Состав и строение древостоев сфагновых ельников в верховьях реки Печора (Печоро-Илычский заповедник)
")
Автор: Алейников А.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Лесные ресурсы
Статья в выпуске: 1-4 т.13, 2011 года.
Бесплатный доступ
В статье проанализирован состав и строение древесной синузии среднетаежных сфагновых лесов Урала в сообществах двух типов леса: ельнике хвощево-сфагновом и чернично-долгомошно-сфагновом. Дана характеристика возрастной и онтогенетической структур ценопопуляций 4 видов деревьев. Показано, что в исследованных сообществах наиболее устойчивы и полночленны ценопопуляции ели.
Еловые леса, ценопопуляции, онтогенетические спектры
Короткий адрес: https://sciup.org/148199780
IDR: 148199780 | УДК: 630*182+630*228.81(1-924.93)
Compound and structure of sphagnum fir forest stands at Pechora riverheads (Pechoro-Ilychsky reserve)
In article compound and structure of forests synusium of middle-taiga sphagnum forests of Urals in communities of two forests types are analysed: a fir grove horsetail-sphagnum and bilberry-sphagnum. The age characteristic and ontoge-netic structures of 4 kinds cenopopulations of trees are given. It is shown that in the investigated communities are more steady and stable cenopopulations of fir-trees.
Текст научной статьи Состав и строение древостоев сфагновых ельников в верховьях реки Печора (Печоро-Илычский заповедник)
В настоящее время важное научное значение приобретает изучение структуры и состава древесной синузии спонтанно развивающихся лесов. Эти исследования необходимы для выяснения природных закономерностей функционирования лесных экосистем. Наибольшая сложность заключается в выборе района исследований, давно не испытавшего антропогенных воздействий. В результате предварительных исследований [5, 6] в качестве модельного объекта выбран бассейн р. Большая Порожняя (правый приток р. Печора), расположенный в Печоро-Илычском заповеднике на юго-востоке республики Коми (средняя подзона тайги). Детальные маршрутные исследования [3, 4] и дешифрирование космоснимков высокого и среднего разрешений показало, что в нижнем течении выбранного модельного бассейна доминируют сфагновые ельники, приуроченные к нижним частям дренированных склонов. Наиболее существенные отличия сфагновых лесов: ярко выраженное доминирование в напочвенном покрове межкроновых и подкроновых пространств сфагновых мхов (Sphagnum fallax, S. magellanicum, S. girgensohnii). Наиболее часто встречаются ельники двух типов леса: хвощево-сфагновые и чер-нично-долгомошно-сфагновые, отличающиеся условиями увлажнения и дренированности территории.
Цель работы: выявить особенности состава и строения древесной синузии сфагновых ельников.
В 2009 г. на правом берегу (восточная экспозиция) р. Б. Порожняя в ельнике чернично-долгомошно-сфагновом и ельнике хвощево-сфаг-новом заложены 2 постоянные пробные площади. Характеристика экотопических условий представлена в табл. 1. На пробных площадях проведен сплошной перечет всех деревьев с D 1.3m >2см. У каждого дерева определены онтогенетическое состояние, календарный возраст, жизненность, высота и диаметр ствола, размеры кроны [1, 2]. При кернении деревьев отмечено наличие стволовой гнили. С помощью ультразвукового дальномера Vertex III проведено картирование деревьев.
Таблица 1. Характеристика экотопических условий пробных площадей
|
№ ПП |
Тип леса |
Площадь ПП, га |
Высота над у.м. |
Уклон, гр. |
Преобладающая почва |
|
5 |
ельник хвощево-сфагновый |
0,25 |
270 |
2,2 |
торфяно-глеезем |
|
6 |
ельник чернично-долгомошно-сфагновый |
0,25 |
283 |
2,1 |
ржавозем торфянистоглеевый оподзоленный |
В сообществах обоих типов леса древесный ярус сложный по составу и состоит из ели сибирской ( Picea obovata ), пихты сибирской ( Abies si-birica ), кедра сибирского ( Pinus sibirica ) и березы
пушистой (Betula pubescence). Формула древостоя ельника хвощево-сфагнового, рассчитанная по сумме площадей сечения – 6,4Е 2,4Б 1,0 Пх 0,2К. Таксационная характеристика приведена в табл. 1. По числу деревьев содоминируют два вида: ель и пихта, значительно меньше березы и кедра. Однако по сумме площадей сечения содоминант ели – береза, а не пихта (табл. 1). Такое несоответствие объясняется тем, что на пробной площади много тонких стволов пихты и значительно меньше крупных стволов березы. Максимальный возраст ели – 260 лет, пихты – 125, кедра – более 400 (бо- лее точно определить не удалось из-за стволовой гнили). В качестве примера на рис. 1 показано пространственное размещение деревьев на этой пробной площади.
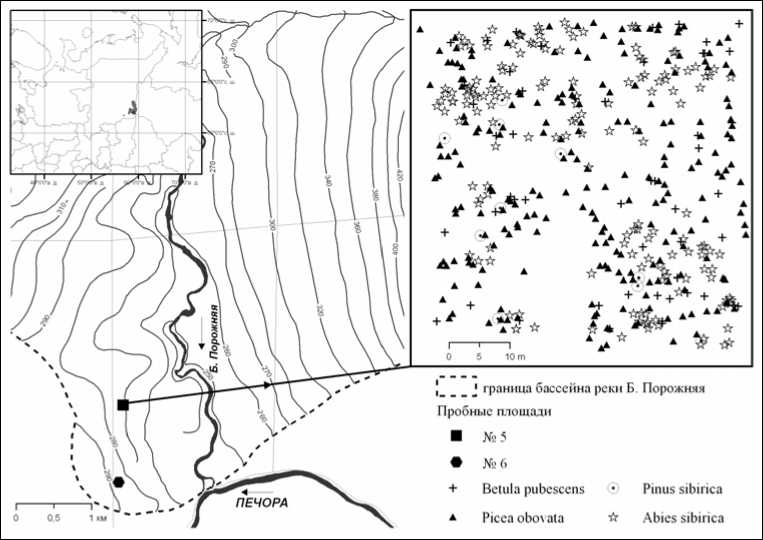
Рис. 1. Район исследования и пространственная структура древесной синузии ельника хвощево-сфагнового (ПП № 5)
В ельнике чернично-долгомошно-сфагно-вом абсолютное число живых деревьев больше, чем в ельнике хвощево-сфагновом, но сумма площадей сечения примерно на 20% меньше. Максимальный возраст ели и пихты здесь значительнее выше, чем в ельнике хвощево-сфагновом, что может свидетельствовать о более благоприятных условиях обитания этих видов. На обеих площадях по числу деревьев содоминируют ель и пихта, значительно меньше число деревьев березы и минимальное – кедра. По сумме площадей поперечных сечений содоминируют ель и береза, однако среди сопутствующих видов на 5 ПП больше березы, а на 6 ПП – кедра (табл. 2).
Таблица 2. Краткая таксационная характеристика пробных площадей
|
Тип леса (№ПП) |
Состав |
Вид* |
Число деревьев, экз./га |
Сумма площадей сечения, м2/га |
Максимальный возраст, лет |
|
|
всего |
обследован-ных |
|||||
|
ельник хвоще-во-сфагновый (5) |
6,4Е 2,4Б 1,0Пх 0,2К |
Е |
892 |
223 |
23,85 |
216 |
|
П |
644 |
161 |
3,69 |
125 |
||
|
К |
40 |
10 |
0,72 |
|||
|
Б |
216 |
54 |
9,04 |
|||
|
итого |
1792 |
448 |
37,29 |
|||
|
ельник чер- нично- долгомошно-сфагновый (6) |
5,4Е 1,9Б 1,6К 1,0 Пх |
Е |
968 |
242 |
14,9 |
303 |
|
П |
620 |
155 |
2,9 |
181 |
||
|
К |
96 |
24 |
4,5 |
|||
|
Б |
148 |
37 |
5,2 |
|||
|
итого |
1832 |
458 |
27,6 |
|||
*Примечание : Е – ель, П – пихта, К – кедр, Б – береза
Для ели и пихты, видов, преобладающих по числу стволов, характерна большая дифференциация по диаметру. Распределение стволов по ступеням толщины показано на рис. 2. Как видно, оба вида сохраняют одни и те же тенденции в разных сообществах. Ель более равномерно распределена в обоих сообществах. Преобладание пихты первых ступеней толщины связано с
Известия Самарского научного центра Российской академии наук, т. 13, №1(4), 2011
укоренением нижних ветвей: практически весь подрост в сообществах обоих типах вегетативного происхождения и почти что полностью неблагонадежный. С увеличением диаметра увеличивается смертность пихты. Деревья толще 22 см в древостое отсутствуют.
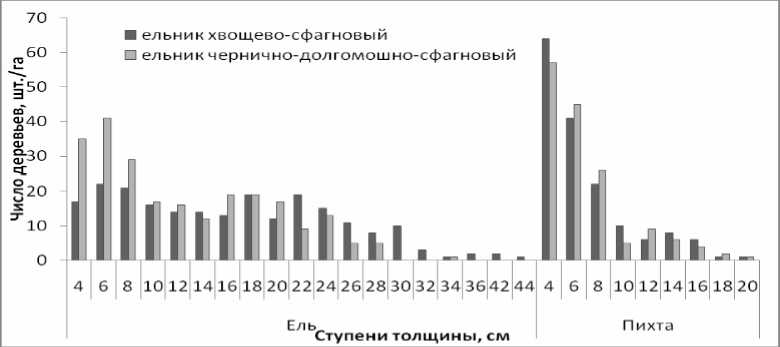
Рис. 2. Распределение деревьев ели и пихты по ступеням толщины
Анализ онтогенетической структуры показал, что и в хвощево-сфагновом и в чернично-долгомошно-сфагновом ельнике полночленна только ценопопуляция ели. В обоих сообществах достаточно высокая численность виргинильных и молодых генеративных особей, что свидетельствует об устойчивом потоке поколений. Распределение близкое к нормальному. Ель возобновляется на микросайтах разных типов, но больше всего на крупном валеже средней и поздней стадии разложения. Онтогенетические спектры це-нопопуляций пихты иные (рис. 3). Несмотря на то, что в обоих сообществах, достаточно много виргинильных особей 1 подгруппы, численность особей следующих состояний значительно меньше, а зрелые и старые генеративные особи отсутствуют вовсе. Распределение близко к экспоненциальному. Оно свидетельствует о том, что в обоих сообществах отсутствуют условия для нормального развития пихты, поэтому она редко встречается в составе древостоя, хотя постоянно присутствует в подросте, часто принимая стланиковую форму роста. Семенное возобновление пихты очень редко встречается на валеже, преобладает вегетативное возобновление в межкроновом и подкроновом пространствах.
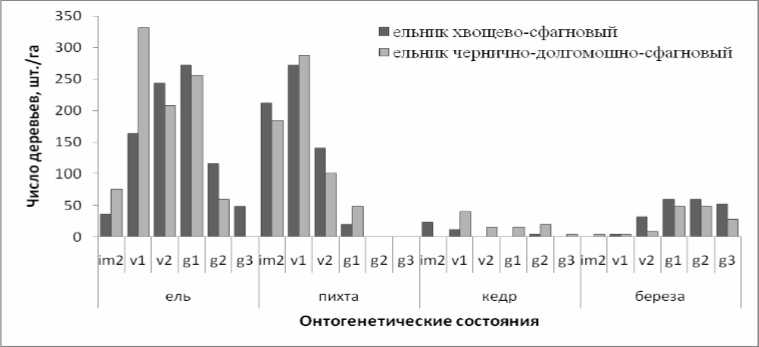
Рис. 3. Онтогенетический спектр ценопопуляций ели, пихты, кедра и березы в разных сообществах
В исследованных сообществах наиболее сильно различаются онтогенетические спектры ценопопуляций кедра. В ельнике хвощево-сфагновом достаточно много имматурных, единичны виргинильные и зрелые генеративные особи. Особей других онтогенетических состояний нет. В ельнике чернично-долгомошно-сфагновом встречаются особи всех, за исключением имматурного, онтогенетических состояний, что свидетельствует о наличие условий более менее устойчивого развития ценопопуляции кедра. Возобновление кедра в обоих сообществах приурочено, главным образом, к подкроновым пространствам ели и пихты, и значительно реже к валежу поздних стадий разложения. В обоих сообществах онтогенетические спектры ценопо-популяций березы регрессивны, Это позволяет предположить, что условия приживания и развития новых поколений березы существенно изменились с момента формирования взрослой фракции ценопопуляций. Единично новые особи приживаются на свежих буграх вывалов и на валеже поздних стадий разложения.
Выводы: исследование древесной синузии сфагновых лесов верховий р. Печоры показало, что стабильную основу древесной синузии формируют устойчивые потоки поколений ценопо-пуляций ели. Присутствие в этих лесах ценопо-пуляций пихты поддерживается в результате толерантности подроста, способного длительно выживать в неблагоприятных условиях, возобновляясь только вегетативно, а при улучшении условий обитания, формировать и взрослые особи. Это позволяет предположить, что присутствие пихты в составе древесной синузии, как и ели, будет длиться неопределенно долго при отсутствии катастрофических нарушений. В цено-популяциях березы отсутствуют молодые особи. Вероятно при сохранении современных условий численность ее постепенно сократится до минимума, ценопопуляции станут фрагментарными и этот вид практически перестанет влиять на развитие древесной синузии в целом. Ценопопуля-ция кедра в ельнике чернично-зеленомошносфагновом в настоящее время способна к само-поддержанию, а в ельнике хвощево-сфагновом она имеет ложно инвазионный характер, поскольку нет условий для полного прохождения онтогенеза молодыми деревьями. Неустойчивое состояние ценопопуляций кедра в значительной части сфагновых лесов требует дальнейшего тщательного исследования биологии и экологии этого чрезвычайно ценного вида.
Работа выполнена при финансовой поддержке фонда РФФИ (проект № 10-04-00355) и Программы фундаментальных исследований Президиума РАН «Научные основы сохранения биоразнообразия России».
Список литературы Состав и строение древостоев сфагновых ельников в верховьях реки Печора (Печоро-Илычский заповедник)
- Методические подходы к экологической оценке лесного покрова в бассейне малой реки/Под ред. Л.Б. Заугольновой, Т.Ю. Браславской. М.: Товарищество научных изданий. КМК. 2010. 383 с.
- Методы изучения лесных сообществ. -СПб., 2002. 240 с.
- Смирнов, Н.С. Основные типы темнохвойных лесов бассейна р. Б. Порожняя (Печоро-Илычский заповедник)//Известия Самарского НЦ РАН. 2010. Т. 12 (33), №1 (3). С. 892-896.
- Смирнов, Н.С. Растительность темнохвойных лесов нижней части бассейна реки Большая Порожняя (приток р. Печора)/Н.С. Смирнов, Т.Ю. Браславская//Труды Печоро-Илычского заповедника. -Сыктывкар. 2010. Вып. 16. С. 149-156.
- Смирнова, О.В. Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов южной части Печоро-Илычского заповедника/О.В. Смирнова, М.В. Бобровский, Л.Г. Ханина, В.Э. Смирнов//Тр. Печоро-Илычского заповедника. 2007. Т. 15. С. 38-52.
- Смирнова, О.В. Сукцессионный статус старовозрастных темнохвойных лесов Европейской России/О.В. Смирнова, М.В. Бобровский, Л.Г. Ханина, В.Э. Смирнов//Успехи совр. биологии. 2006. № 1. С. 26-48.
