Состав нейтральных липидов Hydrilla verticillata (L. Fil.) Royle в условиях аккумуляции и элиминации ионов тяжелых металлов
 Royle в условиях аккумуляции и элиминации ионов тяжелых металлов")
Автор: Нестеров В.Н.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 4 т.9, 2007 года.
Бесплатный доступ
Исследовали особенности аккумуляции и элиминации ионов тяжелых металлов (ТМ) (Cu+2, Pb+2, Zn+2) водным растением Hydrilla verticillatа. Как накопление, так и выведение ионов ТМ, зависело от времени и природы ТМ. Показана роль общих и нейтральных липидов в ответных реакциях на воздей- ствие ионов ТМ как одного из неблагоприятных абиотических факторов антропогенного характера. Выявлено, что несмотря на разную направленность действия ионов ТМ, имеет место перестройка липидного метаболизма растительной клетки с участием триглицеридов как формы хранения энергии. Так же происходят процессы, направленные на регулирование поступления ионов ТМ и повышение устойчивости к данным токсикантам за счет изменения соотношения стерины/эфиры стеринов. Об- щая роль липидов в адаптации к ранее не встречающимся факторам окружающей среды заключается в изменении содержания общих липидов, соотношения индивидуальных нейтральных липидов.
Короткий адрес: https://sciup.org/148198022
IDR: 148198022 | УДК: 581.1
Neutral lipids compositions Hydrilla verticillata (L. Fil.) Royle in conditions of accumulation and elimination of ions of heavy metals
The features of accumulation and elimination of ions of heavy metals (HM) (Cu+2, Zn+2, Pb+2) in water plant Hydrilla verticillata were investigated in this work. Both accumulation and removal of ions of HM depends on time and nature of HM. The responded reactions of neutral lipids were shown in this work as well as the impact of HMs ions of those reactions. Despite the different orientation of impact of HM ions, have place change in lipids consists: contents of common lipids and ratio of individual neutral lipids - triglycerides, free sterols and his esters, waxes.
Текст научной статьи Состав нейтральных липидов Hydrilla verticillata (L. Fil.) Royle в условиях аккумуляции и элиминации ионов тяжелых металлов
Постоянно растущее загрязнение окружающей среды является наиболее угрожающим фактором для нормального функционирования живых организмов [1]. Особое место среди антропогенных воздействий занимают тяжелые металлы (ТМ), так как они представляют собой наиболее широко распространенные поллютанты для многих форм жизни [2, 3].
Часто макрофиты служат индикаторами природной среды. В то же время сами макрофиты оказывают значительное влияние на качество воды [4, 5]. Учитывая тот факт, что многие водоемы представляют собой совокупность лимнических и лотических систем, наряду с оценкой аккумуляции ионов ТМ казалось существенным провести исследование элиминирующей способности водных растений для данных токсикантов.
Одним из адаптивных ответов растительных клеток на воздействие ТМ является изменение липидного метаболизма. Поскольку в последнее время большое количество работ посвящено исследованию экологической роли полярных липидов, а нейтральные липиды (НЛ) наряду с полярными липидами, составляют основу липидного комплекса клетки и участвуют во многих метаболических процессах, большой интерес представляло изучение особенностей состава НЛ под действием ионов ТМ [6, 7].
Целью данной работы было изучить особенности аккумуляции и элиминации ионов ТМ (Cu2+, Zn2+, Pb2+) высшими водными растениями на примере вида Hydrilla verticillata и выявить роль нейтральных липидов в данном процессе.
Методика
Растительный объект. Для исследования выбрано водное погруженное растение Hydrilla verticillata (L. fil.) Royle (гидрилла мутовчатая). Данный вид, широко распространенный в водоемах Европы, является удобным объектом для лабораторных исследований, поскольку неприхотлив к минеральному составу воды, отличается быстрым ростом и не требует обязательного укоренения [8].
Постановка эксперимента . Навески растения в количестве 3 г помещались в стеклянные емкости (объемом 1 л) с отстоянной водопроводной водой, в которые добавляли заданные количества солей металлов Cu, Zn,
Pb (в форме нитратов) таким образом, чтобы концентрация составила 100 мкмолей/л. В данных условиях растения выдерживались 1, 3, 10 суток. По истечении заданного времени эксперимента образцы растения промывали в проточной воде, далее одну часть растений использовали для анализов, а другую часть помещали в отстоянную водопроводную воду. Эта часть растения служила для наблюдения за элиминацией ТМ и биохимическим состоянием испытуемого объекта в период реабилитации. Время реабилитации составило 5 суток. Растения выращивались в контролируемых условиях освещения (800±200 лк) и температуры (200С) при 10часовом световом дне.
Экстракция и анализ липидов . Экстракцию липидов из тканей растений осуществляли по модифицированному методу Блайя и Дайэра [9]. Количество общих липидов в экстрактах определяли взвешиванием высушенных в вакууме до постоянного веса аликвот экстракта [10].
Разделение НЛ проводилось методом одномерной двукратной тонкослойной хроматографии на стеклянных пластинках (10 x 10 см) с закрепленным слоем силикагеля с использованием систем растворителей: толуол - гексан - муравьиная кислота (объемное соотношение 140:60:1), гексан - диэтиловый эфир - муравьиная кислота (объемное соотношение 60:40:1).
Проявление липидов осуществляли обрызгиванием пластинок 10%-ым раствором H2SO4в метаноле и последующем нагревании их при температуре 1800C в течение 15 мин.
Количество НЛ определялось денситометрически.
Определение содержания ТМ в тканях растений проводили по методу Голубкиной [11]. После обесцвечивания растворов концентрацию ТМ измеряли на атомно-абсорбционном спектрофотометре МГА-915 (Россия).
В таблицах и рисунках приведены данные, представляющие собой средние величины из 3 биологических повторностей, каждую из которых анализировали в трех параллельных повторностях.
Результаты и обсуждение
Известно, что погруженные гидрофиты, в категорию которых относится исследуемый нами вид Hydrilla verticillata, обладают рядом биологических особенностей. Среди последних отмечается высокая проницаемость кутикулы листьев, слабое развитие корней и т.д. В связи с этим поглощение ионов ТМ отличается [12].
С помощью атомно-абсорбционного анализа было установлено, что в условиях нашего эксперимента накопление и выведение ионов металлов H. verticillata зависело от природы металла и индивидуальной реакции растения на него. Например, цинк накапливался прогрессивно при увеличении времени воздействия ТМ. Аккумуляция свинца и меди происходила таким образом, что достигнув максимальных значений со временем снижалась. По-видимому, накопительная способность данного растения в отношении меди и свинца ограничена и со временем часть накопленного металла способна вымываться из растения (табл. 1). Характер элиминации ионов ТМ из организма растения также был различным: выведение цинка и свинца из тканей исследуемого растения происходило быстрее, чем удаление меди. Кроме того, медь в условиях данного эксперимента оказалась менее «элиминируемым» элементом, и чем дольше была экспозиция растения данным токсикантом, тем меньше его выводилось в постстрессовый период.
Результаты проведенных исследований по влиянию ионов ТМ на липиды показали, что действие ионов меди достоверно снижало содержание общих липидов (ОЛ) в исследуемом растении (рис. 1). В результате воздействия ионов меди на H. verticillata ионами меди произошло практически двукратное уменьшение количества ОЛ. В период реабилитации после действия ионов меди содержание ОЛ несколько возрастало (рис. 2). Ионы цинка в среде стимулировали увеличение числа ОЛ до 13,6 мг/г сырого веса (рис. 1). Причем в период реабилитации в растениях отмечено повышенное по сравнению с контролем содержание ОЛ. Если в контроле количество ОЛ составило 7,0 мг/г сырого веса, то после воздействия ионами цинка и последующей реабилитации в течении 5 дней количество ОЛ стало равным 9 мг/г (рис. 2).
Таблица 1. Динамика аккумуляции и элиминации ионов ТМ водным растением Hydrilla verticillata (время реабилитации 5 суток)
|
Металл |
Воздействие |
Концентрация металла в растении, мг на г сухой массы |
||
|
Время воздействия, сутки |
||||
|
1 |
3 |
10 |
||
|
Медь |
и £ < |
5 |
8,2 |
5,5 |
|
Цинк |
0,2 |
2,4 |
3,3 |
|
|
Свинец |
9,3 |
9,7 |
4 |
|
|
Медь |
3,5 |
3,8 |
4,7 |
|
|
Цинк |
0 |
0,4 |
0,7 |
|
|
Свинец |
1,1 |
0,66 |
0,5 |
|
Ионы свинца, как в период аккумуляции, так и в период элиминации, действовали на H. verticillata так же стимулирующее (рис. 2). Наибольшее их значение зафиксировано после возвращения растения в нормальные условия (18,3 мг/г сырого веса).
Таким образом, различные металлы по-разному изменяли содержание суммарных липидов в тканях H. verticillata. Медь действовала угнетающе, а свинец и цинк стимулирующе в стрессовой фазе. В постстрессовый период, т.е. после прекращения воздействия, количество ОЛ способно было восстанавливаться в случае с цинком и даже превосходить контроль в случае со свинцом.
Суммарные или общие липиды, выделяемые по используемой нами методике, обычно представляют собой смесь полярных и нейтральных липидов. Среди НЛ исследуемого вида нами были идентифицированы следующие компоненты: углеводороды, триглицериды, стерины, эфиры стеринов, воски, кислоты, спирты, 1,2- и 1,3-диглицериды (табл. 2). Все составляющие НЛ выполняют различные функции. Так, стерины рассматриваются как важные элементы мембран, регулирующие их ионную проницаемость. Воски выступают в качестве защитного материала. Триглицериды являются составляющими НЛ, которые отвечают за запасание энергии. Остальные компоненты НЛ можно рассматривать как промежуточные вещества в
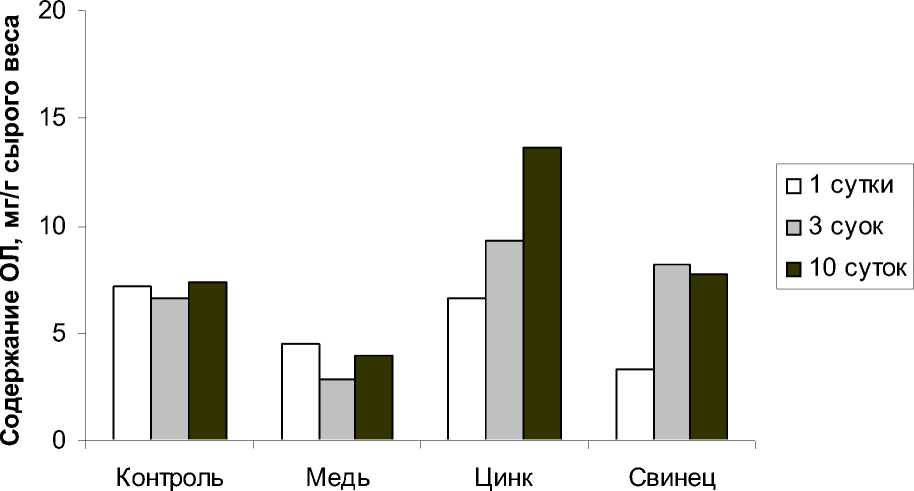
Рис. 1. Содержание общих липидов в Hydrilla verticillata под влиянием ТМ (100 мкмоль/л, аккумуляция)
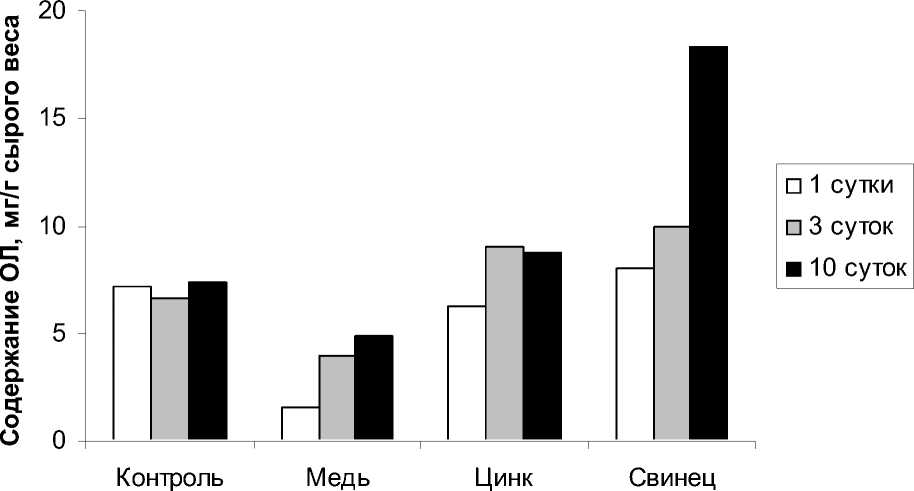
Рис. 2. Содержание общих липидов в Hydrilla verticillata под влиянием ТМ (100 мкмоль/л, элиминация)
синтезе других липидов, хотя свободные кислоты некоторые авторы рассматривают в качестве трансмембранных переносчиков ряда физиологически активных катионов [13], а спирты как регуляторы роста [14].
Под действием ТМ были отмечены изменения в соотношении индивидуальных компонентов НЛ (табл. 2). Причем изменения эти были специфическими и зависели от природы ТМ. Так, под действием ионов меди происходило изменение соотношения практически всех составляющих НЛ. Тогда как при действии ионов цинка в течение 1, 3 суток достоверных изменений в соотношении НЛ не отмечено. Только через 10 суток воздействия данным металлом произошли существенные изменения, выражающиеся, главным образом, в снижении доли стеринов, которое происходило на фоне увеличения триглицеридов и спиртов. Интересно отметить, что действие ионов свинца на компоненты НЛ было аналогичным действию ионов цинка.
Представляло интерес провести сравнительный анализ динамики содержания тех компонентов, которые изменялись под действием всех трех металлов. Этими компонентами оказались стерины, воски, триглицери- ды.
Растительные стерины, подобно холестерину, условно относятся к НЛ. Однако они являются обычными компонентами биологических мембран. Так же как и фосфолипиды, растительные стерины обладают слабыми амфифильными свойствами, т.е. имеют в своей молекуле полярную «головку» (спиртовой участок) и неполярный «хвост» (остальная часть молекулы). Стерины хорошо встраиваются в фосфолипидные слои, образующие клеточные мембраны. В норме они играют роль регуляторов агрегатного состояния липидных ансамблей, образующих биомембраны: слишком плотные мембраны они как бы разжижают, а слишком жидкие, наоборот уплотняют, ограничивая подвижность гидрофобных «хвостов» фосфолипидов [15].
На рис. 3 представлены сравнительные данные по влиянию ионов ТМ на содержание стеринов. В контрольных образцах в начале эксперимента содержание стеринов составило 11,9% от суммы НЛ. Действие ионов меди проявилось в снижении данного показателя и составило в первые сутки 7,7%, а на 10 сутки - 5,7%. Действие ионов свинца и цинка так же снижало содержание стеринов,
Таблица 2. Изменение состава нейтральных липидов в Hydrilla verticillata под влиянием ионов ТМ
|
Металл/ воздействие |
Время воздействия, сутки |
Содержание нейтральных липидов, % от суммы |
||||||||
|
Углеводороды |
Эфиры |
Воски |
Триглицериды |
Кислоты |
Спирты |
Стерины |
Диглицериды |
|||
|
Контроль |
0 |
20,8 |
7,1 |
5,8 |
8,9 |
11,8 |
8,5 |
11,9 |
25,3 |
|
|
1 |
31 |
7,7 |
5,0 |
9,6 |
15,1 |
8,5 |
11,2 |
12,3 |
||
|
Сu |
3 |
26,3 |
6,3 |
4,6 |
10,8 |
14,9 |
13,9 |
7,1 |
16,4 |
|
|
§ =г |
10 |
41,0 |
7,1 |
3,4 |
9,9 |
11,5 |
10 |
5,4 |
12,0 |
|
|
с; |
1 |
23,5 |
7,9 |
4,9 |
10,7 |
11,5 |
10,7 |
9,3 |
21,5 |
|
|
Zn |
LQ |
3 |
21,1 |
6,5 |
4,3 |
10,9 |
11,7 |
7,3 |
11,6 |
26,6 |
|
<С Ш |
10 |
33,9 |
6,6 |
4,9 |
7,0 |
12,9 |
7,1 |
9,3 |
18,4 |
|
|
1 |
22,3 |
7,1 |
3,8 |
8,4 |
12,4 |
6,8 |
11,7 |
27,6 |
||
|
Pb |
3 |
24,9 |
6,4 |
2,4 |
7,5 |
12,0 |
9,3 |
12,6 |
25,2 |
|
|
10 |
33,7 |
6,0 |
2,4 |
5,9 |
11,5 |
6,8 |
10,6 |
23,2 |
||
|
1 |
21,5 |
8,1 |
5,8 |
13,8 |
15 |
11,6 |
7,7 |
16,9 |
||
|
Сu |
3 |
22,8 |
7,4 |
4,4 |
11,1 |
17,7 |
11,1 |
7,0 |
18,8 |
|
|
к |
10 |
29,2 |
9,4 |
3,9 |
12,7 |
14 |
11,1 |
5,7 |
14,3 |
|
|
=г |
1 |
20,5 |
6,3 |
6,3 |
9,0 |
11,8 |
11,3 |
10,9 |
23,9 |
|
|
Zn |
3 |
15,5 |
5,8 |
4,7 |
9,6 |
11,5 |
10,3 |
12,3 |
30,4 |
|
|
10 |
25,7 |
7,4 |
3,9 |
10,3 |
13,3 |
13,3 |
9,0 |
17,4 |
||
|
< |
1 |
20,3 |
6 |
4,1 |
9,2 |
12,2 |
9,4 |
11,9 |
26,9 |
|
|
Pb |
3 |
16,3 |
5,6 |
3,7 |
9,3 |
12,9 |
8,6 |
12,4 |
31,3 |
|
|
10 |
25,3 |
5,2 |
3,2 |
9,1 |
12,1 |
7,3 |
10,2 |
27,7 |
||
но происходило это только к 10 суткам экспозиции и было менее выраженным в сравнении с действием ионов меди.
Если учесть, что стерины действуют как мембранорегуляторы, то снижение их концентрации, по-видимому, ведет к разжижению мембран и увеличению их проницаемости, что в целом негативно сказывается на содержании ОЛ. Дополнительным фактом, свидетельствующим о токсическом действии ионов меди на мембрану, является изменение соотношения стерины/эфиры в сторону увеличения доли эфиров стеринов (рис. 4). Выявлено, что ионы цинка и свинца не оказывали существенного влияния на данные показатели.
Наружная поверхность клеточных стенок эпидермальных клеток покрыта гидрофобной кутикулой, которая состоит из кутина и вос- ка. Эти вещества входят в покровные ткани растений и необходимы для предотвращения потери влаги и защиты от патогенных микроорганизмов или вредных веществ [16]. В условиях нашего эксперимента соединениями, выполняющими в растении защитную функцию, являются воски. Все используемые ТМ достоверно снижали содержание восков (рис. 5). Причем в большей степени этому способствовали ионы свинца и цинка. Так же было замечено, что чем дольше длилось воздействие ТМ, тем значительнее снижалась доля восков в составе растения.
Как отмечалось выше, одной из важных функций НЛ является запасание энергии.
Эту роль выполняют триглицериды, которые служат формой хранения углерода и представляют собой важные вместилища питательных веществ [17]. Данные рис. 6
свидетельствуют о том, что количество триглицеридов увеличивалось под действием ионов меди с 8,9% до 13,8% относительно контроля. Достоверное изменение доли триглицеридов зафиксировано только при 10-суточном воздействии ионов цинка. Превыше- ние данного показателя, по сравнению с контрольными значениями, составило 15,7%. Действие ионов свинца не оказывало значительного влияния на содержание триглицеридов (рис. 6).
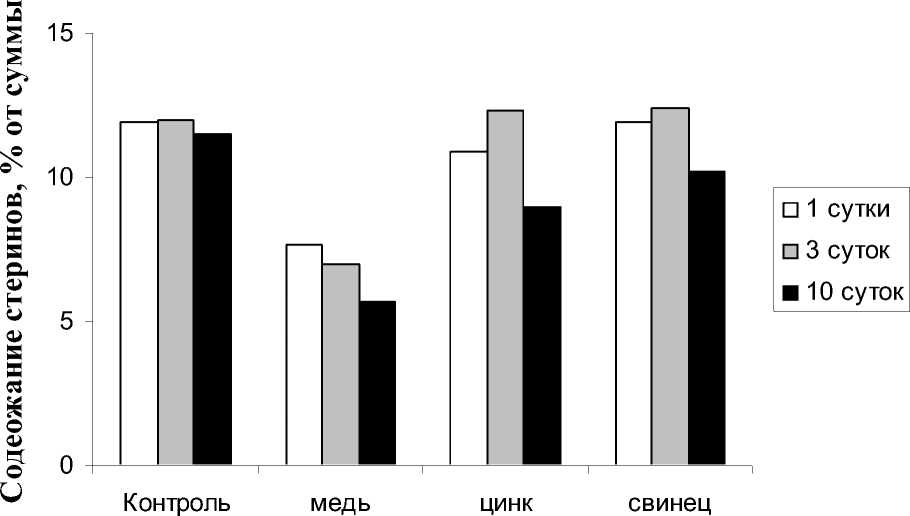
Рис. 3. Изменение содержания стеринов в Hydrilla verticillata при аккумуляции ионов ТМ.

30%
20%
10%
0% П I I I I I I I I I
К 1 3 10 К 1 3 10
Аккумуляция/реабилитация(время воздействия, сутки)
Рис 4. Изменение соотношения стерины/эфиры под действием ионов меди
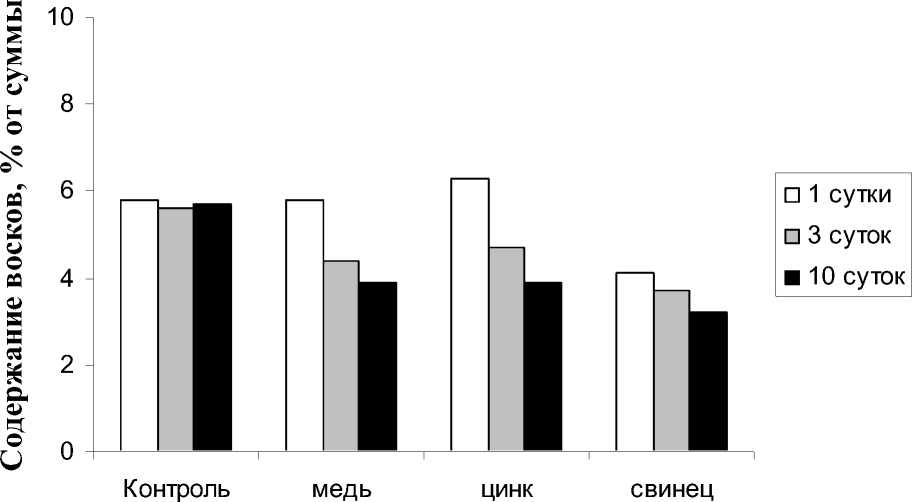
Рис. 5. Изменение содержания восков в Hydrilla verticillata при аккумуляции ионов ТМ.
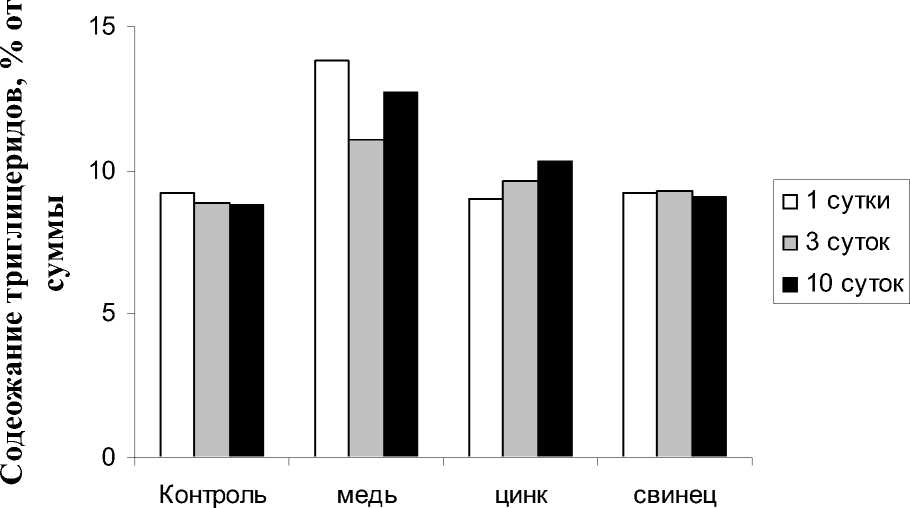
Рис. 6. Изменение содержания триглицеридов в Hydrilla verticillata при аккумуляции ионов ТМ.
Следует отметить, что из всех используемых ТМ, ионы меди оказывали более сильный стрессовый эффект на испытываемое растение. Повышение количества триглицеридов может говорить о сохранении энергии, часто отмечаемое при стрессовых ситуациях
[18], направленное на выживание биологического вида в неблагоприятных условиях [19].
Важным этапом настоящей работы было исследовать состав НЛ в процессе реабилитации, т.е. после снятия «металлического»
пресса. На рис. 7-9 представлены результаты количественных изменений стеринов, восков и триглицеридов в процессе реабилитации. При возвращении исследуемых растений в нормальные условия происходили изменения в содержании индивидуальных НЛ, завися- щие как от природы ТМ, так и продолжительности воздействия данными токсикантами. В частности, происходило практически полное восстановление содержания стеринов при 1-суточном воздействии ионов меди и последующей 5-дневной реабилитации. Су-
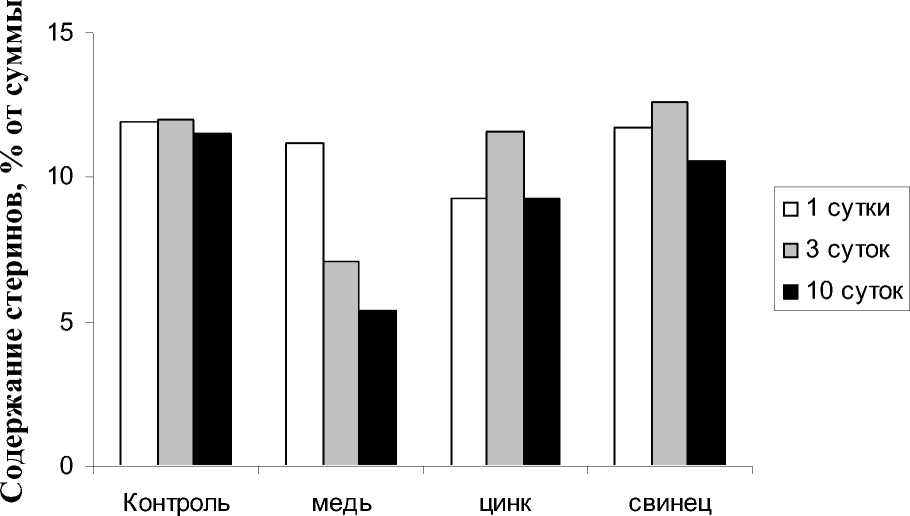
Рис. 7. Изменение содержания стеринов в Hydrilla verticillata при реабилитации (5 суток) ионов ТМ.
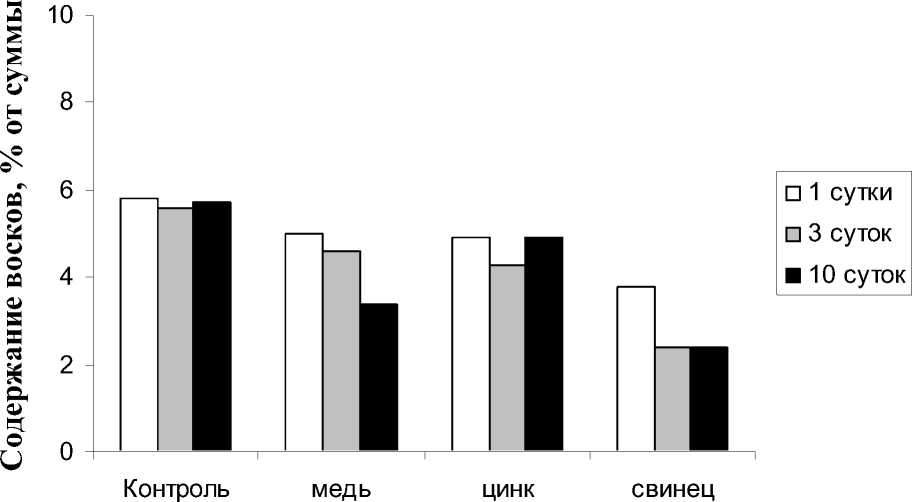
Рис. 8. Изменение содержания восков в Hydrilla verticillata при реабилитации (5 суток) ионов ТМ.
щественных изменений данного компонента НЛ при реабилитации после влияния ионов цинка и свинца не выявлено.
Общей особенностью для всех ионов ТМ стало снижение количества восков в процессе реабилитации по отношению к периоду аккумуляции ТМ.
Что касается триглицеридов, то их реакция в фазе реабилитации была неоднозначной. Если действие ионов меди при их на- коплении увеличивало содержание триглицеридов, то реабилитация приводила к восстановлению уровня данных веществ, соответствующего контролю, т.е. снижению.
В отношении ионов цинка и свинца можно отметить, что, несмотря на то, что уровень триглицеридов при аккумуляции этих металлов, как правило не менялся, реабилитация приводила к снижению относительного вклада указанных структур НЛ.
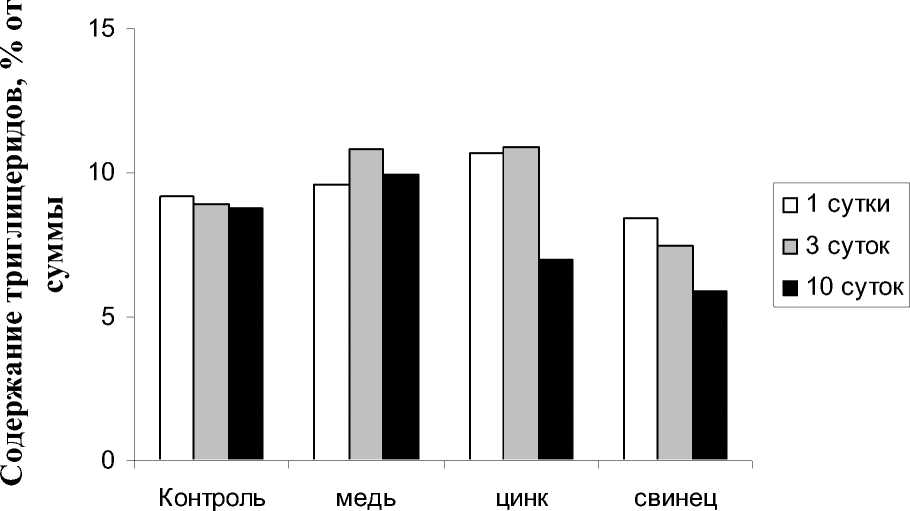
Рис. 9. Изменение содержания триглицеридов в Hydrilla verticillata при реабилитации (5 суток) ионов ТМ.
Выводы
Полученные результаты свидетельствуют о том, что нейтральные липиды принимают активное участие в ответных реакциях на воздействие ионов ТМ. Выявлены общие и специфические особенности динамики состава компонентов нейтральных липидов у Hydrilla verticillata под действием ионов Cu2+, Zn2+, Pb2+ при различных временных интервалах. Показано, что в зависимости от ток-сичночсти ТМ происходит соответствующая перестройка липидного метаболизма расти- тельной клетки с целью сохранения энергии с участием триглицеридов. Кроме того, происходят процессы, направленные на регулирование поступления ионов ТМ и повышение устойчивости к данным токсикантам за счет изменения соотношения стерины/эфи-ры стеринов. На клеточном уровне исследование содержания нейтральных липидов, представляет не меньший интерес для понимания механизмов адаптации растений на изменившиеся условия, чем мембранные полярные липиды.
Список литературы Состав нейтральных липидов Hydrilla verticillata (L. Fil.) Royle в условиях аккумуляции и элиминации ионов тяжелых металлов
- Гуральчук Ж.З. Механизмы устойчивости растений к тяжелым металлам//Физиол. и биохим. культ. раст. 1994. Т. 26.
- Юрин В.М. Основы ксенобиологии. Учебное пособие. Минск: Новое знание, 2002.
- Ипатова В.И. Адаптация водных растений к стрессовым абиотическим факторам. М.: Графикон-принт, 2005.
- Усманов И.Ю., Рахманкулова З.Ф., Кулагин А.Ю. Экологическая физиология растений. -М.: Изд-во «Логос», 2001.
- Прасад М.Н. Практическое использование растений для восстановления экосистем загрязненных металлами//Физиология растений. 2003. Т. 50, № 5.
- Иванова А.Б., Гордон Л.Х., Лыгин А.В. Роль структурных липидов в регуляции ионного транспорта растительных клеток//Цитология. 1997. Т. 39, № 4/5.
- Васьковский В.Е. Липиды//Соросовский образоват. журн. 1997. № 3.
- Полонский А.С. Мир аквариума. Калининград. кн. изд-во. 1994.
- Bligh E.G., Dyer W.J. A rapid method of lipid extraction and purification//Can. J. Biochem. Physiol. 1959. V. 37.
- Vaskovsky V.E., Latyshev N.A. Modified Jungnickel s reagent for detecting phospholipids and other phosphorus compounds on thin-layer chromatograms//J. Chromatogr. 1975. V. 115.
- Голубкина Н.А. Флуорометрический метод определения селена//Журн. аналит. химии. 1995. Т. 50.
- Лукина Л.Ф., Смирнова Н.Н. Физиология высших водных растений. Киев: Наук. думка, 1988.
- Розенцвет О.А., Саксонов С.В., Козлов В.Г., Конева Н.В. Эколого-биохимический подход к изучению липидов высших водных растений//Изв. Самар. НЦ РАН. 2000. Т. 2, № 2.
- Иванова А.Б., Гордон Л.Х., Лыгин А.В. Роль структурных липидов в регуляции ионного транспорта растительных клеток//Цитология. 1997. Т. 39, № 4/5.
- Бергельсон Л.Д. Биологические мембраны. М.: Наука, 1975.
- Гудвин Т., Мерсер Э. Введение в биохимию растений: В 2 т. М.: Мир, 1986. Т. 1.
- Ohlrogge J., Browse J. Lipid Biosynthesis//The Plant Cell. 1995. V. 7.
- Кузнецов В.В., Дмитриева Г.А. Физиология растений. М.: Высш. шк., 2006.
- Хочачка П., Сомеро Д. Биохимическая адаптация. Пер. с англ. М.: Мир, 1988.
