Состояние и продуктивность водных экосистем. Математическое моделирование

Автор: Абакумов А.И., Пак С.Я.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Математическое моделирование экологических систем и технические науки
Статья в выпуске: 3 т.25, 2022 года.
Бесплатный доступ
Исследуется распределение фитопланктона на Западно-Камчатском шельфе Охотского моря. Сравнивается состояние фитопланктона в весенне-летне-осенние сезоны теплого 2015 года и холодного 2016 года. Математические модели используются для оценки численности фитопланктона в толще воды. Спутниковые данные о концентрации хлорофилла «а», температуре и освещенности поверхностного слоя воды используются в качестве начальных либо левых краевых условий для решения системы уравнений в математической модели.
Математическая модель, морской район, хлорофилл, минеральное питание, освещенность, температура, глубина
Короткий адрес: https://sciup.org/143178986
IDR: 143178986 | УДК: 51-76:57.087:574.5 | DOI: 10.31433/2618-9593-2022-25-3-155-157
Water ecosystems state and productivity. Mathematical modeling
In the paper, it is considered phytoplankton distribution on the Sea of Okhotsk West Kamchatka shelf. The researchers compare phytoplankton in spring-summer-autumn seasons of warm 2015 and cold 2016. They apply mathematical models to estimate the phytoplankton abundance in the water column. Satellite measurements of a sea surface are used as initial or as left boundary condition for solving a system of equations in a mathematical model. Depth illumination is calculated by differential equation.
Текст научной статьи Состояние и продуктивность водных экосистем. Математическое моделирование
Рассмотрим изменение биологических компонентов системы фитопланктон–биогены–свет с глубиной в стационарном режиме. Условие стационарности режима в системе уравнений в частных производных с диффузией приводит к следующей модели:
fd2P 1
-^=-Ve(P)-p(C,I,Ty\P dz^ к d2C у
^ = -W.i.n-MPW
VTz = -«(P.c)i .
Распределение объемной плотности P (t, z) биомассы фитопланктона по глубине z зависит от распределения концентрации C (t, z) массы минеральных питательных веществ (биогенов), освещенности (в частности, фотосинтетически активной радиации – PAR) I (t, z) и температуры T (t, z). Процесс происходит в течение времени t ∈ [0, tm] и по глубине z ∈ [0, zm]. Функция удельной скорости роста µ (C, I, T) основана на гипотезе независимо- сти выделенных влияющих факторов и выражается в видец(.с,1,т^ — Ho" HcQQ'i^iUV ИтСО, где µ0 – максимально возможная скорость роста фитопланктона, μс(С) – зависимость скорости роста от биогенов, соответственно μI(I) – от освещенности, μT(T) – от температуры. Для динамики биогенов важно, какую долю составляют учтенные минеральные вещества в растительных организмах – параметр γ, а также какова скорость возврата их в систему после бактериального разложения – параметр β, α (y, z) – удельная скорость ослабления освещенности. Здесь имеется краевая задача для z∈[0,zm], где zm – глубина фотического слоя. Данные дистанционного зондирования, характеризующие хлорофилл, температуру и облучение в поверхностном слое, могут быть использованы в качестве левого граничного условия (z = 0). Системное решение требует информации о количестве биогенов или растительного пигмента на дне или на нижней границе фотического слоя zm.
Следующая модель отображает подобный процесс посредством решения задачи Коши с использованием концепции адаптации [2, 3]:
rdP du
"T=V1"P dz dz dC
U = wf(P)C dz di где ν – определенная максимально возможная скорость роста сообщества. Функция f(P) основана на гипотезе об обратном влиянии фитопланктона на плотность минерального питания [4].
Модели, проверенные на основе данных in situ , показывают адекватное отображение динамики интегрированной биомассы в зависимости от внешних условий. Комплексное использование спутников и информации in situ дает возможность с хорошей точностью получать оценки численности фитопланктона на большой территории. Для сравнения распределений фитопланктона в теплый 2015 г. и холодный 2016 г. используется модель с функцией адаптации. Сравнение теплых и холодных лет на шельфе Западной Камчатки показывает динамику видового состава в зависимости от типа года. Основной вывод, содержащийся в литературных источниках [1], заключается в том, что видовой состав региона стабилен в многолетней динамике. Меняются только соотношения. Набор доминирующих видов изменяется в зависимости от условий окружающей среды. По объемам размножения видов при определенных температурных условиях можно сделать вывод,
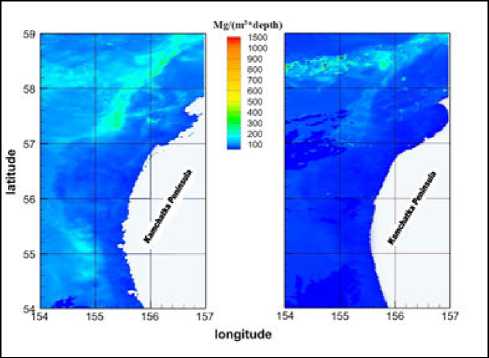
что структура доминирующих видов соответствует температурному режиму. Этот факт учитывает результаты модели в теплые и холодные годы. Видовой состав изменяется для лучшей приспособляемости сообщества фитопланктона к условиям окружающей среды.
Список литературы Состояние и продуктивность водных экосистем. Математическое моделирование
- Авраменко А.С., Черепанова М.В., Пушкарь В.С., Ярусова С.Б. Характеристика некоторых дальневосточных диатомитов // Геология и геофизика. 2015. Т. 56, № 6. С. 1206-1220.
- Beckmann A., Schaum C.-E., Hense I. Phytoplankton adaptation in ecosystem models //j. Theor. Biol. 2019. N 468. P. 60-71.
- Sekerci Y. Adaptation of species as response to climate change: predator-prey mathematical model // AIMS Mathematics. 2020. N 5 (4). P. 3875-3898. DOI: 10.3934/math.2020251.
- Sunda W.G. Feedback Interactions between Trace Metal Nutrients and Phytoplankton in the Ocean // Frontiers in Microbiology. 2012. N 3. P. 204. DOI: 10.3389/fmicb.2012.00204.
