
|
Фактор Factor |
Содержание тестостерона в крови крыс Testosterone level in rat blood |
Количество актов уринации Urination rate |
Количество актов дефекации Defecation rate |
|
Коэффициент корреляции Correlation coefficient |
r=-0,568** |
r=0,219 |
r=0,143 |
Примечание . ** – р<0,001.
Note. ** – p<0.001
В ходе эксперимента была также показана вотных и частоты проявления ряда поведенче-корреляция уровня тестостерона в крови жи- ских актов (табл. 4).
Таблица 4
Table 4
Связь концентрации тестостерона с вероятностностью проявления поведенческих актов в ходе эксперимента
Между уровнем тестостерона и актом «обнюхивание» существует положительная корреляционная связь. Концепция поведенче-
ской детерминации объясняет это необходимостью перемежать поисковую и перцептивную активность. Низкокортизоловые живот-
Correlation between testosterone level and behavioral probability in experiments
|
Поведенческий акт Behaviour act |
я s я О |
й « g СО р |
Я м Я |
я «8й |
е -2 Я © а |
* к 2 j И и я р W о "88 &« а° ® s^ |
|
|
Коэффициент корреляции Correlation coefficient |
r=0,551*** |
r=0,483** |
r=-0,117 |
r=0,118 |
r=0,748* |
r=-0,394 |
r=0,649*** |
Примечание . * – р<0,005, ** – р<0,001, *** – р<0,001.
Note. * – p<0.005, ** – p<0.001, *** – p<0.001.
ные предпочитают поведенческие формы активного сбора информации извне активному и хаотичному исследованию ОП, сохраняют поисково-ориентировочную стратегию поведения, что указывает на тенденцию перехода к высокоэнтропийному поведению в ОП. Сложный характер взаимосвязей уровня тестостерона с другими поведенческими актами также является следствием того, что стрессовое состояние при наличии высокой концентрации тестостерона предполагает стратегию поисковой активности с упором на перцепцию: поведение носит характер поиска новых возможностей, условий для преодоления создавшейся стрессогенной ситуации.
Между содержанием тестостерона и энтропией имеется слабая связь (r=-0,161), поскольку влияние гормона на структуру поведения животного опосредовано комплексом факторов и, скорее всего, не определяет напрямую меру упорядоченности. Тестостерон не только обусловливает поведенческую активность, он направляет ее, исходя из ярко выраженной стратегии исследования обстановки животным, преодоления им средовых обстоятельств. Кроме того, его связь с процес-
сами тканевого развития, в т.ч. нервной ткани, способствует последующему установлению межнейрональных связей, оказывая комплексное воздействие на поведение животного [11].
Противоположные взаимозависимости концентрации тестостерона и частоты урина-ции и дефекации (табл. 5) свидетельствуют о наличии различных комплексных механизмов регуляции указанных процессов. Колоректальные пропульсивные сокращения [12], активируемые при общем подъеме тонуса желудочно-кишечного тракта, вызывают повышенное выделение болюсов, поскольку животное, проявляя двигательную активность в ОП, нуждается в поступлении энергии. В акте дефекации при стрессе задействован гиппокамп, а эмоциональное напряжение и его связь с уринацией объясняются активацией миндалины, что проясняет значительные отличия характера взаимосвязи этих показателей и уровня тестостерона в крови [13, 14]. Гиппокамп влияет на актуализацию воспоминаний при неоднократном предъявлении стимула или информации, связанной с необходимостью ориентации в пространстве [15].
Таблица 5
Table 5
Связь содержания тестостерона в крови с вегетативными показателями Correlation between testosterone level in blood and vegetative indicators
|
Фактор Factor |
Показатель уринации Urination rate |
Показатель дефекации Defecation rate |
|
Коэффициент корреляции Correlation coefficient |
r=-0,756* |
r=0,174 |
Примечание. * – р<0,001.
Note. * – p<0.001
В исследовании показана прямая связь уровня тестостерона с количеством актов дефекации и обратная – с количеством актов уринации. Это дополняет известное суждение о том, что оба показателя являются следствием эмоционального напряжения, которое имеет разную направленность.
Обсуждение. Исходя из концепции поведенческой детерминации адаптацию необходимо рассматривать как комплексный механизм. Животные, демонстрирующие разный характер копинг-стратегий или (и) отличающиеся по показателям ГГН-системы, могут иметь определенные поведенческие и вегетативные маркеры - от элементарных поведенческих актов до содержания гормонов в крови. Кортизол и тестостерон находятся в реципрокных отношениях, и уровень их содержания в совокупности с теми или иными особенностями поведения или вегетативных реакций свидетельствует о наличии индивидуальной стратегии адаптации к стрессовой обстановке, предъявляемой в эксперименте. Кортизол сопровождает и моделирует депрессивно-подобное поведение, а его эффект, по мнению ряда авторов [6], зависит от типа испытуемых. Действительно, клинические наблюдения показывают, что у некоторых групп животных в ситуации развития артериальной гипертензии выявляется высокое содержание кортизола на фоне активации симпатоадреналовой системы, а многократное повторение стрессорной обстановки для субъекта (как в серии описываемых опытов в «открытом поле») сдвигает ГГН-систему в аллостатическое состояние [7], когда стабилизация возможна через существенную перестройку работы систем организма. Функциональный антагонизм и возможность вариативного изменения рецепторного связывания кортикостероидов и половых стероидов в онтогенезе [18] позволяют рассматривать соотношение «кортизол/тестостерон» как маркер эустресса [19, 20].
Различия в динамике вероятности проявления тех или иных поведенческих актов в ходе адаптации крыс разных типологических групп можно интерпретировать с помощью привлечения данных о содержании кортизола или тестостерона в крови. Их концентрация имеет слабовыраженную зависимость от энтропии поведения. Уровень тех или иных гормонов, а также оказанный ими на адаптацию в условиях стресса эффект опосредованы влиянием генетических и эпигенетических факторов, предполагающих формирование индивидуального поведенческого сценария.
Катаболический и анаболический эффекты гормонов (кортизола и тестостерона соответственно) взаимообусловлены геномной и негеномной активацией сателлитов, а также усилением или понижением чувствительности рецептора андрогена и его потенциальной ролью в привлечении белков-коактиваторов и повышении транскрипционной активности [8]. В ряде случаев физиологический эффект глюкокортикоидов опосредован однонуклеотидным полиморфизмом глюкокортикоидных рецепторов, которые изменяют мотивы их связывания или взаимодействующие с ними факторы [17]. Дифференцированное влияние кортикостероидов определяет характер двигательной активности: либо поисковая стратегия поведения, либо стратегия замирания, стагнации с преобладанием «бедного» поведенческого репертуара.
Иными словами, характер стратегии поведения определяется как конституциональным, так и эпигенетическим сценарием формирования адаптационного ответа. Таким образом, концентрации и соотношение тестостерона и кортизола в сочетании с поведенческими маркерами адаптации к условиям ОП могут служить основой для объективной интерпретации поведения животного.
Заключение. Пребывание животного в незнакомой и стрессогенной обстановке способствует проявлению активации ГГН-контура. Эмоциональное напряжение может проявляться не только через высокий уровень кортизола, но и через присутствие тех или иных сочетаний поведенческих актов в «открытом поле». Концентрация кортизола, имея отрицательную взаимосвязь с уровнем тестостерона, демонстрирует сопряжение с отдельными поведенческими актами. В условиях неопределенности (ОП) у крыс проявляются две основные стратегии. Вы-сококортизоловые животные обнаруживают готовность сменить плоскость перемещения с горизонтальной на вертикальную, демонстрируя маловариативный арсенал поведенческих актов или предпочтение поведенческих форм активного сбора информации хаотичному передвижению в ОП. Низкокортизоловые животные предпочитают поведенческие формы активного сбора информации извне активному и хаотичному исследованию ОП, сохраняют поисковоориентировочную стратегию, что является тенденцией перехода к высокоэнтропийному поведению в ОП. Функциональный антагонизм и возможность вариативного изменения рецепторного связывания кортикостероидов и половых стероидов в онтогенезе создает возможность рассматривать соотношение «корти-зол/тестостерон» как маркер эустресса.
Список литературы Сопряженное влияние кортизола и тестостерона на адаптационную стратегию поведения в условиях «открытого поля»
- Андреева А.В., Анциферов М.Б. Современные возможности медикаментозного контроля болезни Иценко-Кушинга. Эндокринология. 2021; 3: 66-74.
- Каде А.Х., Кравченко С.В., Трофименко А.И. Современные методы оценки уровня тревожности грызунов в поведенческих тестах, основанных на моделях без предварительного обусловливания. Кубанский научный медицинский вестник. 2018; 25 (6): 171-176. DOI: 10.25207/1608-6228-201825-6-171-176.
- Забродин И.Ю., Петров Е.С., Вартанян Г.А. Анализ свободного поведения животных на основе его вероятностных характеристик. Журнал высшей нервной деятельности. 1983; 1: 71-80.
- Филатова О.В., Червова И.В., Киселев В.Д., Пискунова Е.Р. Эндотелий-зависимая потоковая реактивность брюшной аорты у крыс с различными типами поведения в открытом поле (кластерный анализ). Известия Алтайского государственного университета. 2005; 3: 111-112.
- Johnson S., Fournier N., Kalynchuk L. Effect of different doses of corticosterone on depression-like behavior and HPA axis responses to a novel stressor. Bchav. Brain Res. 2006; 168 (2): 280-288.
- Reuter М., Netter Р., Rogausch А., Sander Р., Kaltschmidt М., Dorr А., Hennig J. The role of cortisol suppression on craving for and satisfaction from nicotine in high and low impulsive subjects. Human Psychohharmacol. 2002; 17 (5): 213-224.
- Козлов А.И., Козлова М.А. Кортизол как маркер стресса. Физиология человека. 2014; 2: 123-136.
- William J. Kraemer, Wesley C. Hymer, Bradley C. Nindl, Maren S. Fragala. Growth Hormone(s), Testosterone, Insulin-Like Growth Factors, and Cortisol: Roles and Integration for Cellular Development and Growth With Exercise. Front. Endocrinol. 2020; 11: 33.
- Шаляпина В.Г., Ракицкая В.В., Петрова Е.И. Роль кортикотопин-рилизинг гормона в нарушениях поведения после неизбегаемого стресса у активных и пассивных крыс. Журнал высшей нервной деятельности им. И.П. Павлова. 2005; 2: 241-246.
- Родина В.И., Крупина Н.А., Крыжановский Г.Н. Многопараметровый метод комплексной оценки тревожно-фобических состояний у крыс. Журнал высшей нервной деятельности им. И.П. Павлова. 1993; 5: 1006-1017.
- Прокопенко Е.С., Надей О.В., Агалакова Н.И. Исследование экспрессии генов NMDA И AMPA рецепторов в вентральном гиппокампе крыс разных возрастных групп при депрессивно-подобном состоянии. Сборник тезисов 24 съезда Физиологического общества им. И.П. Павлова. 2023: 44.
- Mayuko N., Ishimizu Y, Saitoh S., Okada H., Fukuda H. The defecation reflex in rats: fundamental properties and the reflex center. Auton Neurosci. 2004; 111 (1): 48-56.
- Иванов Д.Г., Подковкин В.Г. Взаимосвязь уровня метаболизма коллагена и поведения крыс в тесте «открытое поле». Успехи современного естествознания. 2010; 5: 16-20.
- Arutjunyan А., Milyutina Y., Shcherbitskaia А., Kerkeshko G., Zalozniaia I. Epigenetic Mechanisms Involved in the Effects of Maternal Hyperhomocysteinemia on the Functional State of Placenta and Nervous System Plasticity in the Offspring. Biochemistry. 2023; 88 (4): 435-456.
- Anagnostou E., Skarlatou V., Mergner T., Anastasopoulos D. Idiothetic signal processing and spatial orientation in patients with unilateral hippocampal sclerosis. Neurophysiol. 2018; 120 (3): 1256-1263.
- Kraemer W., Ratamess N., Hymer W., Nindl B., Fragala M. Growth Hormone(s), Testosterone, InsulinLike Growth Factors, and Cortisol: Roles and Integration for Cellular Development and Growth With Exercise. Endocrinol. 2020; 11: 33.
- Hu W., Jiang C., Kim M., Wenjian W., Zhu K., Guan D., Lv W. Individual-specific functional epigenomics reveals genetic determinants of adverse metabolic effects of glucocorticoids. Cell Metab. 2021; 33 (8): 1592-1609.
- Ордян Н.Э. Гормональные механизмы фенотипической модификации стрессорной реактивности в онтогенезе крыс: дис.... д-ра биол. наук: 03.00.13. Санкт-Петербург; 2003. 262.
- Ветлугина Т.П., Никитина В.Б., Лобачева О.А. Уровень кортизола и тестостерона у больных алкоголизмом при синдроме отмены. Сибирский вестник психиатрии и наркологии. 2017; 3 (96): 5-10.
- Шпак А.Н., Курочкина Е.А. Динамика уровня гормонов тестостерона и кортизола в сыворотке крови крыс при длительной нагрузке разной интенсивности. Международный вестник ветеринарии. 2012; 2: 54-57.
