Состояние и перспективы развития семеноводства картофеля в Тюменской области

Автор: Логинов Юрий Павлович, Казак Анастасия Афонасьевна, Кендус Ксения Алексеевна, Ященко Сергей Николаевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-2 т.20, 2018 года.
Бесплатный доступ
Производство качественных семенных клубней - основа успешного развития картофелеводства в Тюменской области. Освящено состояние семеноводства в лучших хозяйствах области Агрофирме КРиММ Упоровского района, в КХ Дружба-Нива Заводоуковского района, которые работают полностью на сортах зарубежной селекции. Для производства семенного материала сортов отечественной селекции в ГАУ Северного Зауралья создаётся биолаборатория по получению оздоровленных клубней, а также хозяйство по производству суперэлиты и элиты. Представлены данные урожайности и качества клубней в зависимости от элементов технологии возделывания.
Картофель, сорт, семеноводство, урожайность, качество клубней
Короткий адрес: https://sciup.org/148313901
IDR: 148313901 | УДК: 635.21(631.5)571.12
State and the prospects of development of seed farming of potatoes in the Tyumen region
Production of qualitative seed tubers - a basis of successful development of potato growing in the Tyumen region. The condition of seed farming in the best farms of area to Agricultural firm КРиММ of Uporovsky district, in KH Druzhba-Niva of Zavodoukovsky district which work completely at grades of foreign selection is consecrated. For production of seed material of grades of domestic selection in GAU of the Northern Trans-Ural region the biolaboratory on receiving the revitalized tubers and also economy on production of superelite and elite is created. Data of productivity and quality of tubers depending on elements of technology of cultivation are submitted.
Текст научной статьи Состояние и перспективы развития семеноводства картофеля в Тюменской области
Предпосылкой в усовершенствовании технологии выращивания свеклы явилась необходимость выведения сортов, дающих при прорастании один росток.
Такие односемянные спонтанные мутанты были найдены и послужили исходными формами для создания одноростковых сортов и компонентов гибридов свеклы. Первый сорт односемянной свеклы создан в 1956 году [1, 2].
Лишь создание гибридов на основе цитоплазматической мужской стерильности с одно-семянностью свыше 95 % позволяет выращивать свеклу по современной технологии. В настоящее время в мире отсутствует гибрид со 100 % стабильностью этого признака. Только благодаря интенсивной подготовке семян удается довести их до односемянности 95 %.
Ошевнев Валерий Павлович, доктор сельскохозяйственных наук, заведующий лабораторией селекции сахарной свеклы на основе ЦМС. Е-mail: 5891542@ mail.ru Грибанова Нина Павловна, кандидат сельскохозяйственных наук, ведущий научный сотрудник лаборатории селекции сахарной свеклы на основе ЦМС Васильченко Елена Николаевна, старший научный сотрудник отдела биотехнологии.
Бердников Роман Владимирович, кандидат сельскохозяйственных наук, старший научный сотрудник лаборатории селекции сахарной свеклы на основе ЦМС
Причину нестабильности односемянности следует искать в полигенном наследовании этого признака. Отсюда для практической селекции и первичного семеноводства следует вывод о необходимости постоянного и строгого отбора по этому признаку.
В настоящее время считается установленным, что признак односемянности имеется в любой популяции сахарной свеклы и носит рецессивный характер.
Раздельноплодные формы в сростноплод-ных популяциях возникают, по-видимому, в результате естественного мутационного процесса [3, 4]. Мутанты могут скрещиваться со сростно-плодной свеклой или находиться в виде авто-фертильных растений.
В опытах Савицкого В.Ф. по наследованию признака раздельноплодности убедительно и на большом фактическом материале показан многогибридный характер расщепления по локусу М-m. Однако в работах других авторов можно найти как подтверждение этих данных, так и наличие резких отклонений от моногибридной схемы.
Sedlmayr K. [5] объясняет это наличием селективного оплодотворения как между растениями одной группы, так и в пределах отдельных растений, Knapp E. [6] связывает отклонения от моногибридной схемы с различиями исходных раздельноплодных материалов при использовании и гибридизации односемянных форм из России и Германии. Отклонения от многогибридной схемы известны и в более поздних исследованиях. Многочисленные случаи отклонений от моногибридной схемы расщепления приводят иногда к представлению, что генетический контроль признака односемянности у европейских селекционных материалов иной чем у американских. С.И. Малецкий [7] на основании проведенных серий скрещиваний предполагает наличие двухлокусной модели наследования (mmIi) признака односемянности.
Сростноплодные растения постоянно появляются при репродуцировании раздельноплодных сортов – популяций или компонентов гибридов. Обязательным методическим приемом для поддержания односемянности на необходимом уровне является браковка. Проведение ежегодных поддерживающих отборов односемянных популяций поставил вопрос о генетической природе односемянных растений независимо от их континентального происхождения. Этот вопрос до сих пор не решен. В последнее десятилетие в рамках международных программ селекции сортов и гибридов накопилась дополнительная информация, описывающая особенности проявления признака односемянности в селекционных образцах – Ошевнев В.П. [8].
Следует отметить, что и потомства, полученные от самоопыления растений со 100 % одно-семянностью, не всегда были полностью односемянными. Мы попытались проследить связь между односемянностью F1 на основе ЦМС и односемянностью потомств, полученных от ин-цухта опылителей закрепителей стерильности, с которыми проводилась гибридизация. Такая связь не прослеживалась.
Для поиска новых опылителей закрепителей стерильности (О-Типа) и поддержание в чистоте имеющихся проводили парные скрещивания под бязевыми изоляторами. Кандидаты опылителя О-типа создавались путем скрещивания различных по происхождению односемянных селекционных образцов, индивидуальных отборов и инцухта. По односемянности растения классифицировались следующим образом: односемянные, которые имеют только односемянные плоды или одно-двусемянные только на центральном стебле; одно-двусемянные с содержанием односемянных семян 80-90 %; многосе-мянные на ветвях второго порядка, когда наряду с односемянными плодами присутствуют двусемянные и трехсемянные. Сибсовые, насыщающие скрещивания, выращивание суперэлиты проводились на пространственно изолированных участках в посевах озимых культур.
Оценивая итоги изменения признака одно-семянности при репродуцировании фертильных по пыльце опылителей О-типа РФ 8, гибрида РМС 120, можно видеть, что при стабилизации этого признака и повышении интенсивности отбора резко возрастает доля потомств с полной односемянностью (табл. 1).
Таким путем удалось повысить долю односемянных растений как у потомств с ЦМС, так и у опылителей О-типа при размножении семенного материала по схеме: линия О-типа, мужскостерильный аналог – сибсовое скрещивание – суперэлита – элита (табл.2)
Повышение доли односемянных растений и относительно высокий уровень односемянности в семенных партиях можно считать результатами тщательного отбора на стабилизацию этого признака при репродуцировании. В настоящее время нельзя отказаться от браковки по одно-семянности при размножении мужскостерильных компонентов и опылителей О-типа. Одно-семянность у элит с браковкой на 10-15 % выше, чем у элит без отбора. Достигнутый уровень односемянности позволяет использовать семена гибридов нашей селекции, внесенных в Государственный реестр селекционных достижений РМС 120, РМС 121, РМС 129, Рамоза для выра-
Таблица 1. Наследование признака односемянности у опылителя О-типа РФ 8
|
Поколение отбора |
Среднее значение односемянности |
Доля растений |
||
|
со 100 % односемянностью |
одно-двусемянных |
многосемянных |
||
|
G i |
78,2 |
11,0 |
71,0 |
18,0 |
|
g 2 |
83,4 |
24,8 |
69,2 |
6,0 |
|
G3 |
84,8 |
38,9 |
60,2 |
0,9 |
|
g 4 |
96,0 |
68,4 |
31,1 |
0,5 |
Таблица 2. Односемянность различных партий семян гибрида РМС 120
Наличие двух или большего числа генных систем, контролирующих развитие признака односемянности, затрудняет стабилизацию этого признака в популяциях, так как по фенотипу особи нельзя судить о его генотипе по генам односемянности. Это вынуждает селекционеров постоянно вести негативный отбор растений со сростноплодными плодами.
Поэтому при включении различных линий в предбазисную партию семян необходимо проводить предварительную их оценку по M-m локусам. Надежным методом здесь могут служить односторонние циклические скрещивания, которые позволяют изучить односемянность во всех возможных комбинациях.
Согласно схемы, все имеющиеся у селекционера односемянные мужскостерильные линии скрещиваются на разных изолированных участках с имеющимися опылителями О-типа. По результатам анализа потомств по односемянности судят об идентичности генотипа родительских компонентов и на основании этого формируют компоненты суперэлиты.
Таким образом, в связи с нестабильностью фенотипического проявления признака односе-мянности селекционеры вынуждены ежегодно, на всех исходных селекционных образцах, супе- рэлите и даже элите проводить браковку растений со сростноплодными плодами. Поэтому для создания МС компонента гибрида РМС 120 со стабильной односемянностью мы использовали биотехнологию получения удвоенных гаплоидов, которые представляют собой константные нерасщепляющиеся формы, пригодные для практической селекции [9].
Гаплоидный партогенез осуществлялся путем индукции неоплодотворенных семязачатков и культивировании гаплоидных регенерантов, выделенных из опылителей закрепителей стерильности (О-типа).
Необходимыми условиями формирования гаплоидных растений являются: первичная оценка морфологических признаков, проведение цитологического и цитофотометрического анализов уровня плоидности, оптимизация этапов морфогенеза в период стабилизирующих отборов, а также выявление растений со стерильной цитоплазмой.
Проведенный PCR и RFLP-анализ с использованием рестриктазы Hind III позволил идентифицировать тип цитоплазмы у создаваемых гаплоидов сахарной свеклы по числу рестриктов. У гаплоидных микроклонов с нормальной цитоплазмой амплифицировался один фрагмент (800 п.н.). У стерильных (S) форм выявлены два продукта рестрикции 320 и 480 п.н. (рис 1).
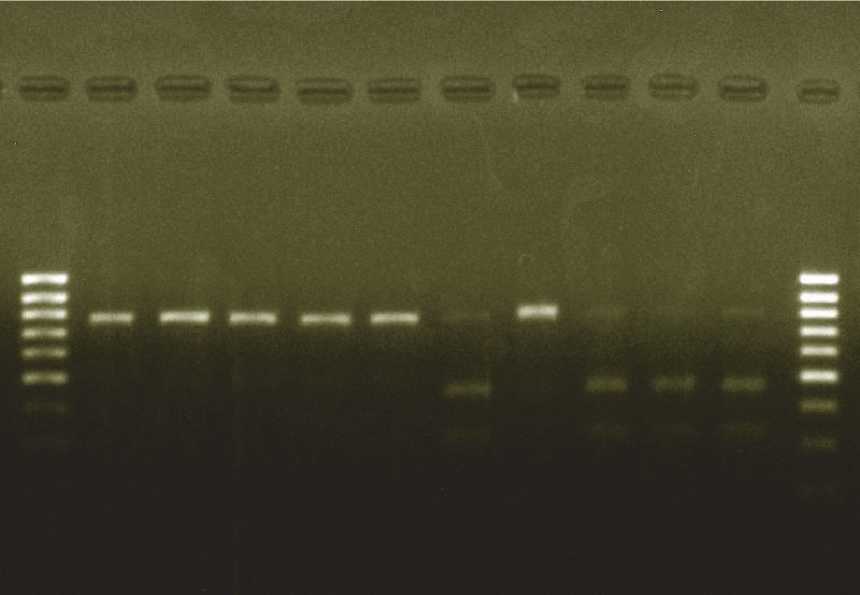
Рис. 1. Электрофореграмма рестриктов амплифицированных фрагментов ДНК (RFLP- анализ, рестриктаза Hind III) у гаплоидных растений-регенерантов сахарной свеклы (Beta vulgaris L.):
К1 — контрольные фертильные растения, К2 — контрольные стерильные растения;
1-5 — формы с нормальной (N) цитоплазмой, 6-8 — формы со стерильной (S) цитоплазмой;
М — маркеры молекулярных масс (ДНК- маркер MassRuler™, 80-1031 п.н., SM0383, «Thermo Scientific», США)
Гаплоиды, у которых этот фрагмент не ре-стрицировался Hind III, были представлены полностью фертильными формами с нормальной цитоплазмой (N) и ядерными генами в рецессивном состоянии (rf). В остальных образцах наблюдался полиморфизм фрагментов, что, по-видимому, предполагает наличие у соответствующих гаплоидных форм стерильной цитоплазмы (S) и разное сочетание рецессивных и доминантных аллелей ядерных генов Rf1/rf1 и Rf2/rf2 . Поэтому выявление растений-регенерантов со стерильной цитоплазмой на ранних этапах культивирования представляет значительный интерес для селекции сахарной свеклы, облегчая создание линий с ЦМС и высокопродуктивных гибридов на стерильной основе [10] .
Для создания жизнеспособного гомозиготного материала гаплоидные растения переводились на более высокий уровень плоидности путем колхицинирования. При дальнейшем культивировании на ростовой среде сформировались диплоидные нормально развитые микроклоны с листовыми пластинками и развитой точкой роста. Затем растения поместили в теплицу в сосуды со смесью песка с перегноем.
Удалось сохранить до 72 % микроклонов. За 3 месяца выросли небольшие корнеплоды (ште-клинги) массой от 20 до 100 грамм (рис. 2).
Индукция пониженной температурой +4о С (яровизация) проводилась в течение 45 дней. По истечении этого срока растения сформировали нормальный габитус куста, наблюдалась бутонизация и цветение (рис. 3).
Высота растений достигала 1,5 метра и выше, толщина основного цветоносного стебля составляла 3,0 – 3,5 см, цветение проходило одновременно и интенсивно, фертильность пыльцевых зерен варьировала от 87 до 92 %. Через 8 -10 недель формировались семена со 100 % раз-дельноплодностью (рис 4).
В результате были созданы 4 удвоенных гомозиготных линии (DH – линии) и получены семена для дальнейшего использования в селекционном процессе и выращивания суперэлиты мужскостерильного компонента гибрида РМС 120.
Разработанная технология позволяет ускоренно получать качественные семена гомозиготных удвоенных линий (DH) – компонентов высокопродуктивных гибридов.


Рис. 3. Семенные растения О-типа

Рис. 4. Семена гомозиготных DH-линий
Список литературы Состояние и перспективы развития семеноводства картофеля в Тюменской области
- Погадаев С.В. Производство семенного картофеля в агрофирме КРиММ/С.В. Погадаев//Селекция, семеноводство и генетика. М., 2015. № 6. С.40-41.
- Логинов Ю.П. Рекомендации по выращиванию картофеля в ЛПХ Тюменской области/Ю.П. Логинов, А.А. Казак. Тюмень, 2017. 47 с.
- Логинов Ю.П. Урожайность и качество клубней раннеспелых сортов картофеля отечественной селекции в северной лесостепи Тюменской области/Ю.П. Логинов, А.А. Казак, Л.И. Якубышина//Агропродовольственная политика. 2017. № 12 (72). С. 93-101.
- Горбунов А.К. Урожайность и качество клубней картофеля в зависимости от срока и глубины посадки/А.К. Горбунов, А.А. Васильев, А.А. Мушинский. АПК России. 2017. Т. 24. № 2. С. 322-328.
- Мушинский А.А. Адаптивная способность и стабильность по признаку урожайности сортов картофеля в условиях степной зоны Южного Урала/Мушинский А.А., Аминова Е.В., Герасимова Е.В.//В сборнике: Научное обеспечение инновационного развития сельского хозяйства в условиях часто повторяющихся засух. Материалы международной научно-практической конференции, посвященной 80-летнему юбилею Оренбургского научно-исследовательского института сельского хозяйства: сборник научных трудов. Федеральное агентство научных организаций Российская академия наук, Оренбургский научно-исследовательский институт сельского хозяйства. 2017. С. 119-124.
