Сопряженность морфогенеза генеративных органов в цветках растений остролодочника Oxytropis DC. (Fabaceae Lindl.)
")
Автор: Круглова Анна Евгеньевна, Маслова Наталья Владимировна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Природопользование
Статья в выпуске: 5-3 т.13, 2011 года.
Бесплатный доступ
На основании анализа впервые полученных цито-гистологических данных по развитию пыльника и семяпочки интродуцированных растений Oxytropis DC. (семейство бобовые) рассмотрена сопряженность морфогенеза этих генеративных органов в цветке. Полученные данные могут быть использованы при проведении экспериментальных исследований интродуцированных растений Oxytropis DC.с целью повышения их реальной семенной продуктивности.
Редкие растения, интродукция, пыльник, семяпочка, морфогенез
Короткий адрес: https://sciup.org/148200475
IDR: 148200475 | УДК: 581.33
The conjugation of morphogenesis of generative organs at the flowers of plants of oxytropis DC. (Fabaceae Lindl.)
Based on the analysis of the first obtained cyto-histological data on the development of anther and ovule of introduced plants Oxytropis DC. ( Fabaceae ) the conjugation of morphogenesis of this generative organs at the flower has considered. The data obtained can be used for conducting experimental studies of introduced plants Oxytropis DC. to improve their actual seed production.
Текст научной статьи Сопряженность морфогенеза генеративных органов в цветках растений остролодочника Oxytropis DC. (Fabaceae Lindl.)
Эндемичные и реликтовые виды рода остролодочник Oxytropis DC. (семейство бобовые Fabaceae Lindl.) относятся к группе редких и исчезающих, занесенных в Красные книги многих регионов России, в том числе Республики Башкортостан [4].
Один из эффективных приемов сохранения, размножения и увеличения численности особей редких и исчезающих видов растений – их интродукция в питомники ботанических садов [1, 14 и др.] с последующей реинтродукцией в естественные местообитания для восстановления природных популяций [10]. Для успешной реинтродукции требуется значительное количество качественных семян интродуцированных растений. В то же время хорошо известно, что у интродуцированных растений в связи со сменой местообитаний могут быть снижена семенная продуктивность (по [9]).
Проблема взаимосвязей различных процессов в онтогенезе растений – одна из интереснейших в ботанике. Помимо большого значения в создании общей теории онтогенеза цветковых растений, решение этой проблемы имеет чисто практический выход – управление качеством и количеством образуемых плодов, раннее и точное их прогнозирование.
В конкретном приложении к интродуцированным растениям рода остролодочник это означает, как важно по морфологическим признакам установить, какие именно морфогенетические процессы проходят в цветке растений на конкретном этапе развития с тем, чтобы определить характер и время возникновения возможных нарушений, приводящих к снижению потенциальной семенной продуктивности, а также определить точные сроки проведения экспериментальных работ по снижению таких нарушений, а значит, увеличению реальной семенной продуктивности (количества качественных семян).
Цель исследования состояла в оценке сопряженности морфогенетических процессов, проходящих в цветке растений рода остролодочник, а именно – сопряженности морфогенеза пыльника и семяпочки как генеративных органов. Данные детальных цито-гистологических исследований морфогенеза пыльника и семяпочки остролодочников получены нами ранее [5, 6, 7]. Установлено при этом, что морфогенез генеративных органов у остролодочников проходит типично для представителей семейства Fabaceae Lindl. [12, 16].
МАТЕРИАЛ И МЕТОДЫ
Объект исследования - редкие и находящиеся под угрозой исчезновения виды рода остролодочник из флоры Южного Урала: реликт о. башкирский O. baschkirensis Knjasev , эндемик о. Гмелина O. gmelinii Fisch. ex Boriss, эндемик о. уральский O. uralensis (Lam.) DC., интродуцированные в питомник редких растений Института биологии УНЦ РАН (на территории Ботанического сада-института УНЦ РАН) в течение 1997-2000 гг.
Растения интродуцированы семенами, собранными в естественных местообитаниях на Южном Урале (коллекторы А.Х. Галеева и А.А. Мулдашев, которым авторы выражают искреннюю благодарность).
Фенологические наблюдения за развитием растений 2-го года жизни, вступивших в генеративный период развития, вели согласно [2] в течение вегетационных сезонов 2006-2010 гг. Для исследования морфогенеза генеративных органов (пыльника и семяпочки) использовали общепринятые цито-гистологические методы [11]. Постоянные препараты просматривали при различном увеличении объектива с применением светового микроскопа Axio Imager 1 (Carl Zeiss, Jena) с программным управлением и вмонтированной цифровой камерой, а также при помощи цифрового микроскопа проходящего света Микровизор mVizo-103 (ООО «ЛОМО ФОТОНИКА», Санкт-Петербург).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Рассмотрим подробнее те фенологические фазы развития изучаемых растений, которые непосредственно связаны с формированием и развитием генеративных структур - пыльника и семяпочки. Приведем данные, полученные для остролодочника башкирского.
В начале фенологической фазы бутонизации (рис. 1а) , когда длина соцветия составляет в среднем 2,2 см, длина бутона - 1,5 мм, в цветках происходит закладка тычиночных бугорков и бугорков завязи, представленных меристематической тканью (рис. 1б, в). Далее, с увеличением размера бугорков тычинки и завязи, в них путем множественных митотических делений меристематических клеток происходит постепенное формирование пыльника и семяпочки. Так, в пыльнике формируются клетки археспория, которые дают начало спорогенной ткани и тканям
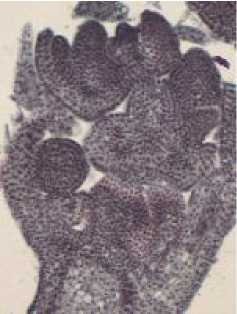

стенки гнезда. В фенофазу бутонизации (рис. 2а), при таких морфологических показателях, как средняя длина соцветия 2,7 см, средняя длина бутона - 2,2 мм, в цветке отмечается сформированный пыльник (рис. 2б), т.е. пыльник, стенка гнезда которого характеризуется наличием сформированных тканей (экзотеций, эндотеций, средний слой, тапетум). Развивающиеся спорогенные клетки при этом находятся в стадии микроспороцита.
Развитие семяпочек тесно связано с развитием завязи. В основании завязи на ее вентральной стороне в виде меристематических бугорков закладываются 12-15 семяпочек.
Семяпочка развивается медленнее пыльника, и на этой стадии развития цветка представлена еще не дифференцированными, но уже хорошо развитыми меристематическими бугорками (рис. 2в).
а
б
в
Рис. 1. а) остролодочник башкирский в начале фенофазы бутонизации, макросъёмка, х1.8; б-в) цветок остролодочника башкирского в начале фенофазы бутонизации, постоянные препараты, продольный (б, х8) и поперечный (в, х10) срезы.
а б в
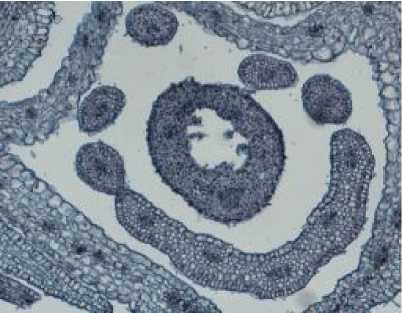
Рис. 2 . а) остролодочник башкирский в фенофазе бутонизации, макросъёмка, х1.2; б-в) цветок остролодочника башкирского в фенофазу бутонизации, постоянные препараты, продольный (б, х8) и поперечный (в, х10) срезы.
В ходе дальнейшего развития цветка в фенофазу бутонизации в пыльниках проходит мейоз микроспороцитов, формируются диады и тетрады микроспор. В процессе активного роста микроспоры одной тетрады обособляются друг от друга, сильно увеличиваясь в размерах. В стенке гнезда пыльника происходят дегенерация клеток тапетума и среднего слоя, формирование фиброзных утолщений в оболочках клеток эндотеция.
Большие изменения претерпевает семяпочка. Дифференцируется центральная ее часть – нуцеллус, который постепенно становится массивным, хорошо развитым. Начинают формироваться интегументы – первоначально внутренний, затем наружный. Формирование интегументов совпадает с изгибом семяпочки за счет неравномерного роста клеток нуцеллуса и клеток формирующихся интегументов. В субэпидермальном слое апикальной части нуцеллуса дифференцируется клетка археспория, дающая начало макроспороциту.
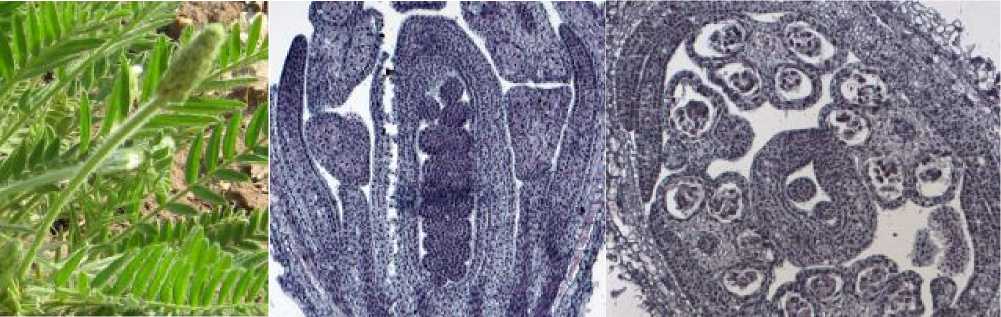
В конце фенофазы бутонизации (рис. 3а), при длине соцветия в среднем 3,1 см, длине бутона – 2,5 мм, пыльник находится в стадии микроспоры, семяпочка – в стадии макроспороцита (рис. 3б, в).
а б в
Рис. 3 . а) остролодочник башкирский в конце фенофазы бутонизации, макросъёмка, х0.7; б-в) цветок остролодочника башкирского в конце фенофазы бутонизации, постоянные препараты, продольный (б, х20) и поперечный (в, х8) срезы.
Цветок развивается далее. Из микроспор путем митотических делений формируются двуклеточные пыльцевые зерна. Зрелый пыльник, готовый к вскрыванию, содержит двуклеточные пыльцевые зерна. Из тканей стенки гнезда ясно выражены только узкоспециализированные ткани – экзотеций, предназначенный для защиты пыльника, и эндотеций, выполняющий функцию вскрывания зрелого пыльника.
Семяпочка также постепенно развивается. Макроспороцит дает начало линейной тетрады макроспор, при этом халазальная макроспора развивается в зародышевый мешок. Эта клетка претерпевает три последовательных митоза, приводящих к формированию 8-ядерного 7клеточного зародышевого мешка Polygonum-типа. В фенофазу начала цветения в цветках отмена сформированная семяпочка, содержащая сформированный зародышевый мешок.
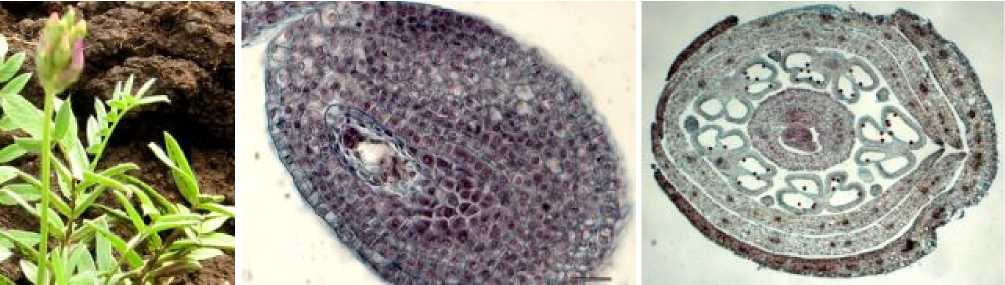
В фенофазу начала цветения (рис. 4а), при средней длине соцветия 4,5 см, средней длине цветка – 3,8 мм, в цветках отмечаются зрелые пыльники и сформированные семяпочки (рис. 4б, в).
а
б
в
Рис. 4. а) остролодочник башкирский в фенофазу начала цветения, макросъёмка, х1.2; б-в) цветок остролодочника башкирского в фенофазу начала цветения, постоянные препараты, продольный (б, х30) и поперечный (в, х6) срезы.
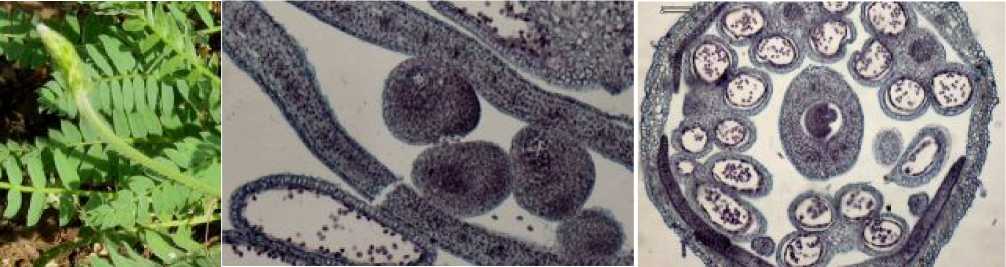
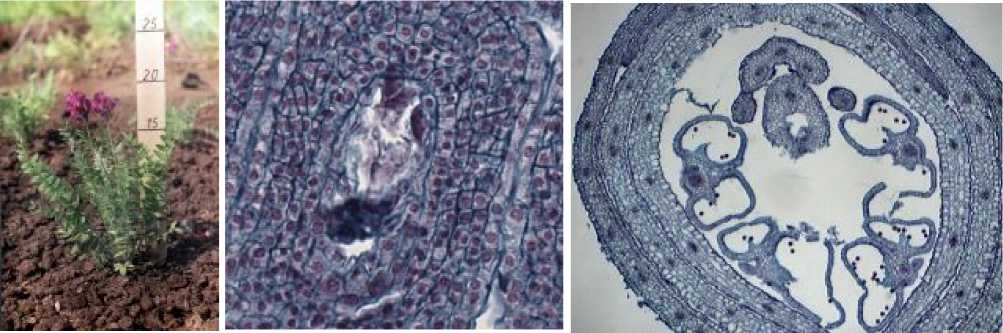
Зрелые семяпочки, содержащие зрелые зародышевые мешки, отмечены в цветках в фенофазу массового цветения (рис. 5а,б), когда средняя длина соцветия достигает 6,2 см, а средняя длина цветка – 4,5 мм. Зрелые пыльники при этом уже вскрыты внутри закрытого цветка (рис. 5в).
Таким образом, семяпочка развивается медленнее по сравнению с пыльником. По-видимому, в данном случае сказывается более сложная структура этого генеративного органа. Кроме того, на наш взгляд, в такой асинхронности большую, если не основную, роль играет перекрестный способ опыления остролодочников.
Действительно, в отличие, например, от самоопыляемых растений, для которых характерна синхронность в развитии генеративных органов и гаметофитов [15], у перекрёстно опыляемых растений нет необходимости в такой синхронизации.
В целом, согласно результатам наблюдений, формирование генеративных органов приходится на фенофазу бутонизации. Зрелые пыльники в цветках растений всех трёх изучаемых видов отмечены в фенофазу начала цветения, зрелые семяпочки – в фенофазу массового цветения.
Важно по внешним признакам цветка определить, на каком именно этапе морфогенеза находятся пыльник и семяпочка растения. Предложено, например, определять этап развития пыльника лилии – по длине пыльника [13], стадию развития гаметофитов злаков – по расположению колосковых чешуй [8].
Мы предлагаем оценивать развитие семяпочки и пыльника остролодочника по соотношению длины венчика и чашечки. Так, закладка тычиночных бугорков и бугорков завязи происходит в плотном бутоне, при этом венчик полностью скрыт в чашечке. В цветке, венчик которого на треть выступает над чашечкой, находятся сформированный пыльник и недифференцированные, но уже хорошо развитые меристематические бугорки семяпочки. Когда венчик по длине значительно превосходит чашечку, в цветках отмечаются зрелые пыльники и сформированные семяпочки.
Этот способ, предложенный Т.П. Белковской [3] при изучении развития цветка представителя семейства бобовых астрагала уральского, на наш взгляд, достаточно удобен и, кроме того, наглядно демонстрирует корреляцию в развитии генеративных структур растений.
Таким образом, в пределах одного цветка изученных растений рода остролодочник наблюдается определенная сопряженность в морфогенезе генеративных структур (пыльника и семяпочки), обусловленная перекрестным способом опыления цветков.
а б в
Рис. 5 . а) остролодочник башкирский в фенофазу массового цветения, макросъёмка, х0.7; б-в) цветок остролодочника башкирского в фенофазу массового цветения, постоянные препараты, продольный (б, х40) и поперечный (в, х8) срезы.
Исследование поддержано программой «Ведущие научные школы РФ» (грант № НШ 7637.2010.4, лидер Школы – член-корр. РАН Т.Б. Батыгина, БИН РАН, Санкт-Петербург).
Список литературы Сопряженность морфогенеза генеративных органов в цветках растений остролодочника Oxytropis DC. (Fabaceae Lindl.)
- Абрамова Л.М., Маслова Н.В., Каримова О.А. Интродукция редких видов как способ сохранения биоразнообразия (на примере Республики Башкортостан)//Бюлл. Главного ботанического сада. 2004. Вып. 188. С. 110-118.
- Бейдеман И.Н. Методика изучения фенологии растений и растительных сообществ. Новосибирск: Наука, 1974. 155 с.
- Белковская Т.П. К антэкологии некоторых реликтовых и эндемичных видов астрагалов Кунгурской лесостепи//Экология опыления растений: Межвуз. сб. научн. трудов. Пермь, 1984. С. 34-49.
- Красная книга Республики Башкортостан. Т. 1. Редкие и исчезающие виды высших сосудистых растений. Уфа: Китап, 2001. 280 с.
- Круглова А.Е. Эмбриология редкого вида Южного Урала остролодочника сходного: морфогенез пыльника//Вестник Оренбургского гос. ун-та. 2009. № 6 (100). С. 172-173.
- Круглова А.Е., Катасонова А.А., Маслова Н.В., Круглова Н.Н. Эмбриология редкого вида Южного Урала остролодочника сходного: морфогенез семяпочки//Известия Самарского НЦ РАН. 2010. Т. 12 (33). № 1. С. 727-729.
- Круглова А.Е. Оценка качества пыльцевых зерен в зрелых пыльниках остролодочника сходного в условиях интродукции//Вестник Удмуртского ун-та. Серия Биология. Науки о Земле. 2011. Вып. 1. С. 67-74.
- Круглова Н.Н. Визуальная оценка стадий развития гаметофитов злаков//Ш Всеросс. конф. по ботан. ресурсоведению: Труды. СПб., БИН РАН, 1996. С. 185-186.
- Левина Р.Е. Полноценность семян и интродукция//Биологические основы семеноведения и семеноводства интродуцентов. Новосибирск: Наука, 1974. С. 7-8.
- Мулдашев А.А., Маслова Н.В., Галеева А.Х. Некоторые итоги изучения редких видов рода остролодочник (Oxytropis DC. -Fabaceae) в Республике Башкортостан и проблемы их охраны//II междунар. научно-практич. конф. «Природное наследие России в 21 веке»: Материалы. Уфа, 2008. С. 297-301.
- Паушева З.П. Практикум по цитологии растений. М.: Колос, 1988. 170 с.
- Поддубная-Арнольди В.А. Характеристика семейств покрытосеменных растений по цитоэмбриологическим признакам. М.: Наука, 1982. 351 с.
- Резникова С.А. Цитология и физиология развивающегося пыльника. М.: Наука, 1984. 270 с.
- Стратегия ботанических садов по охране растений. М., 1993. 62 с.
- Терёхин Э.С. Семя и семенное размножение. СПб.: Мир и семья, 1996. 377 с.
- Чубирко М.М. Семейство Fabaceae/М.М. Чубирко, Л.Н. Кострикова//Сравнительная эмбриология цветковых растений. Т. 3. Brunelliaceae-Tremandraceae/отв. ред. М.С. Яковлев. Л.: Наука, 1985. С. 67-77.
