Состояние и воспроизводство нижнедонских популяций копеечника крупноцветкового (Hedysarum grandiflorum Pall., Fabaceae)
")
Автор: Карасва Татьяна Александровна, Ермолаева Ольга Юрьевна, Бакулин Семн Дмитриевич, Пукалов Михаил Евгеньевич
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2022 года.
Бесплатный доступ
Представлены итоги мониторинговых исследований популяций Hedysarum grandiflorum в бассейне Нижнего Дона (Ростовская обл.). Выявлено 8 новых ценопопуляций вида в регионе, описаны условия их произрастания. Количественные характеристики популяций и фитоценотическая роль H. grandiflorum в составе ценозов ниже, чем в оптимальных условиях произрастания вида. По возрастной структуре популяции нормальные, неполночленные, реже полночленные, с правосторонним или центрированным онтогенетическим спектром. По классификации «дельта-омега» они отнесены к зрелым и зреющим, по величине индекса замещения - к неустойчивым, реже перспективным. Основными лимитирующими факторами в изученных популяциях выступают антропогенное воздействие, преимущественно в форме пасквальной нагрузки, и, в меньшей степени, частичное несоответствие климатических условий экологическим потребностям вида. Для H. grandiflorum в ценопопуляциях Аксайского р-на характерны крайне низкие значения числа и доли полноценных семян на плод, что в большей степени обусловлено нарушениями созревания завязавшихся семян. Значения реальной семенной продуктивности на плод существенно варьируют по годам наблюдений, что может обусловливать значительные колебания общей семенной продуктивности и выступать одной из причин нерегулярного семенного возобновления. В благоприятные для вызревания семян годы семенную продуктивность растений копеечника можно считать удовлетворительной.
Hedysarum grandiflorum, ценопопуляция, онтогенетическая структура, семенная продуктивность
Короткий адрес: https://sciup.org/147237568
IDR: 147237568 | УДК: 581.522
Conditions and reproduction of lower don populations of Hedysarum grandiflorum Pall. (Fabaceae)
The article presents the results of the monitoring research of Hedysarum grandiflorum Lower Don basin populations (Rostov Region). Eight new populations of the species are revealed in the region, their habitat conditions are described. Quantitative characters of populations studied and coenotic role of H. grandiflorum are lower than it is observed in optimal conditions. In relation to their ontogenetic structure, the populations are normal, mostly incomplete, having right-sided or centered ontogenetic spectrum. According to delta-omega classification, they belong to mature and ripening; according to replacement index, most of them are unstable or rarely perspective. The principal limiting factors in populations observed are an anthropogenic impact, mainly grazing, and partially discordance between climatic conditions of the region and ecologic requirements of the plant species. H. grandiflorum coenopopulations of the Aksay District are characterized by extremely low number and proportion of the viable seeds per fruit because of seed developmental disorder. Actual seed productivity values vary significantly between years that may condition fluctuations of total seed productivity per individual plant and cause an irregular renewal by seed. In years good for seed maturing, seed productivity of H. grandiflorum may be considered satisfactory.
Текст научной статьи Состояние и воспроизводство нижнедонских популяций копеечника крупноцветкового (Hedysarum grandiflorum Pall., Fabaceae)
Hedysarum grandiflorum Pall. (копеечник крупноцветковый) – ксерофильный каудексообразующий многолетник, наиболее широкоареальный представитель секции Subacaulia (Boiss.) B. Fedtsch.; восточноевропейский по характеру распространения вид, ареал которого охватывает среднее и нижнее течение Дона, Приволжскую возвышенность, Ергени, Заволжье, Южный Урал; за пределами России произрастает в пределах Донецкой и Луганской народных республик, Днепропетровской обл. Украины, известны местонахождения на территории Болгарии и Румынии [Васильева, 1987; Дёмина, Никитина, 2008; Шишлова, Шмараева, 2014]. В Красной книге Российской Федерации копеечник крупноцветковый имеет категорию статуса редкости 3 в – редкий вид, большая часть ареала которого находится в России. Произрастание вида подтверждено современными находками в 12 регионах России. В Пензенской обл. вид считается исчезнувшим [Новикова, 2013]; для Курской обл. приводился ошибочно [Дёмина, Никитина, 2008]. Наибольшее число местонахождений вида отмечено в республиках Башкортостан [Мулдашев, Маслова, Галеева, 2011] и Татарстан [Шайхутдинова, 2016], Волгоградской [Супрун, 2017], Ростовской [Шишлова, Шмараева, 2014] и Самарской [Ильина, Митрошенкова, Саксонов, 2017] обл. Популяции H. grandiflorum в перечисленных регионах многочисленны, стабильны; в Красных книгах Самарской и Волгоградской обл. вид приводится со статусом 5 – восстанавливающийся в численности.
Копеечник крупноцветковый как вид, обладающий федеральным статусом охраны, детально и всесторонне изучался на большем протяжении его российского ареала. Исследовались ценотическая приуроченность вида, его отношение к ведущим факторам среды, пространственная и возрастная структура популяций [Ильина, 2013а, 2015, 2019; Лаврентьев, Болдырев, 2017], процессы расообразования и интрогрессии в местах контакта с другими видами секции Subacaulia с использованием сравнительноморфологических и молекулярных методов [Князев, 2014; Супрун, 2014]. Описан большой жизненный цикл вида, даны характеристики онтогенетических состояний [Ильина, 2007; Супрун, 2014]. Проведены эксперименты и разработаны рекомендации к реинтродукции и восстановлению популяций вида [Мул-дашев и др., 2012; Лаврентьев, 2019; Maslova, Muldashev, Elizaryeva, 2019], к размножению в культуре in vitro [Ахметова, Зарипова, 2013; Малаева, 2016]. Особенности семенной репродукции данного вида копеечника изучены слабее, преимущественно в связи с поиском оптимальных условий проращивания и методик предпосевной обработки семян [Ильина, 2013б; Супрун, 2014; Лаврентьев, 2016]. Работы по определению семенной продуктивности немногочисленны [Кузнецова, 2008; Лаврентьев, 2016].
В Красной книге Ростовской обл. H. grandiflorum придана категория статуса редкости 3 в, д: редкий вид, имеющий узкую экологическую приуроченность, связанный со специфическими условиями произрастания и имеющий ограниченный ареал, часть которого находится в Ростовской обл. Известные местонахождения вида сосредоточены в северной (правобережье среднего течения Дона) и северо-западной (восточный отрог Донецкого кряжа, р. Калитва с притоками, Северное Приазовье) частях региона, доходя на восток по правому берегу Дона до Константиновского р-на. К югу от нижнего течения Дона копеечник крупноцветковый отмечался в нескольких точках по балкам, впадающим в ложе Цимлянского водохранилища. Таким образом, по долине Нижнего Дона в целом проходит южная граница распространения вида. Популяции H. grandiflorum в западных районах области многочисленны (100–800 тыс. особей), их состояние не вызывает опасений. Популяции в Константиновском и Дубовском р-нах у южной границы ареала малочисленны и фрагментарны, их численность составляет от нескольких десятков до 1–5 тыс. особей, стабильность их существования зависит от уровня антропогенной нагрузки на экотоп [Шишлова, Шмараева, 2014]. Возрастная структура популяций не определялась. Детальные сведения о состоянии и воспроизводстве популяций копеечника крупноцветкового в Ростовской обл. в литературе отсутствуют.
Цель нашей работы – оценка состояния и воспроизводства популяций Hedysarum grandiflorum в бассейне нижнего течения р. Дона. Поставленная цель включает уточнение распространения и условий произрастания вида на Нижнем Доне, определение онтогенетической структуры популяций, выявление основных лимитирующих факторов, влияющих на воспроизводство вида, а также определение основных показателей семенной продуктивности на примере вновь обнаруженной ценопопуляции H. grandiflorum в Аксайском р-не Ростовской обл.
Материалы и методы
Исследования проводили в 2019–2021 гг. в десяти районах Ростовской обл., относящихся к бассейнам р. Дон, Северский Донец, Сал, а также охватывающих побережье Цимлянского водохранилища (рис. 1). При обнаружении ценопопуляций (ЦП) H. grandiflorum выполнялись геоботанические описания фитоценозов, определялась примерная численность популяции и занимаемая ею площадь. Измерялись некоторые морфометрические показатели генеративных растений (диаметр в проекции вегетативных частей; высота, определяемая по размерам наиболее длинного репродуктивного побега; число цветоносов) на 20–30 особях каждой ЦП; для каждого показателя вычислялось среднее значение и его ошибка. Плотность особей определяли в местах концентрации на учётных площадках 1 м2. Учёт особей разных возрастных состояний проводился также на учётных площадках, либо (при небольшой численности и низкой плотности растений в ЦП) методом сплошного учёта на 70–120 экземпляров. Статистическая обработка числовых данных осуществлялась по стандартным методикам [Лакин, 1990] с использованием MS Excel 1997-2003. Вычисление индексов восстановления (I в ), замещения (I з ), старения (I ст ), индекса воз-растности дельта ( Δ ) и индекса эффективности омега ( ω ), определение положения популяций по классификации «дельта-омега» и по величине индекса замещения проводились по общепринятым методикам [Уранов, 1975; Жукова, 1987; Животовский, 2001; Жукова, Полянская, 2013].
Определение показателей семенной продуктивности H. grandiflorum осуществлялось по методике И.В. Вайнагий [1974] для ЦП1 в сезоны 2019 и 2020 гг., для ЦП2 – в 2019 г. Объём выборки в каждой ЦП составлял 130–150 плодов. Потенциальная семенная продуктивность (ПСП), или общее число семязачатков в завязи, принималась равной числу члеников плода. Помимо реальной семенной продуктивности (РСП), определяемой как число полноценных семян в бобе, подсчитывалась условно-реальная семенная продуктивность (УРСП) как сумма полноценных и начавших развитие, но не вызревших (поражённых или щуплых) семян. Коэффициент семенификации вычислялся как для реальной семенной продуктивности (K С(РСП) ), так и для условно-реальной (K С(УРСП) ). В связи с тем, что распределение счётных показателей семенной продуктивности отклоняется от нормального, статистическое сравнение частот вариантов для каждого показателя было выполнено с помощью непараметрического критерия хи-квадрат (χ2). В 2020 г. определялись также показатели семенной продуктивности в расчёте на соцветие и на особь.
Результаты исследования
Экспедиционные исследования 2019–2021 гг. показали, что распространение Hedysarum grandiflorum в регионе шире, чем приводится в Красной книге Ростовской обл. [Шишлова, Шмараева, 2014]. Все рассматриваемые далее местонахождения вида обнаружены впервые. Произрастание копеечника крупноцветкового впервые отмечено в низовьях р. Калитва (Белокалитвинский р-н, первая находка в районе), западной пограничной части Тарасовского р-на по р. Митякинка. В Аксайском р-не выявлены наиболее южные местонахождения по правобережью долины Нижнего Дона (две ценопопуляции, первые находки в районе). Особый интерес представляет обнаружение H. grandiflorum в системе балок по правому берегу р. Сал – левого притока нижнего течения Дона (Волгодонский р-н, первая находка в районе). До настоящего времени местонахождения данного вида к югу от нижнего течения Дона были известны лишь по берегу Цимлянского водохранилища.
В настоящей работе представлены результаты изучения 8 ЦП Hedysarum grandiflorum из 6 административных районов Ростовской обл.: Аксайского (2 ЦП), Белокалитвинского, Волгодонского, Дубовско-го (2 ЦП), Красносулинского и Тарасовского (рис. 1). Для Аксайского, Белокалитвинского и Волгодонского р-нов данный вид приводится впервые. По географической локализации популяции H. grandiflorum были разделены на правобережные (популяции 1–4 и 8), принадлежащие к бассейну р. Дона в нижнем его течении (Аксайский р-н) или к бассейну Северского Донца, крупнейшего правого притока Нижнего Дона (Красносулинский, Белокалитвинский, Тарасовский р-ны), и левобережные (популяции 5–7), относящиеся к бассейну р. Сал или балок, впадающих непосредственно в ложе Цимлянского водохранилища по его левому борту.
Описание условий произрастания ЦП представлено ниже.
ЦП1. Аксайский р-н, 0.5 км к северу от пос. Пчеловодный, правый коренной берег р. Аксай, верхняя часть остепнённого приводораздельного склона. Почвы: маломощный (смытый) щебневатый северопри- азовский высококарбонатный чернозём на известняке-ракушечнике. Сообщество: разнотравно-дерновиннозлаковая каменистая степь; ассоциация Stipa lessingiana + петрофитно-степное разнотравье.
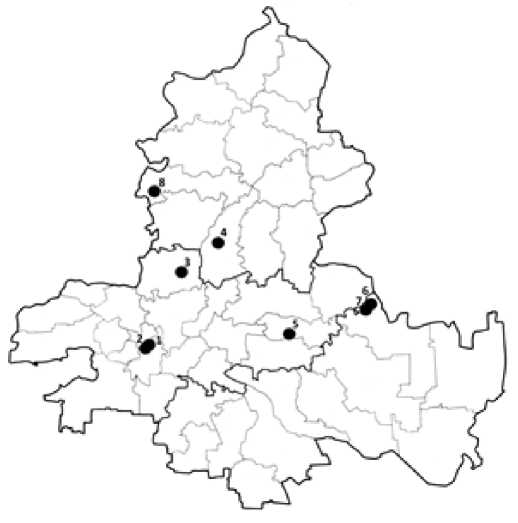
Рис. 1. Географическая локализация обследованных ценопопуляций Hedysarum grandiflorum [Geographical location of studied populations of Hedysarum grandiflorum ]
ЦП2. Аксайский р-н, близ южной окраины пос. Реконструктор, степной склон балки, открывающейся в долину р. Аксай. Почвы: маломощный североприазовский чернозём на известняке-ракушечнике. Сообщество: разнотравно-крупноковыльная каменистая степь; ассоциация Stipa zalesskii + петрофильно-степное разнотравье.
ЦП3. Красносулинский р-н, 4.5 км на запад от хутора (х.) Большая Федоровка, вершина поперечного отвершка балки Ясеновской, склон юго-западной экспозиции. Почвы: маломощный чернозём на плотном песчанике. Сообщество: разнотравно-дерновиннозла-ковая степь; ассоциация: Stipa lessingiana – Bromopsis riparia – Festuca valesiaca + разнотравье.
ЦП4. Белокалитвинский р-н, восточная окраина х. Крутинского, средняя часть склона левого коренного берега р. Калитвы. Почвы: чернозём примитивный на выветрелом песчанике. Сообщество: каменистая степь; ассоциация: Festuca valesiaca – Stipa lessingiana – Thymus dimorphus .
ЦП5. Волгодонский р-н, в 6.5 км западнее от х. Семёнкин, правый берег р. Сал, водоразделы поперечных балочек балки Чапелы. Почвы: чернозём южный маломощный. Со- общество: каменистая степь; ассоциация Galatella villosa + Festuca rupicola + разнотравье.
ЦП6. Дубовский р-н, 4 км к югу от ст-цы Малая Лучка, вершина склона балки, впадающей в Цимлянское водохранилище, в ее истоке. Почвы: каштановые на лёссовидном суглинке. Сообщество: дерновин-но-злаковая степь; ассоциация Stipa lessingiana + Festuca rupicola – Centaurea taliewii.
ЦП7. Дубовский р-н, 2.9 км юго-западнее х. Алдабульского, вершина склона балки Баклановской, мергелевая гряда. Почвы: чернозём примитивный на щебнистом мергеле. Сообщество: каменистая степь; ассоциация Stipa lessingiana + Cephalaria uralensis .
ЦП8. Тарасовский р-н, в 1.5 км северо-западнее х. Зелёновка, правый берег р. Митякинка, верхняя часть мелового склона западной экспозиции. Почвы: смытый чернозем на плотном мергеле. Сообщество: каменистая степь; ассоциация: Festuca rupicola + Stipa capillata + разнотравье.
Сведения о структурных характеристиках травостоя и количественных показателях ЦП приведены в табл. 1.
Все изученные ценопопуляции относительно невелики по численности (от 48 до 400–500 особей без учёта проростков) и занимаемой площади (как правило, несколько сотен квадратных метров, лишь для популяций 1, 3 и 6 свыше 1000 м2).
В большинстве местонахождений H. grandiflorum отличается сравнительно более высоким обилием относительно других видов разнотравья, не поднимаясь при этом до статуса субдоминанта, как это отмечается Н.А. Супрун [2013] в Волгоградской, а В.Н. Ильиной [2014, 2019] – в Самарской обл.
Пространственное распределение особей копеечника характеризуется отчётливо выраженной конта-гиозностью. Участки сгущений имеют от 2 до 5 м в диаметре; между ними растения копеечника встречаются со значительно меньшей плотностью. Плотность расположения агрегаций растений, число и распределение особей в промежутках между агрегациями находятся в прямой зависимости от общей численности популяции. Результаты наблюдений согласуются с данными, полученными В.Н. Ильиной [2013а, 2019] для средневолжских популяций вида. Общая плотность растений в местах концентрации на площади геоботанического описания варьирует от 2 ± 0 до 7 ± 2 экз./м2, плотность генеративных особей – от 2 ± 0 до 5 ± 0 экз./м2. Средняя плотность растений, с учётом характера их распределения, оказывается существенно ниже. Наблюдаемые значения плотности далеки от оптимальных величин, что объясняется общей малочисленностью популяций [Ильина, 2019].
Возрастная структура ценопопуляций определялась в 3 ЦП (ЦП1 в 2019 и 2020 гг., ЦП6 и 7 в 2021 г.) с выделением всех категорий онтогенетических состояний, в ЦП3 (2020 г.) и 5 (2021 г.) – с выделением трёх онтогенетических периодов: прегенеративного, генеративного и постгенеративного. Для копеечника крупноцветкового в Ростовской области во второй половине лета характерно явление полупокоя, выражающееся в усыхании и сбрасывании листочков на трети – половине листьев текущего года, что существенно усложняет учёт растений ранних прегенеративных состояний. В этой связи в популяциях 2, 4 и 8, которые обследовались в середине – второй половине июля, выявления онтогенетических спектров не проводилось.
Таблица 1 Основные количественные характеристики ценопопуляций Hedysarum grandiflorum и содержащих их фитоценозов
[General quantitative characters of Hedysarum grandiflorum coenopopulations and plant communities containing them]
|
Д £ |
§ и о ^ о ко о =^ |
1=1 ой В о ч к В ко О |
н о о у В ко О |
н о о CQ О и § к Д |
о д о |
& £ СО к К 9 ь< |
ко ^ н и |
и ,к ^ К Л К Н о О О 1^ со к 5 № К О и к |
О & < |
|
1 |
2019 |
5000 |
400 – 500 |
50 |
80 |
25 |
3 ± 1 |
2 ± 0 |
Умеренная (рекреация) |
|
2 |
2019 |
200 |
75 – 80 |
31 |
75 |
10 |
– |
– |
Незначительная (рекреация) |
|
3 |
2020 |
2000 |
300 – 400 |
60 |
75 |
40 |
7 ± 2 |
3 ± 1 |
Слабая (следы выпаса) |
|
4 |
2020 |
300 |
48 |
51 |
60 |
20 |
– |
– |
Высокая (выпас) |
|
5 |
2021 |
350 |
50 |
35 |
40 |
25 |
5 ± 1 |
5 ± 0 |
Высокая (выпас) |
|
6 |
2021 |
1300 |
150 – 200 |
28 |
50 |
30 |
– |
– |
Умеренная (выпас) |
|
7 |
2021 |
500 |
100 – 120 |
22 |
30 |
30 |
– |
– |
Слабая (выпас) |
|
8 |
2021 |
500 |
150 |
51 |
80 |
30 |
– |
2 ± 0 |
Высокая (выпас) |
* – величина не оценивалась.
Для всех обследованных популяций оказались характерны такие особенности онтогенетического спектра, как низкая доля особей постгенеративного периода (от 0 до 3.3%, при этом сенильные особи были обнаружены лишь в 2019 г. в ЦП1) и весомое преобладание генеративных растений (70.0–94.7%). Исключение составила лишь ЦП Красносулинского р-на, где в местах концентрации растений доля особей прегенеративного возраста составила 59.5%, в том числе суммарная доля имматурных и виргиниль-ных растений равнялась общей доле генеративных экземпляров. Наблюдаемое успешное семенное возобновление может быть отчасти обусловлено относительно высокой плотностью ценопопуляции (7 ± 2 растения на 1 м 2 в местах учёта). Как показано В.Н. Ильиной и др. [2021], взрослые растения, формируя условия среды, играют защитную роль для проростков. Существенный вклад могли также внести благоприятные погодные условия весны – начала лета 2020 г. (ранняя тёплая и солнечная весна и большое количество осадков в мае – начале июня).
Онтогенетическая структура ЦП 1, 6 и 7 отражена в табл. 2.
Подробно охарактеризованные популяции можно оценивать как нормальные, полночленные (ЦП1, 2019 г.) или чаще неполночленные (отсутствуют проростки и сенильные особи, иногда также ювенильные растения). Модальный класс распределения в ЦП1 за оба года наблюдений и в ЦП6 – зрелые генеративные растения. Онтогенетический спектр ЦП1 (Аксайский р-н) имеет выраженный правосторонний характер с долей зрелых генеративных особей, равной или превышающей долю молодых генеративных растений, тогда как в обоих ценопопуляциях Дубовского р-на (ЦП6 и 7) доля растений стадии g 3 оказалась крайне низкой (0–1.3%), при этом в ЦП7 доля молодых генеративных особей незначительно превышает процентное содержание растений среднего генеративного возраста.
Соотношение особей различных возрастных состояний в значительной степени определяется сезоном года. В особенности отчётливо это проявляется на примере ЦП1 Аксайского р-на: в 2019 г., когда выявление онтогенетической структуры проводилось в середине мая, суммарная доля проростков и ювенильных растений текущего года составила 5.0%, тогда как в 2020 г. в результате учёта, проведённого в середине июля, особей данных возрастных состояний выявлено не было. Наблюдаемое изменение обусловлено главным образом массовой гибелью проростков к середине вегетационного периода, что выступает характерной особенностью сезонной динамики возрастной структуры популяций изучаемого вида [Ильина, 2012]. Неполночленность онтогенетических спектров ЦП H. grandiflorum Дубовского района в части отсутствия проростков и ювенильных особей уже во второй половине мая (в ЦП6 также имматурных растений), вероятнее всего, обусловлена их элиминацией в результате пасквальной нагрузки.
Таблица 2
Характеристика возрастного состава популяций H. grandiflorum
[Characterization of H. grandiflorum populations’ ontogenetic structure]
|
№ популяции, год набл. |
Особи |
Возрастные группы |
||||||||
|
p |
j |
im |
v |
g1 |
g2 |
g3 |
ss |
s |
||
|
ЦП1, 2019 |
кол-во |
1 |
5 |
4 |
11 |
22 |
51 |
22 |
2 |
2 |
|
% |
0.83 ± 0.83 |
4.17 ± 1.82 |
3.33 ± 1.64 |
9.17 ± 2.63 |
18.33 ± 3.53 |
42.50 ± 4.51 |
18.33 ± 3.53 |
1.67 ± 1.17 |
1.67 ± 1.17 |
|
|
ЦП1, 2020 |
кол-во |
0 |
0 |
2 |
2 |
11 |
50 |
16 |
2 |
0 |
|
% |
0 ± 0 |
0± 0 |
2.41 ± 1.68 |
2.41 ± 1.68 |
13.25 ± 3.72 |
60.24± 5.67 |
19.28 ± 4.33 |
2.41 ± 1.68 |
0 ± 0 |
|
|
ЦП6, 2021 |
кол-во |
0 |
0 |
0 |
4 |
14 |
56 |
1 |
0 |
0 |
|
% |
0 ± 0 |
0 ± 0 |
0 ± 0 |
5.33± 2.59 |
18.67 ± 4.50 |
74.67 ± 5.02 |
1.33 ± 1.32 |
0 ± 0 |
0 ± 0 |
|
|
ЦП7, 2021 |
кол-во |
0 |
0 |
4 |
19 |
29 |
27 |
0 |
1 |
0 |
|
% |
0 ± 0 |
0 ± 0 |
5.00 ± 2.44 |
23.75 ± 4.76 |
36.25 ± 5.37 |
33,75 ± 5.29 |
0 ± 0 |
1.25 ± 1.24 |
0 ± 0 |
|
Демографические показатели, отражающие состояние изученных популяций, представлены в табл. 3.
Таблица 3
Некоторые демографические показатели нижнедонских популяций H. grandiflorum [ H. grandiflorum Lower Don populations’ demographic characteristics]
|
6 в 2 в сЗ В в ° ® к У О |
X 3 • • 1 ? ? Он . Д О + .. м Г В 60 о в + § & и |
X О 60 ” ад В °х и |
В В 2 и в К |
в 9 § со в |
в б о в |
Он и в К |
Я в К |
2 & 2 в S у 5 в о в в |
В в cd В 2 8 о о « i У о g £ н в я |
|
ЦП1, 2019 |
17.43:79.16:3.34 |
23.16:53.68:23.16 |
0.22 |
0.21 |
0.04 |
0.440 |
0.771 |
зрелая |
Неустойчивая |
|
ЦП1, 2020 |
4.82:92.77:2.41 |
14.29:64.94:20.78 |
0.05 |
0.05 |
0.03 |
0.503 |
0.883 |
зрелая |
Неустойчивая |
|
ЦП3, 2020 |
59.52:40.48:0 |
1.47 |
1.47 |
0.00 |
Перспективная |
||||
|
ЦП5, 2021 |
11.54:88.46:0 |
0.13 |
0.13 |
0.00 |
Неустойчивая |
||||
|
ЦП6, 2021 |
5.33:94.67:0 |
19.72:78.87:1.41 |
0.06 |
0.06 |
0.00 |
0,440 |
0.926 |
зрелая |
Неустойчивая |
|
ЦП7, 2021 |
28.75:70.00:1.25 |
51.79:48.21:0 |
0.41 |
0.40 |
0.02 |
0.308 |
0.737 |
зреющая |
Неустойчивая |
По классификации, предложенной Л.А. Жуковой и Т.А. Полянской [2013] на основе индекса замещения, большинство обследованных ЦП с долей растений прегенеративного периода от 4,8 до 28.8% и индексами замещения, колеблющимися от 0.05 до 0.40, было отнесено к категории неустойчивых. Исключение составляет ЦП3 (Красносулинский р-н), онтогенетическая структура которой позволяет рассматривать её как перспективную (Iз = 1,47). По классификации «дельта – омега» ЦП1 и 6 были отнесены к группе зрелых, тогда как ЦП7 – к категории зреющих.
Сравнение полученных данных с результатами исследований В.Н. Ильиной [2013а, 2019],
А.А. Мулдашева и др. [2013], Н.А. Супрун [2013], Л.М. Абрамовой и др. [2015, 2019], Б.М. Фардеевой и
А.М. Зариповой [2018] в других регионах России показывает следующее. Онтогенетический спектр нижнедонских популяций H. grandiflorum по соотношению групп растений разных периодов онтогенеза и преобладанию средневозрастных генеративных особей в целом соответствует базовому онтогенетическому спектру популяций вида Средней Волги [Ильина, 2019] и Предуралья [Абрамова и др., 2015, 2019]. Вместе с тем, правосторонний характер распределения, связанный с более слабой представленностью в большинстве изученных ЦП молодых генеративных и в особенности прегенеративных растений, обусловил отнесение их к зрелым неустойчивым. Это в большей степени сближает демографическую структуру нижнедонских ЦП копеечника со структурой среднедонских популяций, изученных Н.А. Супрун [2013], среди которых также преобладают зрелые, а также средневолжских, представленных преимущественно зрелыми со значительной долей переходных [Ильина, 2019]. В Татарском Закамье [Фардеева, Зарипова, 2018] и Предуралье [Мулдашев и др., 2013; Абрамова и др., 2015, 2019] большинство ценопопуляций вида отнесено к молодым перспективным, в волгоградской части Приволжской возвышенности [Супрун, 2013] – к переходным. Таким образом, онтогенетическая структура популяций H. grandiflorum у южных пределов распространения в европейской части России свидетельствует о большей уязвимости их естественного воспроизводства по сравнению с популяциями в центральной части ареала и у северных и восточных его границ.
Сравнительный анализ количественных характеристик и демографической структуры ЦП копеечника на Нижнем Доне в различных условиях произрастания свидетельствует о том, что ведущим фактором, в современных условиях Ростовской обл., определяющим численность, устойчивость и способность к са-мовоспроизводству популяций H. grandiflorum , выступает антропопрессия, выражающаяся прежде всего в форме пасквальной нагрузки. Этот фактор наиболее значим для левобережных популяций, обусловливая низкую плотность ЦП6 и 7 копеечника в Дубовском р-не, малую общую численность ЦП5 в Волгодонском, а также ЦП4 в Белокалитвинском р-нах. Влияние рекреационной нагрузки (ЦП1) несколько менее существенно.
Решающая роль антропогенного фактора подтверждается данными о возрастной структуре ЦП1, 3, 5– 7. Как показано В.Н. Ильиной [2015] по итогам многолетних исследований H. grandiflorum в Самарской обл., в условиях значительной антропогенной нагрузки общая доля генеративных особей в популяциях данного вида возрастает с 64 до более 80%. Среди анализируемых популяций из Ростовской обл. такое распределение свойственно ЦП1, 5 и 6; популяция 7 занимает промежуточное положение, тогда как в ЦП3, произрастающей в условиях незначительной антропопрессии, прегенеративные особи преобладают над генеративными. Одновременно с этим, увеличения доли старовозрастных генеративных и сенильных растений в популяциях 1, 6 и 7 не наблюдалось, абсолютный максимум здесь приходится на особи возрастной стадии g 2 , реже g 1 . Это позволяет предположить, что уровень антропогенной нагрузки на описываемые сообщества не является критическим для поддержания популяций копеечника.
Среди других значимых факторов, определяющих состояние популяций копеечника крупноцветкового в Ростовской обл., необходимо назвать климатические и эдафические условия местообитания. По данным М.В. Лаврентьева и В.А. Болдырева [2017], H. grandiflorum – достаточно стенобионтный вид, оптимум развития которого наблюдается на маломощных небогатых почвах, развивающихся на карбонатных породах, при среднем степном увлажнении. Лимитирующим фактором для нижнедонских популяций вида у юго-восточного предела распространения (ЦП5, 6), вероятнее всего, является недостаточное количество осадков. Относительная малочисленность и низкая общая средняя плотность левобережных популяций H. grandiflorum согласуется с данными, приводимыми в Красных книгах Ростовской обл. [Шишлова, Шмараева, 2014] и Калмыкии [Бакташева, 2014]. Невысокая доля прегенеративных особей в ЦП5–6, наряду с влиянием сбоя, может объясняться сухостью субстрата, ветровой и водной эрозией, как было выявлено Л.М. Абрамовой и др. [2019] в Предуралье. Вместе с тем, возрастной спектр ЦП7 отражает более благоприятную ситуацию в отношении семенного возобновления как по причине менее выраженной пасквальной нагрузки, так и по характеру субстрата – чернозём примитивный на щебнистом мергеле. В ЦП8 сходные эдафические условия частично компенсируют высокую интенсивность выпаса. Популяции копеечника, описанные в западной (ЦП1–3) и северной (ЦП8) частях области, находятся в более благоприятных условиях увлажнения, что положительно сказывается на их состоянии и воспроизводстве.
Структурные характеристики травостоя как фактор, определяющий численность и воспроизводство копеечника, в обследованных популяциях играет менее значимую роль по сравнению с рассмотренными выше. Невысокое проективное покрытие травостоя, наблюдаемое в местообитаниях ЦП 4–7, в большей степени является следствием пастбищной дигрессии степных сообществ. Строгой зависимости между величиной общего проективного покрытия и количественными показателями популяций копеечника не выявлено.
Морфометрические характеристики генеративных растений H. grandiflorum в обследованных популяциях приведены в табл. 4.
Таблица 4
Основные морфометрические показатели растений H. grandiflorum нижнедонских популяций [General morphometric characteristics of H. grandiflorum plants from Lower Don populations]
|
№ популяции, год обследования |
Высота растения, см |
Диаметр растения, см |
Число цветоносов |
|
(x min ) x ср ± s x (x max ) |
(x min ) x ср ± s x (x max ) |
(x min ) x ср ± s x (x max ) |
|
|
ЦП1, 2019 |
(16) 31.6 ± 1.1 (43) |
– |
(1) 9 ± 1 (42) |
|
19.23* |
– |
94.43 |
|
|
ЦП1, 2020 |
(22) 36.3 ± 1.0 (52) |
– |
(1) 9 ± 1 (27) |
|
19.00 |
– |
61.25 |
|
|
ЦП3, 2020 |
(19) 35.2 ± 2.3 (47) |
(36) 51.0 ± 4.3 (61) |
(1) 9 ± 3 (28) |
|
20,43 |
18.81 |
92.08 |
|
|
ЦП5, 2021 |
(13) 18.4 ± 1.9 (24) |
(40) 47.0 ± 3.0 (57) |
(1) 5 ± 1 (11) |
|
22.61 |
14.43 |
65.10 |
|
|
ЦП6, 2021 |
(26) 30.9 ± 1.1 (38) |
(38) 47.8 ± 2.2 (60) |
(9) 23 ± 5 (62) |
|
12.04 |
14.28 |
69.41 |
|
|
ЦП7, 2021 |
(16) 20.4 ± 1.4 (25) |
(23) 39.5 ± 3.1 (58) |
(2) 8 ± 1 (15) |
|
21.70 |
25.04 |
53.19 |
|
|
ЦП8, 2021 |
(40) 50.6 ± 1.2 (58) |
(9) 26.5 ± 3.4 (42) |
(10) 16 ± 2 (31) |
|
9.35 |
43.93 |
46.94 |
* Для каждого показателя под чертой указано значение коэффициента вариации Cv, %.
Сравнительный анализ полученных данных с литературными демонстрирует, что наиболее значительные отличия наблюдаются в значении среднего числа цветоносов на растение. По данным В.Н. Ильиной [2005], этот показатель в средневолжских популяциях колеблется от 1.0 ± 0.2 до 4.2 ± 0.35 в зависимости от возрастного состояния и уровня жизненности особей, тогда как в ЦП Ростовской обл. он существенно выше – от 5 ± 1 до 23 ± 5. Это свидетельствует о более благоприятных условиях для формирования соцветий и, вероятнее всего, обусловлено более высокими температурами периода вегетации.
Семенная продуктивность (СП) копеечника крупноцветкового изучалась в двух ЦП Аксайского р-на: в ЦП1 в 2019‒2020 гг. и в ЦП2 ‒ в 2019 г. Показатели СП в расчёте на плод приведены в табл. 5 и на рис. 2.
Таблица 5
Показатели семенной продуктивности Hedysarum grandiflorum в Аксайском р-не в расчёте на плод [Seed productivity characteristics per fruit of Hedysarum grandiflorum from Aksay district]
|
№ популяции, |
ПСП |
РСП |
УРСП |
К С(РСП) , % |
К С(УРСП) , % |
|
год наблюдений |
(x min )M±m M (x max ) |
(x min )M±m M (x max ) |
(x min )M±m M (x max ) |
(x min )M±m M (x max ) |
(x min )M±m M (x max ) |
|
ЦП1, 2019 |
(1) 4 ± 0 (6) |
(0) 0 ± 0 (2) |
(1) 2 ± 0 (5) |
9.29 ± 1.40 |
58.87 ± 1.68 |
|
ЦП2, 2019 |
(1) 3 ± 0 (6) |
(0) 0 ± 0 (3) |
(1) 2 ± 0 (5) |
13.11 ± 1.69 |
74.72 ± 1.76 |
|
ЦП1, 2020 |
(1) 3 ± 0 (5) |
(0) 1 ± 0 (5) |
(1) 2 ± 0 (5) |
34.54 ± 2.90 |
77.28 ± 1.85 |
Потенциальная семенная продуктивность варьирует от 1 до 6 семязачатков на плод в 2019 г. и от 1 до 5 в ЦП1 в 2020 г. Модальный класс распределения совпадает со средним значением показателя и составил 4 семязачатка на плод в ЦП1 по данным 2019 г. и 3 семязачатка на плод в двух остальных случаях. Выявленные отличия в характере распределения показателей ПСП в ЦП1 в 2019 г. от двух других изученных случаев статистически подтверждены (χ2 = 71.8 по годам наблюдений и χ2 = 31.3 для двух ЦП в 2019 г.). Наблюдаемая картина в целом соответствует данным других авторов [Кузнецова, 2008; Супрун и др., 2020]. Реальная семенная продуктивность в ЦП копеечника Аксайского р-на в годы наблюдений оказалась очень низкой. Во всех учтённых выборках преобладают плоды, не содержащие полноценных семян. В 2019 г. их доля составила 70.5% в ЦП1 и 66.0% – в ЦП2. Среднее число развитых семян на плод – 0; величины коэффициента семенификации составили 9.29% и 13.11% соответственно. В 2020 г. в ЦП1 большинство плодов также не имело развитых семян, однако их доля снизилась до 40.9%, что сопровождалось существенным увеличением числа плодов с 2 и 3 развитыми семенами. Наблюдаемые различия в структуре реальной семенной продуктивности ценопопуляции 1 по годам наблюдений носят достоверный характер (χ2 = 28.8), между популяциями в 2019 г. недостоверны. Среднее значение РСП в ЦП1 в 2020 г. составило 1 полноценное семя на плод, коэффициент семенификации K С(РСП) возрос до 34.54%, что можно рассматривать как достаточно высокое значение.
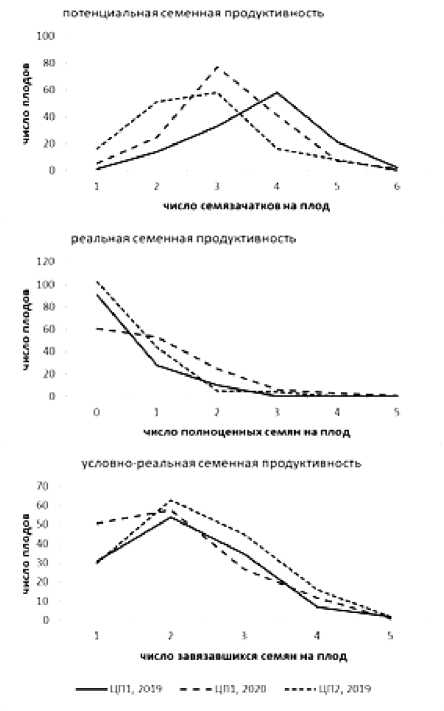
Рис. 2. Структура семенной продуктивности H. grandiflorum в расчёте на плод
[ H. grandiflorum seed productivity structure per one fruit]
Структура условно-реальной семенной продуктивности в 2019 г. в двух ЦП выглядит очень схожей, с модальным классом распределения 2 завязавшихся семян на плод, что совпадает со средним значением показателя. В 2020 г. в ЦП1 значения УРСП несколько ниже за счёт значительного увеличения доли плодов с одним завязавшимся семенем и менее существенного уменьшения числа плодов с 3 и более семенами. Тем не менее, в отличие от двух предыдущих показателей, значение УРСП на плод не отличается достоверно как в 2019 г. между ценопопуляциями, так и в ЦП1 по годам наблюдений. Коэффициент семенификации для УРСП (K С(УРСП) ) весьма высок и варьирует от 58.87 до 77.28%.
Предполагая, что значения РСП в большей степени зависят от отклонений в развитии семязачатков на презиготических и ранних постзиготических стадиях, тогда как величины УРСП отражают успешность последующих стадий развития формирующихся семян, можно заключить следующее. Величина условно-реальной семенной продуктивности наиболее стабильна и наименее зависит от условий произрастания и текущего сезона. Это свидетельствует об успешности процессов опыления и оплодотворения цветков, как благодаря стабильно низкой степени стерильности семязачатков, так и вследствие предполагаемой высокой активности и регулярной работы опылителей. Крайне невысокие значения реальной семенной продуктивности и её достоверные различия по годам наблюдений однозначно свидетельствуют об экзогенной природе нарушений в развитии завязавшихся семян, причём наибольший вклад, вероятно, вносят погодные условия текущего сезона.
Показатели семенной продуктивности в расчёте на растение определялись в 2020 г. в ЦП1. Среднее число развитых соцветий составило 9 ± 1; среднее число плодов в соцветии – 37 ± 1. Среднее число семязачатков в плоде оказалось равным 3 ± 0, а у особи – 914. При этом среднее число полноценных семян в плоде составило 1 ± 0, а у особи – 318.
Сходные значения были получены М.В. Лаврентьевым [2016] в популяциях копеечника южной части Приволжской возвышенности (Саратовская обл.): при пересчёте потенциальная семенная продуктивность оказывается в среднем равна 442 семенам на особь, а реальная – 286 на особь. Однако, учитывая низкие значения KС(РСП) в обеих изученных популяциях в 2019 г., есть все основания предполагать, что в годы, неблагоприятные для вызревания семян, семенная продуктивность H. grandiflorum существенно снижается. В частности, коэффициент семенификации для РСП в ЦП1 в 2019 г. был почти в 4 раза ниже по сравнению с 2020 г., тогда как среднее число развитых соцветий у растений ЦП1 за два сезона наблюдений достоверно не различалось. Таким образом, можно заключить, что фактическая семенная продуктивность H. grandiflorum в расчёте на растение существенно варьирует по годам, что согласуется с литературными данными о нерегулярном семенном возобновлении данного вида [Супрун, 2013].
Заключение
Ценопопуляции копеечника крупноцветкового в бассейне нижнего течения р. Дона приурочены к типичным местообитаниям вида – каменистым степям на маломощных почвах, сформированных на подстилающих породах различного литологического состава, преимущественно карбонатных. По общей численности, плотности, ценотическим характеристикам популяции H. grandiflorum на изучаемой территории уступают популяциям центральной, северной и восточной частей ареала. Основные факторы, ограничивающие распространение и затрудняющие воспроизводство вида на Нижнем Дону – это антропогенное воздействие и, в меньшей степени, климатические и эдафические условия произрастания. Антропогенная нагрузка, преимущественно в форме пастбищной, обусловливает низкую плотность и относительную малочисленность ценопопуляций вида, затрудняя его семенное воспроизводство за счёт эли- минации растений ранних прегенеративных стадий. В условиях незначительной антропопрессии структурные характеристики и онтогенетический состав ценопопуляций приближаются к оптимальным.
Семенная продуктивность H. grandiflorum в расчёте на плод в изученных ценопопуляциях в целом очень низка и подвержена погодичным колебаниям, при этом среднее число генеративных единиц на растение больше, чем наблюдается в средневолжских популяциях вида. Фактическую семенную продуктивность в расчёте на особь можно расценивать как удовлетворительную.
Состояние большинства обследованных популяций центральных и западных районов Ростовской обл. стабильно и не вызывает опасений, тогда как численность и воспроизводство ценопопуляций у юговосточной границы распространения вида лимитируется уровнем антропогенной нагрузки на экотоп. Результаты исследования подтверждают соответствие категории редкости Hedysarum grandiflorum , принятой в действующем издании Красной книги Ростовской области, текущему состоянию популяций вида в пределах региона.
Список литературы Состояние и воспроизводство нижнедонских популяций копеечника крупноцветкового (Hedysarum grandiflorum Pall., Fabaceae)
- Абрамова Л.М., Каримова О.А., Мустафина А.Н. Структура ценопопуляций редкого вида Hedysarum grandiflorum Pall. в петрофитных степях Предуралья // Принципы и способы сохранения биоразнообразия: материалы VI Всерос. конф. Йошкар-Ола, 2015. С. 136-138.
- Абрамова Л.М. и др. Структура и состояние популяций трёх редких видов рода Hedysarum (Faba-ceae) на Южном Урале // Ботанический журнал. 2019. Т. 104, № 5. С. 729-740.
- Ахметова А.Ш., Зарипова А.А. Размножение видов рода Hedysarum L. in vitro. // Принципы и способы сохранения биоразнообразия: материалы V Междунар. науч. конф. Йошкар-Ола, 2013. Ч. I. С. 227230.
- Бакташева Н.М. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Республики Калмыкия. Элиста: Джангар, 2014. Т. 2. Редкие и находящиеся под угрозой исчезновения растения и грибы. С. 130.
- Вайнагий И.В. О методике изучения семенной продуктивности растений // Ботанический журнал. 1974. Т. 59, вып. 6. С. 826-831.
- Васильева Л.И. Род Копеечник - Hedysarum L. // Флора европейской части СССР. Л.: Наука, 1987. Т. 7. С. 87-93.
- Дёмина О.Н., Никитина С.В. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Российской Федерации (растения и грибы). М.: Т-во науч. изд. КМК, 2008. С. 240-241.
- Животовский Л.А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология. 2001. № 1. С. 3-7.
- Жукова Л.А. Динамика ценопопуляций луговых растений в естественных фитоценозах // Динамика популяций травянистых растений. Киев: Наукова думка, 1987. С. 9-19.
- Жукова Л.А., Полянская Т.А. О некоторых подходах к прогнозированию перспектив развития це-нопопуляций растений // Вестник Тверского государственного университета. Сер. Биология и экология. 2013. Вып. 32, № 31. С. 160-171.
- Ильина В.Н. Жизненность и виталитетная структура ценопопуляций Hedysarum grandiflorum Pall. и H. rasoumovianum Fisch. et Helm в Самарской области // Самарская Лука: Бюл. 2005. № 16. С. 179-186.
- Ильина В.Н. Онтогенез копеечника крупноцветкового (Hedysarum grandiflorum Pall.) // Онтогенетический атлас растений. Йошкар-Ола: МарГУ, 2007. Т. 5. С. 126-132.
- Ильина В.Н. Особенности погодичной и сезонной динамики онтогенетической структуры популяций копеечника крупноцветкового // Раритеты флоры Волжского бассейна: докл. участников II Рос. науч. конф. Тольятти: Кассандра, 2012. С. 109-110.
- Ильина В.Н. О биоэкологических особенностях копеечника крупноцветкового (Hedysarum grandi-florum Pall., Fabaceae) в Самарской области // Самарский научный вестник. 2013а. № 4(5). С. 78-80.
- Ильина В.Н. Перспективы интродукции некоторых видов семейства бобовые в связи с особенностями начальных периодов онтогенеза // Самарский научный вестник. 2013б. № 3(4). С. 44-47.
- Ильина В.Н. Структура и состояние популяций средневолжских видов рода Hedysarum L. (Fabaceae) // Самарский научный вестник. 2014. № 2(7). С. 37-40.
- Ильина В.Н. Изменения базовых онтогенетических спектров популяций некоторых редких видов растений Самарской области при антропогенной нагрузке на местообитания // Самарская Лука: проблемы региональной и глобальной экологии. 2015. Т. 24, № 3. С. 144-170.
- Ильина В.Н. Онтогенетическая структура и типы ценопопуляций копеечника крупноцветкового (Hedysarum grandiflorum Pall.) в бассейне Средней Волги // Весщ Нацыянальнай акадэмп навук Беларусь Серыя бiялагiчных навук. (Известия Национальной академии наук Беларуси. Сер. Биол. науки). 2019. Т. 64, № 3. C. 302-310.
- Ильина В.Н. и др. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Самарской области. Самара, 2017. Т. 1. Редкие виды растений и грибов. С. 121.
- Ильина В.Н., Атаджанов И.Р., Власенко Н.В. Об онтогенетических консорциях Hedysarum grandi-florum на особо охраняемых природных территориях Самарской области // Самарская Лука: проблемы региональной и глобальной экологии. 2021. Т. 30, № 2. С. 59-60.
- Князев М.С. Бобовые (Fabaceae Lindl.) Урала: видообразование, географическое распространение, эколого-исторические свиты: дис. ... д-ра биол. наук. Екатеринбург, 2014. Т. 1. 144 с.
- Кузнецова М.Н. Семенное воспроизведение Копеечника крупноцветкового (Hedysarum grandiflorum Pall.) // Современные проблемы морфологии и репродуктивной биологии семенных растений: материалы междунар. конф. Ульяновск, 2008. С. 75-84.
- Лаврентьев М.В. Характеристика репродуктивных особенностей Hedysarum grandiflorum (Fabaceae) в южной части Приволжской возвышенности // Бюллетень Бот. сада Саратов. гос. ун-та. 2016. Т. 14, вып. 2. С. 35-43.
- Лаврентьев М.В. Особенности охраны Hedysarum grandiflorum Pall. // Экобиотех. 2019, Т. 2, № 4. С. 515-519.
- Лаврентьев М.В., Болдырев В.А. Характеристика местообитаний и адаптации к ним Hedysarum grandiflorum Pall. (Fabaceae, Dicotyledones) в южной части Приволжской возвышенности // Поволжский экологический журнал. 2017. № 1. С. 54-61.
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 350 с.
- Малаева Е.В. Изучение особенностей микроклонального размножения некоторых видов редких растений // Актуальные вопросы теории и практики биологического образования: материалы 10 всерос. науч.-практ. конф. М.: Планета, 2016. С. 93-95.
- Мулдашев А.А., Маслова Н.В., Галеева А.Х. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Республики Башкортостан. Уфа: МедиаПринт, 2011. Т. 1. Растения и грибы. С. 153.
- Мулдашев А.А. и др. Создание искусственных популяций редких видов рода Hedysarum L. (Fabaceae) в Республике Башкортостан // Известия Самарского научного центра РАН. 2012. Т. 14, № 1(7). С. 1791-1795.
- Мулдашев А.А. и др. Характеристика возрастного состава популяций Hedysarum grandiflorum Pall. (Fabaceae) в Башкирском Предуралье // Вестник Оренбургского государственного университета. 2013. № 10(159). С. 198-201.
- Новикова Л.А. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Пензенской области. Пенза, 2013. Т. 1. Грибы, лишайники, мхи, сосудистые растения. С. 124.
- Супрун Н.А. Структура популяций Hedysarum grandiflorum Pall. в Волгоградской области // Известия Самарского научного центра РАН. 2013. Т. 15, № 3(1). С. 346-351.
- Супрун Н.А. Копеечники (Hedysarum L.) Нижнего Поволжья: изменчивость и систематика: дис. ... канд. биол. наук. М., 2014. 160 с.
- Супрун Н.А. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Волгоградской обл. Воронеж: Издат-Принт, 2017. Т. 2. Растения и другие организмы. С. 136.
- Супрун Н.А., Малаева Е.В., Шумихин С.А. Особенности семенного размножения Hedysarum grandiflorum Pall. ex situ и in vitro // Вестник Пермского университета. Сер. Биология. 2020. Вып. 4. С. 286-293.
- Уранов А.А. Возрастной спектр фитоценопопуляции как функция времени и энергетических волновых процессов // Биологические науки. 1975. № 2. С. 7-34.
- Фардеева М.Б., Зарипова А.М. Состояние популяций Hedysarum grandiflorum Pall. и Oxytropis hip-polyti Boriss. на границе ареала // Систематические и флористические исследования Северной Евразии: материалы II междунар. конф. М., 2018. С. 80-84.
- Шайхутдинова Г.А. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Республики Татарстан (животные, растения, грибы). Казань: Идел-Пресс, 2016. С. 388-389.
- Шишлова Ж.Н., Шмараева А.Н. Копеечник крупноцветковый Hedysarum grandiflorum Pall. // Красная книга Ростовской области. Ростов-на-Дону, 2014. Т. 2. Растения и грибы. С. 196.
- Maslova N.V., Muldashev A.A., Elizaryeva O.A. Creating Rare Species Artificial Populations of the Genus Hedysarum L. (Fabaceae) // IOP Conf. Series: Earth and Environmental Science. 2019. Р. 022200.
